
Aptamer BC 007’s Affinity to Specific and Less-Specific Anti-SARS-CoV-2 Neutralizing Antibodies

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Nuclear Magnetic Resonance (NMR) Spectroscopy
2.3. High-Sensitivity Isothermal Titration Calorimetry (ITC)
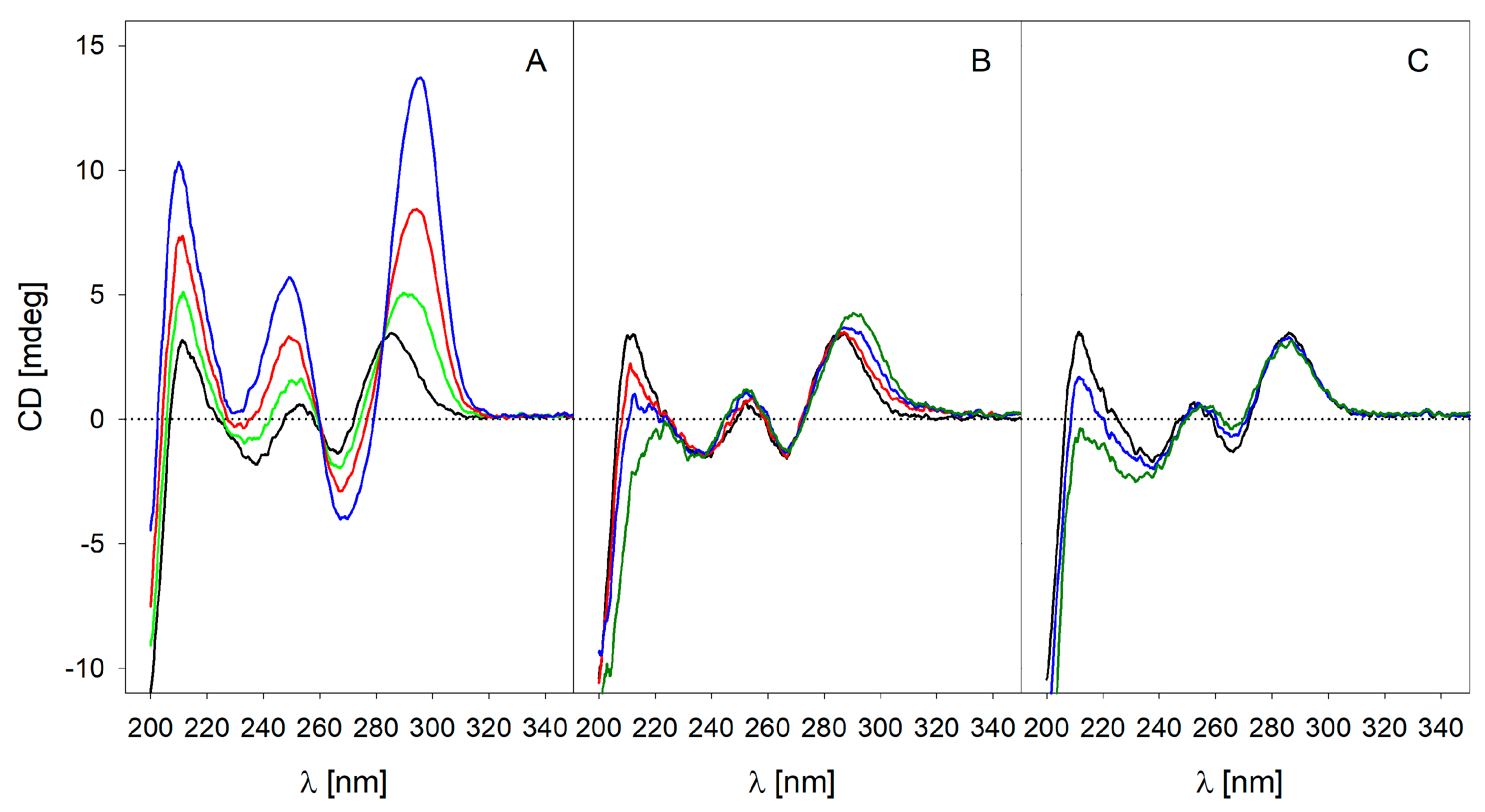
2.4. Circular Dichroism (CD) Spectroscopy
2.5. Enzyme-Linked Immunosorbent Assay (ELISA) Binding Experiments
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weisshoff, H.; Krylova, O.; Nikolenko, H.; Düngen, H.-D.; Dallmann, A.; Becker, S.; Göttel, P.; Müller, J.; Haberland, A. Aptamer BC 007—Efficient Binder of Spreading-Crucial SARS-CoV-2 Proteins. Heliyon 2020, 6. [Google Scholar] [CrossRef]
- Becker, N.-P.; Haberland, A.; Wenzel, K.; Göttel, P.; Wallukat, G.; Davideit, H.; Schulze-Rothe, S.; Hönicke, A.-S.; Schimke, I.; Bartel, S.; et al. A Three-Part, Randomised Study to Investigate the Safety, Tolerability, Pharmacokinetics and Mode of Action of BC 007, Neutraliser of Pathogenic Autoantibodies Against G-Protein Coupled Receptors in Healthy, Young and Elderly Subjects. Clin. Drug Investig. 2020, 49, 433–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberland, A.; Holtzhauer, M.; Schlichtiger, A.; Bartel, S.; Schimke, I.; Müller, J.; Dandel, M.; Luppa, P.B.; Wallukat, G. Aptamer BC 007—A Broad Spectrum Neutralizer of Pathogenic Autoantibodies against G-Protein-Coupled Receptors. Eur. J. Pharmacol. 2016, 789, 37–45. [Google Scholar] [CrossRef]
