
The Emergence of SARS-CoV-2 within the Dog Population in Croatia: Host Factors and Clinical Outcome

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Methods and Seroprevalence
3.2. Predisposing Factors
3.3. Clinical Manifestation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, 218–225. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental infection of domestic dogs and cats with SARS-CoV-2: Pathogenesis, transmission, and response to reexposure in cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef]
- Sit, T.H.; Brackman, C.J.; Ip, S.M.; Tam, K.W.; Law, P.Y.; To, E.M.; Veronica, Y.T.; Sims, L.D.; Tsang, D.N.; Chu, D.K.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Stevanovic, V.; Vilibic-Cavlek, T.; Tabain, I.; Benvin, I.; Kovac, S.; Hruskar, Z.; Mauric, M.; Milasincic, L.; Antolasic, L.; Skrinjaric, A.; et al. Seroprevalence of SARS-CoV-2 infection among pet animals in Croatia and potential public health impact. Transbound. Emerg. Dis. 2020, 00, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.I.; Elia, G.; Grassi, A.; Giordano, A.; Desario, C.; Medardo, M.; Smith, S.L.; Anderson, E.R.; Prince, T.; Patterson, G.T. Evidence of exposure to SARS-CoV-2 in cats and dogs from households in Italy. Nat. Commun. 2020, 11, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.; Rosolen, B.; Krafft, E.; Becquart, P.; Elguero, E.; Vratskikh, O.; Denolly, S.; Boson, B.; Vanhomwegen, J.; Gouilh, M.A.; et al. High prevalence of SARS-CoV-2 antibodies in pets from COVID-19+ households. One Health 2021, 11, 100192. [Google Scholar] [CrossRef]
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-acute COVID-19 syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef]
- Moritz, C.P.; Tholance, Y.; Lassablière, F.; Camdessanché, J.P.; Antoine, J.C. Reducing the risk of misdiagnosis of indirect ELISA by normalizing serum-specific background noise: The example of detecting anti-FGFR3 autoantibodies. J. Immunol. Methods 2019, 466, 52–56. [Google Scholar] [CrossRef]
- Priestnall, S.L.; Brownlie, J.; Dubovi, E.J.; Erles, K. Serological prevalence of canine respiratory coronavirus. Vet. Microbiol. 2006, 115, 43–53. [Google Scholar] [CrossRef]
- More, G.D.; Dunowska, M.; Acke, E.; Cave, N.J. A serological survey of canine respiratory coronavirus in New Zealand. N. Z. Vet. J. 2020, 68, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Koronavirus—Statistički Pokazatelji za Hrvatsku i EU (Coronavirus—Statistical Indicators for Croatia and the EU). Available online: https://www.koronavirus.hr (accessed on 16 May 2021). (In Croatian).
