
Development of a Dendrimeric Peptide-Based Approach for the Differentiation of Animals Vaccinated with FlagT4G against Classical Swine Fever from Infected Pigs

 , ,
, ,  ,
, 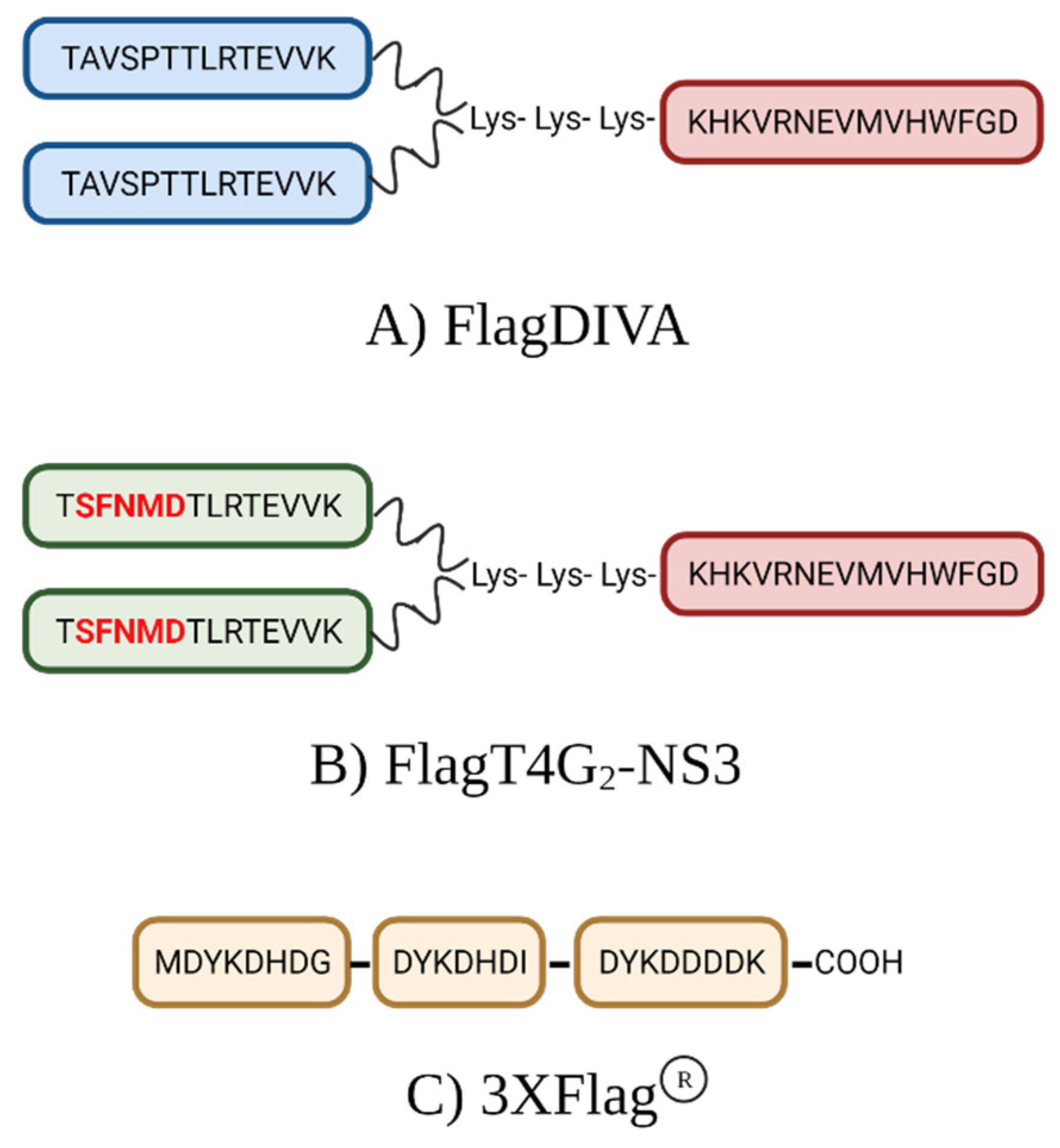{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Synthesis of Dendrimeric Peptides
2.3. Experimental Vaccination
2.4. Neutralizing and CSFV E2-Specific Antibody Detection
2.5. Determination of Humoral Response Elicited by FlagT4G Using Dendrimeric Peptides and Flag® Peptide ELISA
2.6. CSFV Humoral Response Detection by Dendrimeric Peptides
3. Results
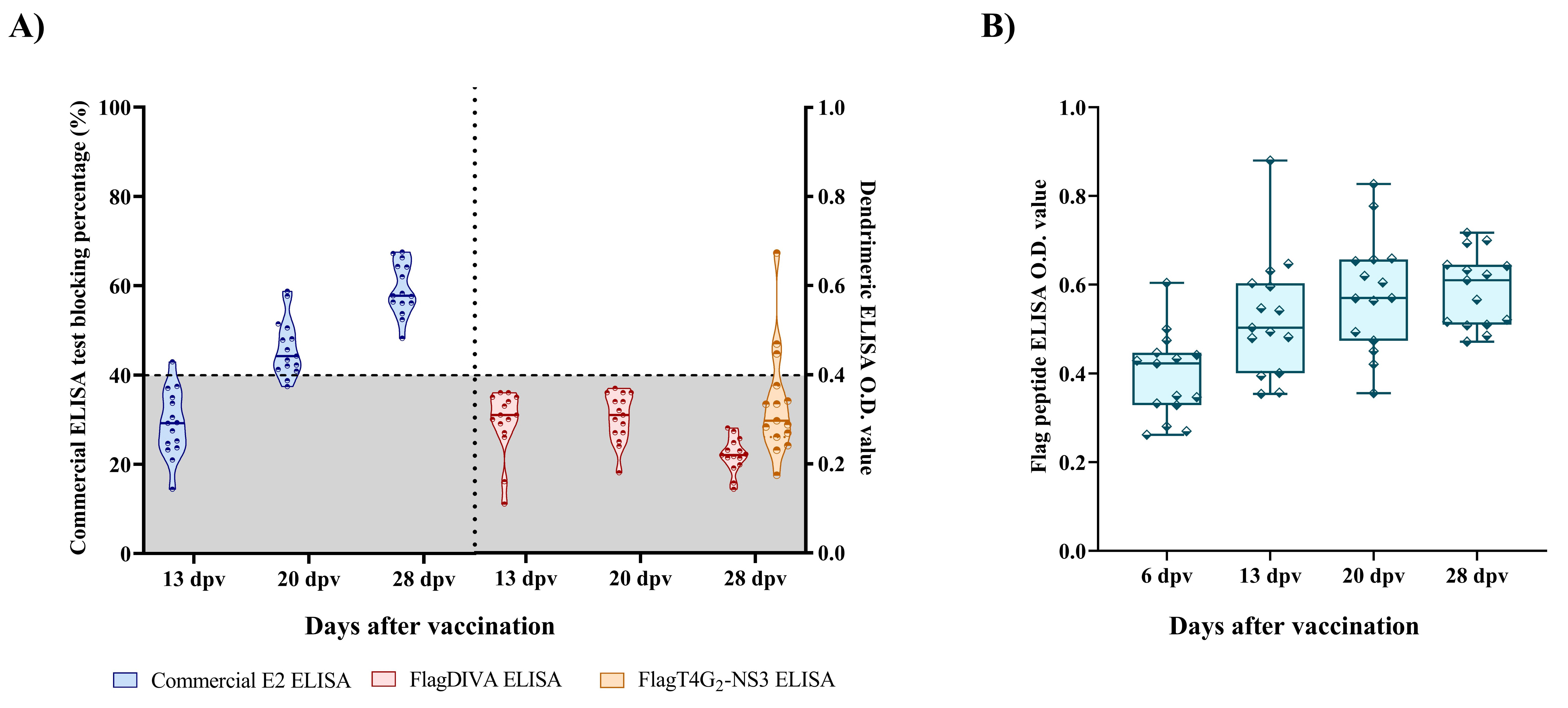
3.1. The Antibody Response Elicited by the FlagT4G Vaccine against CSFV Is Not Detected by the FlagDIVA Peptide
3.2. Comparative Performance between the FlagDIVA ELISA and the Commercial ELISA in the Detection of e2-Csfv Antibody Response
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef] [PubMed]
- Postel, A.; Nishi, T.; Kameyama, K.-I.; Meyer, D.; Suckstorff, O.; Fukai, K.; Becher, P. Reemergence of Classical Swine Fever, Japan, 2018. Emerg. Infect. Dis. 2019, 25, 1228–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blome, S.; Moß, C.; Reimann, I.; König, P.; Beer, M. Classical swine fever vaccines—State-of-the-art. Vet. Microbiol. 2017, 206, 10–20. [Google Scholar] [CrossRef] [PubMed]
- OIE Recognition of the Classical Swine Fever Status of Members; Paris. 2019. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Resolutions/2019/A_R22_CSF_status.pdf (accessed on 22 February 2020).
- EMA Suvaxyn CSF Marker - EMEA/V/C/002757 - R/0006. 2019. Available online: https://www.ema.europa.eu/en/medicines/veterinary/EPAR/suvaxyn-csf-marker (accessed on 7 March 2020).