- Müller, J.; Haberland, A.; Wallukat, G.; Becker, N.-P.; Wenzel, K.; Göttel, P.; Schulze-Rothe, S.; Schimke, I.; Yilmaz, T.; Abay, A.; et al. The DNA-Based Drug BC 007 Neutralizes Agonistically Acting Autoantibodies Directed Against G Protein–Coupled Receptors Successful Mode of Action Demonstrated in Clinical Phase 1 Trial. Chem. Today 2019, 37, 65–67. [Google Scholar]
- Düngen, H.-D.; Dordevic, A.; Felix, S.B.; Pieske, B.; Voors, A.A.; McMurray, J.J.V.; Butler, J. Β1-Adrenoreceptor Autoantibodies in Heart Failure: Physiology and Therapeutic Implications. Circ. Heart Fail. 2020, 13, e006155. [Google Scholar] [CrossRef] [PubMed]
- Caso, F.; Costa, L.; Ruscitti, P.; Navarini, L.; Del Puente, A.; Giacomelli, R.; Scarpa, R. Could Sars-Coronavirus-2 Trigger Autoimmune and/or Autoinflammatory Mechanisms in Genetically Predisposed Subjects? Autoimmun. Rev. 2020, 19, 102524. [Google Scholar] [CrossRef]
- Halpert, G.; Shoenfeld, Y. SARS-CoV-2, the Autoimmune Virus. Autoimmun. Rev. 2020, 19, 102695. [Google Scholar] [CrossRef] [PubMed]
- Masuccio, F.G.; Barra, M.; Claudio, G.; Claudio, S. A Rare Case of Acute Motor Axonal Neuropathy and Myelitis Related to SARS-CoV-2 Infection. J. Neurol. 2020, 17, 1–4. [Google Scholar] [CrossRef]
- Rodríguez, Y.; Novelli, L.; Rojas, M.; De Santis, M.; Acosta-Ampudia, Y.; Monsalve, D.M.; Ramírez-Santana, C.; Costanzo, A.; Ridgway, W.M.; Ansari, A.A.; et al. Autoinflammatory and Autoimmune Conditions at the Crossroad of COVID-19. J. Autoimmun. 2020, 114, 102506. [Google Scholar] [CrossRef]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Liu, F.; Zheng, N.S.