- Parker, H.G.; Dreger, D.L.; Rimbault, M.; Davis, B.W.; Mullen, A.B.; Carpintero-Ramirez, G.; Ostrander, E.A. Genomic analyses reveal the influence of geographic origin, migration, and hybridisation on modern dog breed development. Cell. Rep. 2017, 19, 697–708. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, D.G.; Jackson, C.; Guy, J.H.; Church, D.B.; McGreevy, P.D.; Thomson, P.C.; Brodbelt, D.C. Epidemiological associations between brachycephaly and upper respiratory tract disorders in dogs attending veterinary practices in England. Canine Genet. Epidemiol. 2015, 2, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packer, R.M.A.; Hendricks, A.; Tivers, M.S.; Burn, C.C. Impact of facial conformation on canine health: Brachycephalic obstructive airway syndrome. PLoS ONE 2015, 10, e0137496. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhang, H.; Gao, J.; Huang, K.; Yang, Y.; Hui, X.; He, X.; Li, C.; Gong, W.; Zhang, Y.; et al. A serological survey of SARS-CoV-2 in cat in Wuhan. Emerg. Microbes Infec. 2020, 9, 2013–2019. [Google Scholar] [CrossRef]
- Perera, R.A.; Mok, C.K.; Tsang, O.T.; Lv, H.; Ko, R.L.; Wu, N.C.; Yuan, M.; Leung, W.S.; Chan, J.M.; Chik, T.S.; et al. Serological assays for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Euro Surveill. 2020, 25, 2000421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Schuurman, N.; Li, W.; Wang, C.; Smit, L.A.; Broens, E.M.; Wagenaar, J.A.; van Kuppeveld, F.J.; Bosch, B.J.; Egberink, H.; et al. Serologic screening of severe acute respiratory syndrome coronavirus 2 infection in cats and dogs during first coronavirus disease wave, the Netherlands. Emerg. Infect. Dis. 2021, 27, 1362–1370. [Google Scholar] [CrossRef]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S.; et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef]
- Suthar, M.S.; Zimmerman, M.G.; Kauffman, R.C.; Mantus, G.; Linderman, S.L.; Hudson, W.H.; Vanderheiden, A.; Nyhoff, L.; Davis, C.W.; Adekunle, O.; et al. Rapid generation of neutralizing antibody responses in COVID-19 patients. Cell. Rep. Med. 2020, 1, 100040. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Stevanovic, V.; Ilic, M.; Barbic, L.; Capak, K.; Tabain, I.; Krleza, J.L.; Ferenc, T.; Hruskar, Z.; Kaliterna, V.; et al. SARS-CoV-2 Seroprevalence and Neutralizing Antibody Response after the First and Second COVID-19 Pandemic Wave in Croatia. Pathogens 2020, 10, 774. [Google Scholar] [CrossRef]
- Okba, N.M.; Müller, M.A.; Li, W.; Wang, C.; GeurtsvanKessel, C.H.; Corman, V.M.; Lamers, M.M.; Sikkema, R.S.; de Bruin, E.; Chandler, F.D.; et al. Severe acute respiratory syndrome coronavirus 2-specific antibody responses in coronavirus disease 2019 patients. Emerg. Infect. Dis. 2020, 26, 1478–1488. [Google Scholar] [CrossRef]
- Decaro, N.; Balboni, A.; Bertolotti, L.; Martino, P.A.; Mazzei, M.; Mira, F.; Pagnini, U. SARS-CoV-2 infection in dogs and cats: Facts and speculations. Front. Vet. Sci. 2021, 8, 80. [Google Scholar] [CrossRef]
- Martina, B.E.; Haagmans, B.L.; Kuiken, T.; Fouchier, R.A.; Rimmelzwaan, G.F.; Van Amerongen, G.; Peiris, J.M.; Lim, W.; Osterhaus, A.D. Virology: SARS virus infection of cats and ferrets. Nature 2003, 425, 915. [Google Scholar] [CrossRef]
- Michelitsch, A.; Hoffmann, D.; Wernike, K.; Beer, M. Occurrence of antibodies against SARS-CoV-2 in the domestic cat population of Germany. Vaccines 2020, 8, 772. [Google Scholar] [CrossRef]
- Li, X. Cats under the shadow of the SARS-CoV-2 pandemic. Transbound. Emerg. Dis. 2020, 67, 1416–1417. [Google Scholar] [CrossRef]
- Hamer, S.A.; Pauvolid-Corrêa, A.; Zecca, I.B.; Davila, E.; Auckland, L.D.; Roundy, C.M.; Tang, W.; Torchetti, M.; Killian, M.L.; Jenkins-Moore, M.; et al. Natural SARS-CoV-2 infections, including virus isolation, among serially tested cats and dogs in households with confirmed human COVID-19 cases in Texas, USA. bioRxiv 2020. [Google Scholar] [CrossRef]
- Grijalva, C.G.; Rolfes, M.A.; Zhu, Y.; McLean, H.Q.; Hanson, K.E.; Belongia, E.A.; Halasa, N.B.; Kim, A.; Reed, C.; Fry, A.M.; et al. Transmission of SARS-COV-2 infections in households—Tennessee and Wisconsin, April–September 2020. Morb. Mortal. Wkly. Rep. 2020, 69, 1631. [Google Scholar] [CrossRef]
- Yi, B.; Fen, G.; Cao, D.; Cai, Y.; Qian, L.; Li, W.; Wen, Z.; Sun, X. Epidemiological and clinical characteristics of 214 families with COVID-19 in Wuhan, China. Int. J. Infect. Dis. 2021, 105, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Laidoudi, Y.; Sereme, Y.; Medkour, H.; Watier-Grillot, S.; Scandola, P.; Ginesta, J.; Andréof, V.; Labarde, C.; Comtet, L.; Pourquier, P.; et al. SARS-CoV-2 antibodies seroprevalence in dogs from France using ELISA and an automated western blotting assay. Méditerranée Infection 2021. [Google Scholar] [CrossRef]