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse Functional Autoantibodies in Patients with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Novelli, L.; Motta, F.; De Santis, M.; Ansari, A.A.; Gershwin, M.E.; Selmi, C. The JANUS of Chronic Inflammatory and Autoimmune Diseases Onset during COVID-19—A Systematic Review of the Literature. J. Autoimmun. 2020, 117, 102592. [Google Scholar] [CrossRef]
- Khamsi, R. Rogue Antibodies Could Be Driving Severe COVID-19. Nature 2021, 590, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Lerma, L.A.; Chaudhary, A.; Bryan, A.; Morishima, C.; Wener, M.H.; Fink, S.L. Prevalence of Autoantibody Responses in Acute Coronavirus Disease 2019 (COVID-19). J. Transl. Autoimmun. 2020, 3, 100073. [Google Scholar] [CrossRef] [PubMed]
- Wallukat, G.; Hohberger, B.; Wenzel, K.; Fürst, J.; Schulze-Rothe, S.; Wallukat, A.; Hönicke, A.S.; Müller, J. Functional Autoantibodies against G-Protein Coupled Receptors in Patients with Persistent Long-COVID-19 Symptom. J. Transl. Autoimmun. 2021, 4, 100100. [Google Scholar] [CrossRef]
- Bock, L.C.; Griffin, L.C.; Latham, J.A.; Vermaas, E.H.; Toole, J.J. Selection of Single-Stranded DNA Molecules That Bind and Inhibit Human Thrombin. Nature 1992, 355, 564–566. [Google Scholar] [CrossRef]
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A Neutralizing Human Antibody Binds to the N-Terminal Domain of the Spike Protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Kreer, C.; Zehner, M.; Weber, T.; Ercanoglu, M.S.; Gieselmann, L.; Rohde, C.; Halwe, S.; Korenkov, M.; Schommers, P.; Vanshylla, K.; et al. Longitudinal Isolation of Potent Near-Germline SARS-CoV-2-Neutralizing Antibodies from COVID-19 Patients. Cell 2020, 182, 843–854.e12. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Su, B.; Guo, X.; Sun, W.; Deng, Y.; Bao, L.; Zhu, Q.; Zhang, X.; Zheng, Y.; Geng, C.; et al. Potent Neutralizing Antibodies against SARS-CoV-2 Identified by High-Throughput Single-Cell Sequencing of Convalescent Patients’ B Cells. Cell 2020, 182, 73–84.e16. [Google Scholar] [CrossRef]
- Brouwer, P.J.M.; Caniels, T.G.; van der Straten, K.; Snitselaar, J.L.; Aldon, Y.; Bangaru, S.; Torres, J.L.; Okba, N.M.A.; Claireaux, M.; Kerster, G.; et al. Potent Neutralizing Antibodies from COVID-19 Patients Define Multiple Targets of Vulnerability. Science 2020, 369, 643–650. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Shen, C.; Peng, W.; Li, D.; Zhao, C.; Li, Z.; Li, S.; Bi, Y.; Yang, Y.; et al. A Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding to Its Receptor ACE2. Science 2020, 368, 1274–1278. [Google Scholar] [CrossRef]
- Kreye, J.; Reincke, S.M.; Kornau, H.-C.; Sánchez-Sendin, E.; Corman, V.M.; Liu, H.; Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.-C.D.; et al. A Therapeutic Non-Self-Reactive SARS-CoV-2 Antibody Protects from Lung Pathology in a COVID-19 Hamster Model. Cell 2020, 183, 1058–1069.e19. [Google Scholar] [CrossRef]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in Humanized Mice and Convalescent Humans Yield a SARS-CoV-2 Antibody Cocktail. Science 2020, 369, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Brown-Augsburger, P.L.; Corbett, K.S.; Westendorf, K.; Davies, J.; Cujec, T.P.; Wiethoff, C.M.; Blackbourne, J.L.; Heinz, B.A.; Foster, D.; et al. LY-CoV555, a Rapidly Isolated Potent Neutralizing Antibody, Provides Protection in a Non-Human Primate Model of SARS-CoV-2 Infection. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zost, S.J.; Gilchuk, P.; Case, J.B.; Binshtein, E.; Chen, R.E.; Nkolola, J.P.; Schäfer, A.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; et al. Potently Neutralizing and Protective Human Antibodies against SARS-CoV-2. Nature 2020, 584, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Luscombe, N.M.; Thornton, J.M. Protein-DNA Interactions: Amino Acid Conservation and the Effects of Mutations on Binding Specificity. J. Mol. Biol. 2002, 320. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
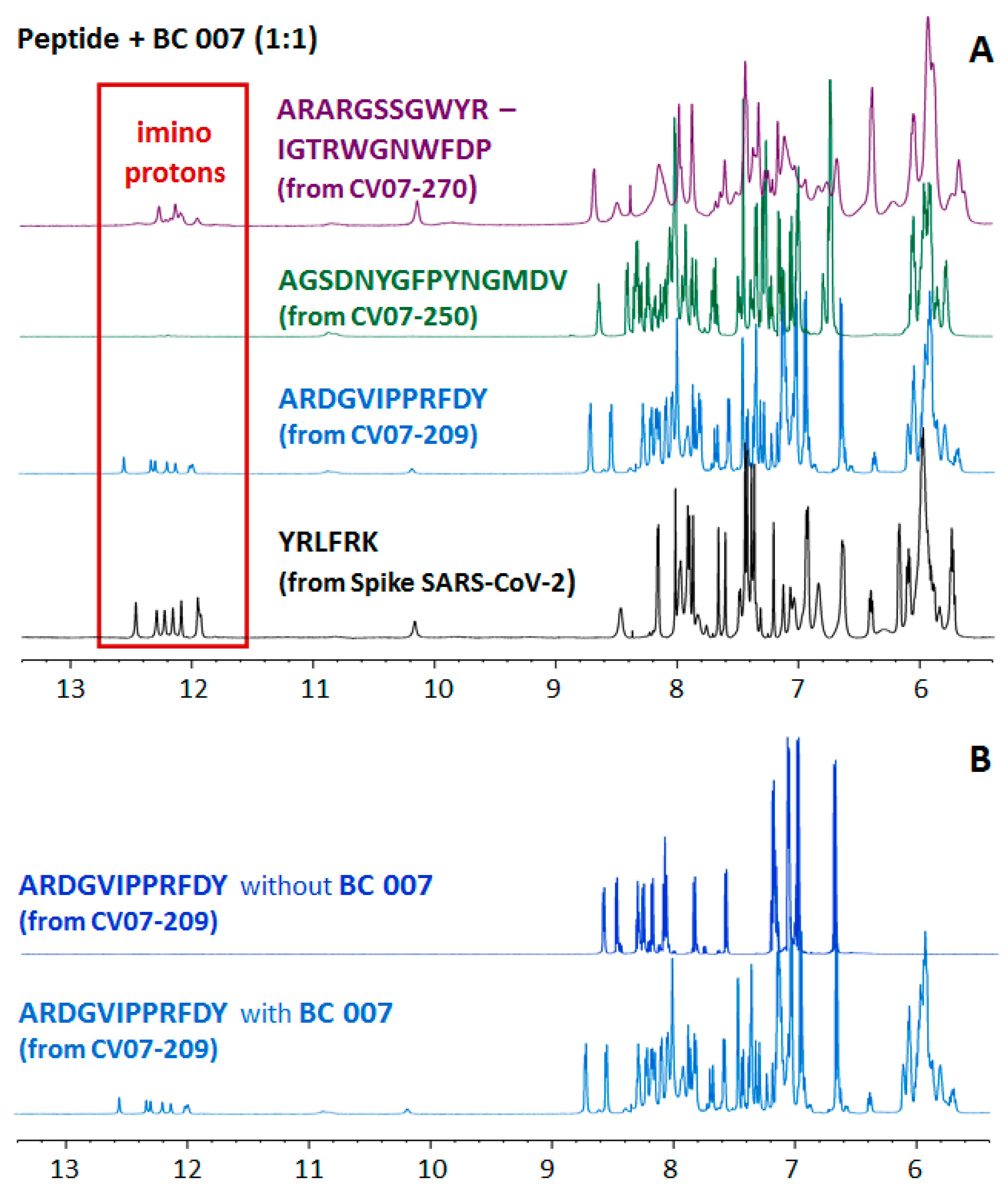
| Clone Name | Cross-Reactive [Reference] | PDB No. | Sections of CDR-H3 | Binding of Sequence Sections Onto BC 007 |
|---|---|---|---|---|
| CV07-250 | no [21] | 6XKQ | AGSDNYGFPYNGMDV | no |
| CV07-209 | no [21] | n.a. | ARDGVIPPRFDY | no |
| CV07-270 | yes [21] | 6XKP | ARARGSSGWYRIGTRWGNWFDP | yes |
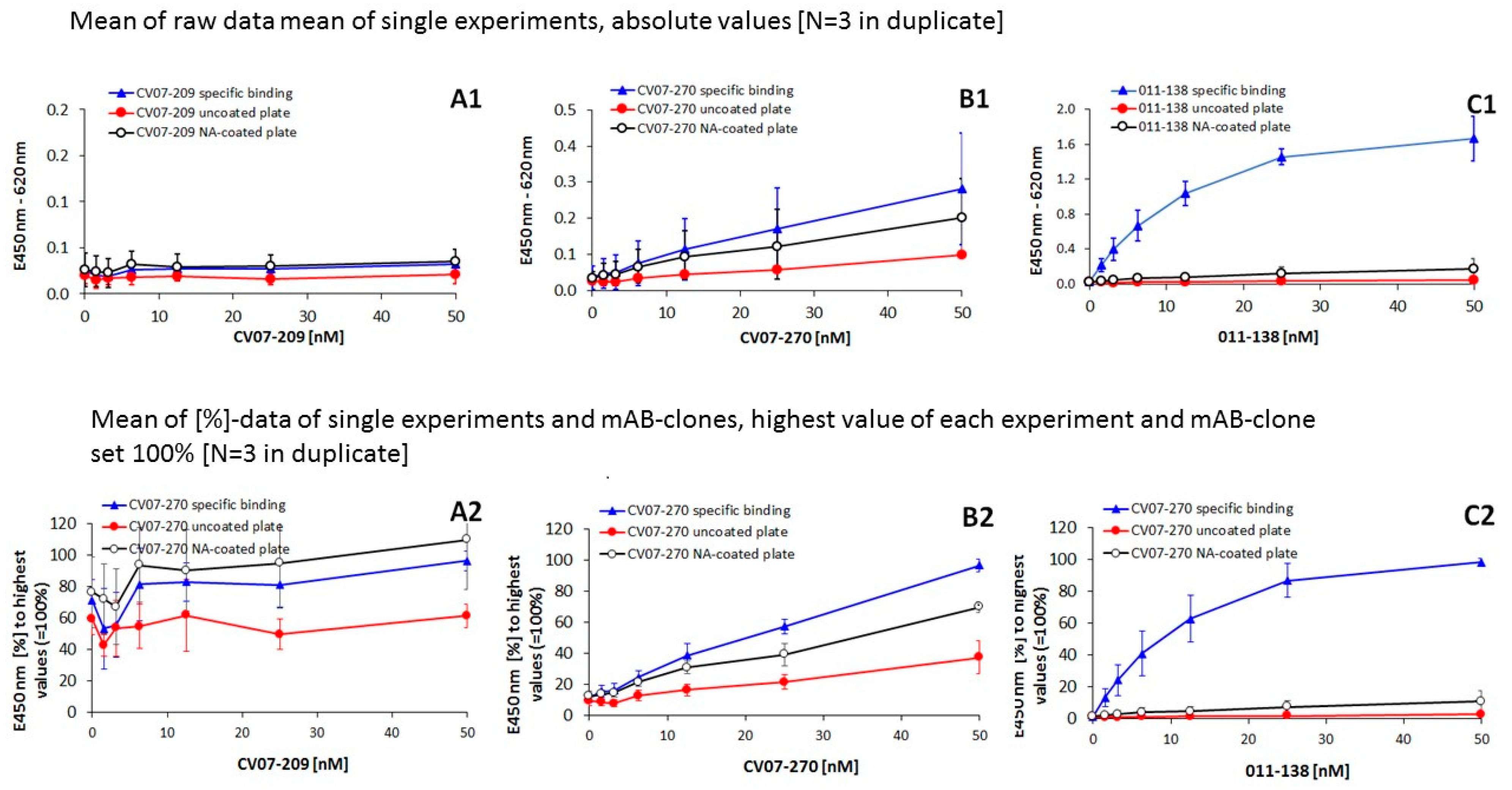
| Concentration mAB Clone | 50 (nM) | 25 (nM) | 12.5 (nM) | 6.25 (nM) | 3.125 (nM) | 1.56 (nM) | |
|---|---|---|---|---|---|---|---|
| Condition vs. Condition | |||||||
| CV07-209 | |||||||
| Figure 3(A1) | specif/uncoated | n.s. | * | n.s. | n.s. | n.s. | n.s. |
| uncoated/NA-coated | n.s. | * | n.s. | n.s. | n.s. | n.s. | |