- Abate, B.B.; Kassie, A.M.; Kassaw, M.W.; Aragie, T.G.; Masresha, S.A. Sex difference in coronavirus disease (COVID-19): A systematic review and meta-analysis. BMJ Open 2020, 10, 040129. [Google Scholar] [CrossRef]
- Stall, N.M.; Wu, W.; Lapointe-Shaw, L.; Fisman, D.N.; Giannakeas, V.; Hillmer, M.P.; Rochon, P.A. Sex-specific differences in COVID-19 testing, cases and outcomes: A population-wide study in Ontario, Canada. J. Am. Geriatr. Soc. 2020, 68, 2188–2191. [Google Scholar] [CrossRef] [PubMed]
- Gargaglioni, L.H.; Marques, D.A. Let’s talk about sex in the context of COVID-19. J. App. Physiol. 2020, 128, 1533–1538. [Google Scholar] [CrossRef] [PubMed]
- Peckham, H.; de Gruijter, N.M.; Raine, C.; Radziszewska, A.; Ciurtin, C.; Wedderburn, L.R.; Rosser, E.C.; Webb, K.; Deakin, C.T. Male sex identified by global COVID-19 meta-analysis as a risk factor for death and ITU admission. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, A.J.; Li, X. Sex-dependent immune response and lethality of COVID-19. Stem Cell Res. 2021, 50, 102116. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Talal, N. Sex hormones and the immune system—Part 2: Animal data. Baillieres Clin. Rheumatol. 1990, 4, 13–31. [Google Scholar] [CrossRef]
- Kissick, H.T.; Sanda, M.G.; Dunn, L.K.; Pellegrini, K.L.; On, S.T.; Noel, J.K.; Arredouani, M.S. Androgens alter T-cell immunity by inhibiting T-helper 1 differentiation. Proc. Natl. Acad. Sci. USA 2014, 111, 9887–9892. [Google Scholar] [CrossRef] [Green Version]
- Arredouani, M.S. New insights into androgenic immune regulation. Oncoimmunology 2014, 3, 954968. [Google Scholar] [CrossRef] [Green Version]
- Wallis, L.J.; Range, F.; Müller, C.A.; Serisier, S.; Huber, L.; Virányi, Z. Lifespan development of attentiveness in domestic dogs: Drawing parallels with humans. Front. Psychol. 2014, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Cereda, D.; Tirani, M.; Rovida, F.; Demicheli, V.; Ajelli, M.; Poletti, P.; Trentini, F.; Guzzetta, G.; Marziano, V.; Barone, A.; et al. The Early Phase of the COVID-19 Outbreak in Lombardy, Italy. arXiv 2020, arXiv:2003.09320. [Google Scholar]
- Shim, E.; Tariq, A.; Choi, W.; Lee, Y.; Chowell, G. Transmission potential and severity of COVID-19 in South Korea. Int. J. Infect. Dis. 2020, 93, 339–344. [Google Scholar] [CrossRef]
- Sun, K.; Chen, J.; Viboud, C. Early epidemiological analysis of the coronavirus disease 2019 outbreak based on crowdsourced data: A population-level observational study. Lancet Digit. Health 2020, 2, 201–208. [Google Scholar] [CrossRef]
- Davies, N.G.; Klepac, P.; Liu, Y.; Prem, K.; Jit, M.; Eggo, R.M. Age-dependent effects in the transmission and control of COVID-19 epidemics. Nat. Med. 2020, 26, 1205–1211. [Google Scholar] [CrossRef]
- Mossong, J.; Hens, N.; Jit, M.; Beutels, P.; Auranen, K.; Mikolajczyk, R.; Massari, M.; Salmaso, S.; Tomba, G.S.; Wallinga, J.; et al. Social contacts and mixing patterns relevant to the spread of infectious diseases. PLoS Med. 2008, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- Tennant, B.J.; Gaskell, R.M.; Jones, R.C.; Gaskell, C.J. Studies on the epizootiology of canine coronavirus. Vet. Rec. 1993, 132, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Vin, R.; Leutenegger, C.; Mittel, L.D.; Divers, T.J. Equine coronavirus: An emerging enteric virus of adult horses. Equine Vet. Educ. 2016, 28, 216–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Jabeen, N.; Raza, F.; Shabbir, S.; Baig, A.A.; Amanullah, A.; Aziz, B. Structural variations in human ACE2 may influence its binding with SARS-CoV-2 spike protein. J. Med. Virol. 2020, 92, 1580–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Li, L.; Feng, Z.; Wan, S.; Huang, P.; Sun, X.; Wen, F.; Huang, X.; Ning, G.; Wang, W. Comparative genetic analysis of the novel coronavirus (2019-nCoV/SARS-CoV-2) receptor ACE2 in different populations. Cell Discov. 2020, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Oran, D.P.; Topol, E.J. The proportion of SARS-CoV-2 infections that are asymptomatic: A systematic Review. Ann. Intern. Med. 2020, 173, 362–367. [Google Scholar] [CrossRef]
- Gupta, A.; Madhavan, M.V.; Sehgal, K.; Nair, N.; Mahajan, S.; Sehrawat, T.S.; Bikdeli, B.; Ahluwalia, N.; Ausiello, J.C.; Wan, E.Y.; et al. Extrapulmonary manifestations of COVID-19. Nat. Med. 2020, 26, 1017–1032. [Google Scholar] [CrossRef]