| Figure 3(A2) | specif/uncoated | *** | ** | n.s. | * | n.s. | n.s. |
| uncoated/NA-coated | * | ** | n.s. | * | n.s. | * | |
| CV07-270 | |||||||
| Figure 3(B1) | specif/uncoated | * | * | * | n.s | n.s | n.s |
| uncoated/NA-coated | * | n.s. | n.s. | n.s. | n.s. | n.s. | |
| Figure 3(B2) | specif/uncoated | *** | *** | *** | *** | ** | ** |
| uncoated/NA-coated | *** | ** | *** | *** | *** | ** | |
| 011-138 | |||||||
| Figure 3(C1) | specif/uncoated | *** | *** | *** | *** | *** | *** |
| uncoated/NA-coated | ** | ** | * | * | * | n.s. | |
| Figure 3(C2) | specif/uncoated | *** | *** | *** | *** | *** | ** |
| uncoated/NA-coated | * | * | * | n.s. | n.s. | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haberland, A.; Krylova, O.; Nikolenko, H.; Göttel, P.; Dallmann, A.; Müller, J.; Weisshoff, H. Aptamer BC 007’s Affinity to Specific and Less-Specific Anti-SARS-CoV-2 Neutralizing Antibodies. Viruses 2021, 13, 932. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050932
Haberland A, Krylova O, Nikolenko H, Göttel P, Dallmann A, Müller J, Weisshoff H. Aptamer BC 007’s Affinity to Specific and Less-Specific Anti-SARS-CoV-2 Neutralizing Antibodies. Viruses. 2021; 13(5):932. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050932
Chicago/Turabian StyleHaberland, Annekathrin, Oxana Krylova, Heike Nikolenko, Peter Göttel, Andre Dallmann, Johannes Müller, and Hardy Weisshoff. 2021. "Aptamer BC 007’s Affinity to Specific and Less-Specific Anti-SARS-CoV-2 Neutralizing Antibodies" Viruses 13, no. 5: 932. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050932