- Carfi, A.; Bernabei, R.; Landi, F. Post-Acute Care Study Group. Persistent symptoms in patients after acute COVID-19. J. Am. Med. Assoc. 2020, 324, 603–605. [Google Scholar] [CrossRef]
- Chopra, V.; Flanders, S.A.; O’Malley, M.; Malani, A.N.; Prescott, H.C. Sixty-day outcomes among patients hospitalised with COVID-19. Ann. Intern. Med. 2020. [Google Scholar] [CrossRef]
- Garrigues, E.; Janvier, P.; Kherabi, Y.; Le Bot, A.; Hamon, A.; Gouze, H.; Doucet, L.; Berkani, S.; Oliosi, E.; Mallart, E.; et al. Post-discharge persistent symptoms and health-related quality of life after hospitalisation for COVID-19. J. Infect. 2020, 81, 4–6. [Google Scholar] [CrossRef]
- Hosie, M.J.; Epifano, I.; Herder, V.; Orton, R.; Stevenson, A.; Johnson, N.; MacDonald, E.; Dunbar, D.; McDonald, M.; Howie, F.; et al. Respiratory disease in cats associated with human-to-cat transmission of SARS-CoV-2 in the UK. bioRxiv 2020. [Google Scholar] [CrossRef]
- Garigliany, M.; Van Laere, A.S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; Van Der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 natural transmission from human to cat, Belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069. [Google Scholar] [CrossRef]
- Ferasin, L.; Fritz, M.; Ferasin, H.; Becquart, P.; Legros, V.; Leroy, E.M. Myocarditis in naturally infected pets with the British variant of COVID-19. bioRxiv 2021. [Google Scholar] [CrossRef]
- Moriguchi, T.; Harii, N.; Goto, J.; Harada, D.; Sugawara, H.; Takamino, J.; Ueno, M.; Sakata, H.; Kondo, K.; Myose, N.; et al. A first case of meningitis/encephalitis associated with SARS-Coronavirus-2. Int. J. Infect. Dis. 2020, 94, 55–58. [Google Scholar] [CrossRef]
- Domingues, R.B.; Mendes-Correa, M.C.; de Moura Leite, F.B.V.; Sabino, E.C.; Salarini, D.Z.; Claro, I.; Santos, D.W.; de Jesus, J.G.; Ferreira, N.E.; Romano, C.M.; et al. First case of SARS-COV-2 sequencing in cerebrospinal fluid of a patient with suspected demyelinating disease. J. Neurol. 2020, 267, 3154–3156. [Google Scholar] [CrossRef] [PubMed]
- Pezzini, A.; Padovani, A. Lifting the mask on neurological manifestations of COVID-19. Nat. Rev. Neurol. 2020, 16, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Ellul, M.A.; Benjamin, L.; Singh, B.; Lant, S.; Michael, B.D.; Easton, A.; Kneen, R.; Defres, S.; Sejvar, J.; Solomon, T. Neurological associations of COVID-19. Lancet Neurol. 2020, 19, 767–783. [Google Scholar] [CrossRef]
- Nersesjan, V.; Amiri, M.; Lebech, A.M.; Roed, C.; Mens, H.; Russell, L.; Fonsmark, L.; Berntsen, M.; Sigurdsson, S.T.; Carlsen, J.; et al. Central and peripheral nervous system complications of COVID-19: A prospective tertiary center cohort with 3-month follow-up. J. Neurol. 2021, 1–19. [Google Scholar] [CrossRef]
- Mohamed, M.S.; Moulin, T.C.; Schiöth, H.B. Sex differences in COVID-19: The role of androgens in disease severity and progression. Endocrine 2020, 71, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Abdool Karim, S.S.; de Oliveira, T. New SARS-CoV-2 variants—clinical, public health, and vaccine implications. N. Engl. J. Med. 2021, 384, 1866–1868. [Google Scholar] [CrossRef]
- O’Toole, Á.; Hill, V.; Pybus, O.G.; Watts, A.; Bogoch, I.I.; Khan, K.; Messina, J.P.; COVID-19 Genomics UK (COG-UK) Consortium; Network for Genomic Surveillance in South Africa (NGS-SA); Brazil-UK CADDE Genomic Network; et al. Tracking the international spread of SARS-CoV-2 lineages B.1.1.7 and B.1.351/501Y-V2. Wellcome Open Res. 2021, 6, 121. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
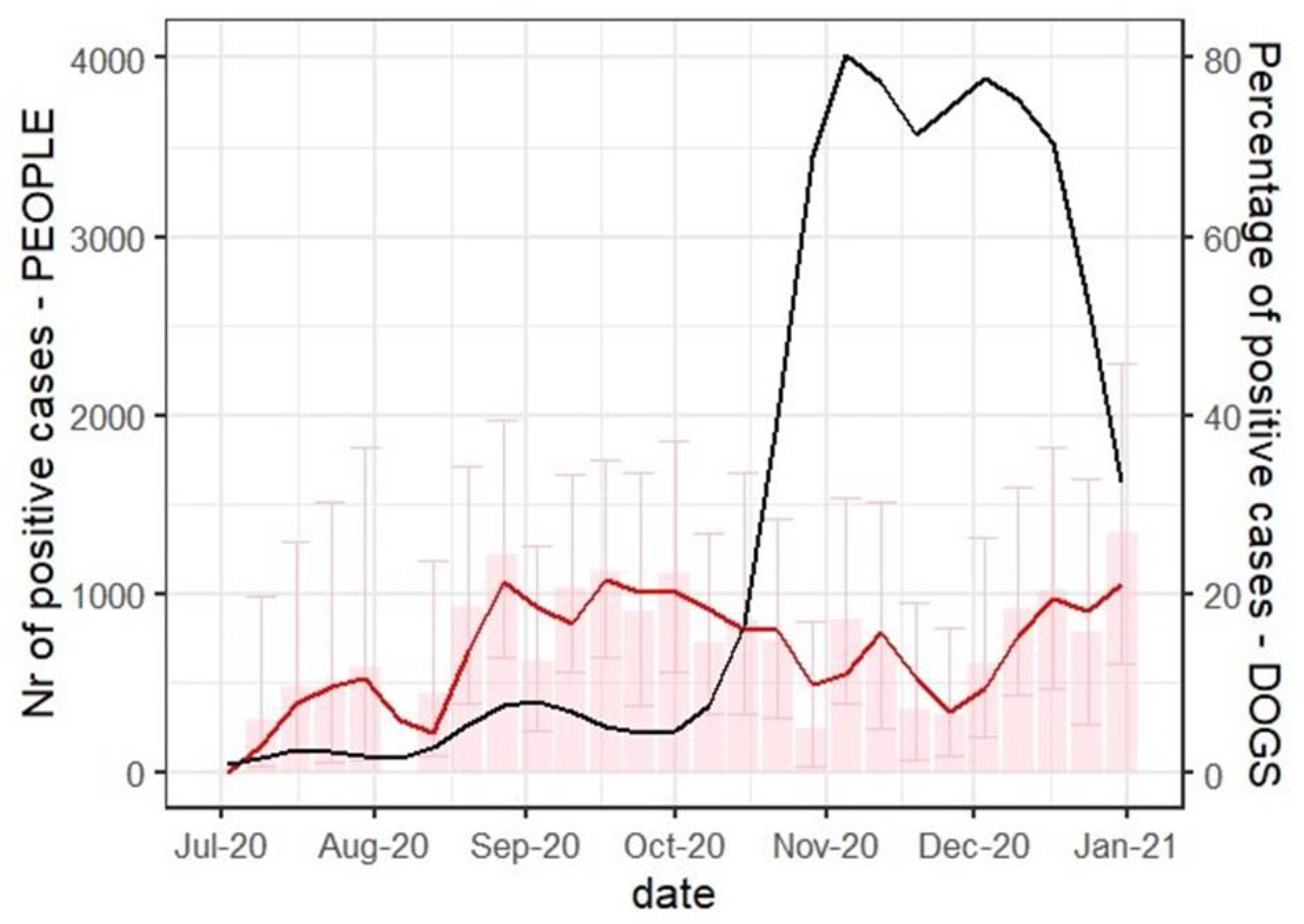
| Number of Tested Samples | ELISA a Positive N (%) | Seroprevalence 95% CI b (%) | OR c | OR c 95% CI | p | |
|---|---|---|---|---|---|---|
| July | 126 | 9 (7.14) | 3.32–13.13 | 0.08 | 0.04–0.14 | - |
| August | 152 | 23 (15.13) | 9.84–21.83 | 2.32 | 1.06–5.47 | 0.042 |
| September | 233 | 46 (19.74) | 14.83–25.44 | 3.2 | 1.58–7.2 | 0.002* |
| October | 191 | 24 (12.57) | 8.22–18.12 | 1.87 | 0.86–4.38 | 0.126 |
| November | 200 | 24 (12) | 7.84–17.33 | 1.77 | 0.82–4.15 | 0.161 |
| December | 167 | 31 (18.56) | 12.97–25.3 | 2.96 | 1.41–6.84 | 0.006 * |
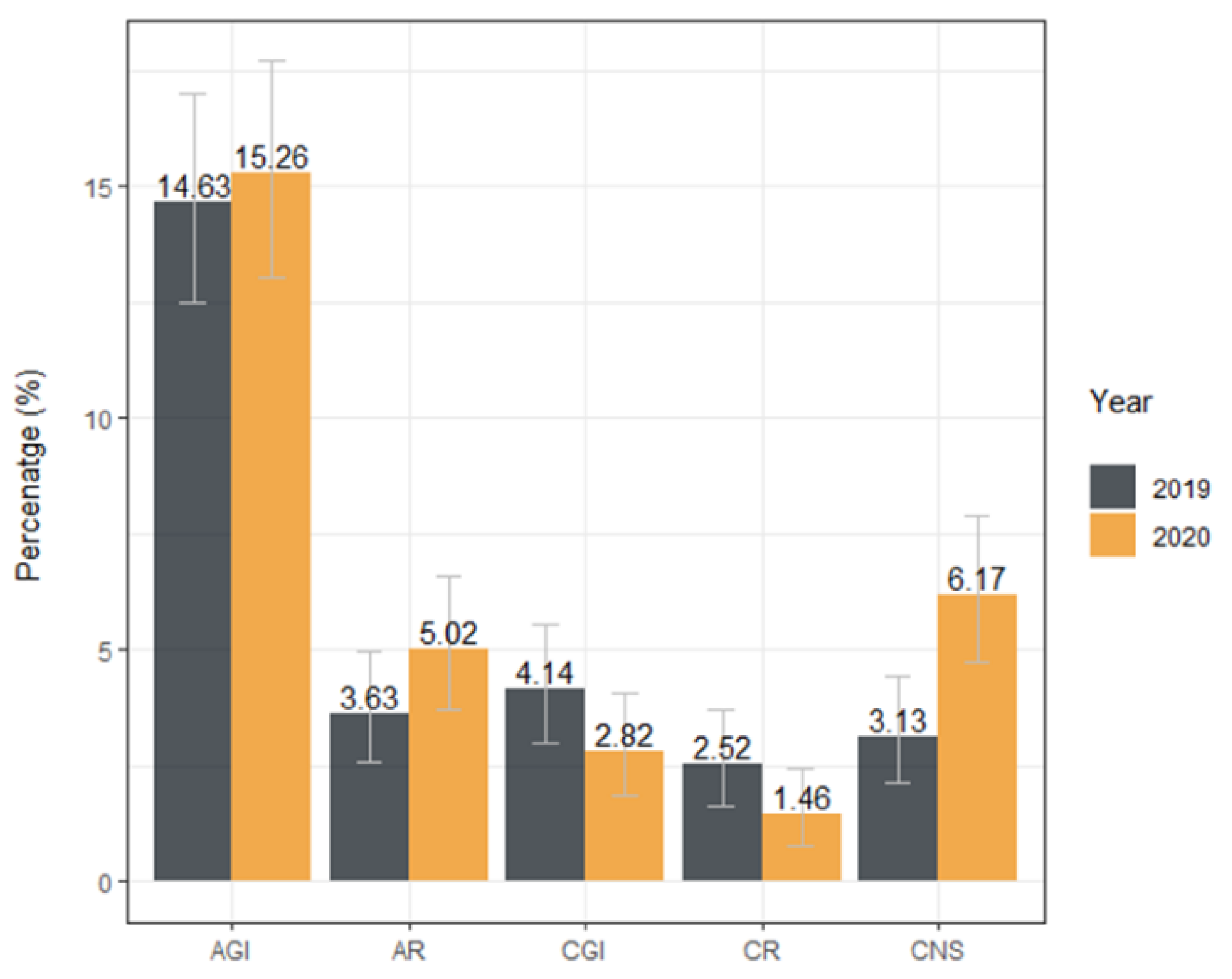
| Diagnosis | Number of Tested Samples | Number of ELISA a Positive Samples (%) | Seroprevalence 95% CI b | OR c | OR 95%CI | p |
|---|---|---|---|---|---|---|
| Surgical | 269 | 37 (13.75) | 9.87–18.46 | 0.16 | 0.11–0.22 | - |
| Acute GI d | 146 | 23 (15.75) | 10.26–22.69 | 1.17 | 0.66–2.05 | 0.58 |
| Acute respiratory | 48 | 8 (16.67) | 7.48–30.22 | 1.25 | 0.51–2.77 | 0.6 |
| Cardio | 57 | 8 (14.04) | 6.26–25.79 | 1.02 | 0.42–2.23 | 0.96 |
| Chronic GI d | 27 | 4 (14.81) | 4.19–33.73 | 1.09 | 0.31–3.03 | 0.88 |
| Chronic respiratory | 14 | 1 (7.14) | 0.18–33.87 | 0.48 | 0.03–2.53 | 0.49 |
| CNS e | 59 | 15 (25.42) | 14.98–38.44 | 2.14 | 1.06–4.17 | 0.03* |
| Dermatologic | 77 | 12 (15.58) | 8.32–25.64 | 1.16 | 0.55–2.29 | 0.68 |
| Early infection | 31 | 6 (19.35) | 7.45–37.47 | 1.5 | 0.53–3.71 | 0.40 |
| Endocrine | 18 | 1 (5.56) | 0.14–27.29 | 0.37 | 0.02–1.88 | 0.34 |
| Healthy | 28 | 4 (14.29) | 4.03–32.67 | 1.05 | 0.3–2.9 | 0.94 |
| Malignancy | 70 | 7 (10) | 4.12-19-52 | 0.7 | 0.27–1.55 | 0.41 |
| Ophthalmic | 18 | 1 (5.56) | 0.14–27.29 | 0.37 | 0.02–1.88 | 0.34 |
| Other | 95 | 8 (8.42) | 3.71–15.92 | 0.58 | 0.24–1.23 | 0.18 |
| ELISA a | Seroprevalence % (95% CI b) | Statistical Significance | |||
|---|---|---|---|---|---|
| Positive (n) | Negative (n) | ||||
| Dog | 31 | 136 | 18.56 (12.97–25.3) | ELISA positivity rate dog/human n = 625 χ2e = 0.184 df = 1 p = 0.667 | |
| MNT c | Positive | 16 | NT d | - | |
| Negative | 15 | NT d | - | ||
| Human | 94 | 364 | 20.52 (16.92–24.52) | MNT positive dog/human samples n = 125 χ2e = 4.658 df = 1 p = 0.031 * | |
| MNT c | Positive | 70 | NT d | - | |
| Negative | 24 | NT d | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevanovic, V.; Tabain, I.; Vilibic-Cavlek, T.; Mauric Maljkovic, M.; Benvin, I.; Hruskar, Z.; Kovac, S.; Smit, I.; Miletic, G.; Hadina, S.; et al. The Emergence of SARS-CoV-2 within the Dog Population in Croatia: Host Factors and Clinical Outcome. Viruses 2021, 13, 1430. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081430
Stevanovic V, Tabain I, Vilibic-Cavlek T, Mauric Maljkovic M, Benvin I, Hruskar Z, Kovac S, Smit I, Miletic G, Hadina S, et al. The Emergence of SARS-CoV-2 within the Dog Population in Croatia: Host Factors and Clinical Outcome. Viruses. 2021; 13(8):1430. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081430
Chicago/Turabian StyleStevanovic, Vladimir, Irena Tabain, Tatjana Vilibic-Cavlek, Maja Mauric Maljkovic, Iva Benvin, Zeljka Hruskar, Snjezana Kovac, Iva Smit, Gorana Miletic, Suzana Hadina, and et al. 2021. "The Emergence of SARS-CoV-2 within the Dog Population in Croatia: Host Factors and Clinical Outcome" Viruses 13, no. 8: 1430. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081430