- Postel, A.; Becher, P. Genetically distinct pestiviruses pave the way to improved classical swine fever marker vaccine candidates based on the chimeric pestivirus concept. Emerg. Microbes Infect. 2020, 9, 2180–2189. [Google Scholar] [CrossRef] [PubMed]
- Holinka, L.; Fernandez-Sainz, I.; Sanford, B.; O’donnell, V.; Gladue, D.; Carlson, J.; Lu, Z.; Risatti, G.; Borca, M. Development of an improved live attenuated antigenic marker CSF vaccine strain candidate with an increased genetic stability. Virology 2014, 471–473, 13–18. [Google Scholar] [CrossRef]
- Holinka, L.G.; O’Donnell, V.; Risatti, G.R.; Azzinaro, P.; Arzt, J.; Stenfeldt, C.; Velazquez-Salinas, L.; Carlson, J.; Gladue, D.; Borca, M.V. Early protection events in swine immunized with an experimental live attenuated classical swine fever marker vaccine, FlagT4G. PLoS ONE 2017, 12, e0177433. [Google Scholar] [CrossRef]
- Hopp, T.P.; Prickett, K.S.; Price, V.L.; Libby, R.T.; March, C.J.; Cerretti, D.P.; Urdal, D.L.; Conlon, P.J. A Short Polypeptide Marker Sequence Useful for Recombinant Protein Identification and Purification. Bio/Technology 1988, 6, 1204–1210. [Google Scholar] [CrossRef]
- Heegaard, P.; Boas, U.; Sorensen, N.S. Dendrimers for Vaccine and Immunostimulatory Uses. A Review. Bioconjugate Chem. 2010, 21, 405–418. [Google Scholar] [CrossRef]
- Mhlwatika, Z.; Aderibigbe, B.A. Application of Dendrimers for the Treatment of Infectious Diseases. Molecules 2018, 23, 2205. [Google Scholar] [CrossRef] [Green Version]
- Blanco, E.; Guerra, B.; de la Torre, B.G.; Defaus, S.; Dekker, A.; Andreu, D.; Sobrino, F. Full protection of swine against foot-and-mouth disease by a bivalent B-cell epitope dendrimer peptide. Antivir. Res. 2016, 129, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Forner, M.; Cañas-Arranz, R.; Defaus, S.; de León, P.; Rodríguez-Pulido, M.; Ganges, L.; Blanco, E.; Sobrino, F.; Andreu, D. Peptide-Based Vaccines: Foot-and-Mouth Disease Virus, a Paradigm in Animal Health. Vaccines 2021, 9, 477. [Google Scholar] [CrossRef] [PubMed]
- Bosnjakovic, A.; Mishra, M.K.; Han, H.J.; Romero, R.; Kannan, R.M. A dendrimer-based immunosensor for improved capture and detection of tumor necrosis factor-α cytokine. Anal. Chim. Acta 2012, 720, 118–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, J.; Dong, C.; Li, Y.; Kou, Q.; Yan, J.; Zhang, L. Ultrasensitive ELISA for the detection of hCG based on assembled gold nanoparticles induced by functional polyamidoamine dendrimers. Anal. Chim. Acta 2018, 1042, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Malvano, F.; Pilloton, R.; Albanese, D. Sensitive Detection of Escherichia coli O157:H7 in Food Products by Impedimetric Immunosensors. Sensors 2018, 18, 2168. [Google Scholar] [CrossRef] [Green Version]
- Tarradas, J.; Monsó, M.; Fraile, L.; de la Torre, B.G.; Muñoz, M.; Rosell, R.; Riquelme, C.; Pérez, L.J.; Nofrarías, M.; Domingo, M.; et al. A T-cell epitope on NS3 non-structural protein enhances the B and T cell responses elicited by dendrimeric constructions against CSFV in domestic pigs. Veter- Immunol. Immunopathol. 2012, 150, 36–46. [Google Scholar] [CrossRef]
- Tarradas, J.; Monsó, M.; Muñoz, M.; Rosell, R.; Fraile, L.; Frías, M.T.; Domingo, M.; Andreu, D.; Sobrino, F.; Ganges, L. Partial protection against classical swine fever virus elicited by dendrimeric vaccine-candidate peptides in domestic pigs. Vaccine 2011, 29, 4422–4429. [Google Scholar] [CrossRef] [PubMed]
- Bohórquez, J.A.; Defaus, S.; Muñoz-González, S.; Perez-Simó, M.; Rosell, R.; Fraile, L.; Sobrino, F.; Andreu, D.; Ganges, L. A bivalent dendrimeric peptide bearing a T-cell epitope from foot-and-mouth disease virus protein 3A improves humoral response against classical swine fever virus. Virus Res. 2017, 238, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Wensvoort, G.; Terpstra, C.; Boonstra, J.; Bloemraad, M.; Zaane, D. Van Production of monoclonal antibodies against swine fever virus and their use in laboratory diagnosis. Vet. Microbiol. 1986, 12, 101–108. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Terpstra, C.; Bloemraad, M.; Gielkens, A.L.J. The neutralizing peroxidase-linked assay for detection of antibody against swine fever virus. Vet. Microbiol. 1984, 9, 113–120. [Google Scholar] [CrossRef]
- Sozzi, E.; Lavazza, A.; Gaffuri, A.; Bencetti, F.C.; Prosperi, A.; Lelli, D.; Chiapponi, C.; Moreno, A. Isolation and Full-Length Sequence Analysis of a Pestivirus from Aborted Lamb Fetuses in Italy. Viruses 2019, 11, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohórquez, J.A.; Sozzi, E.; Wang, M.; Alberch, M.; Abad, X.; Gaffuri, A.; Lelli, D.; Rosell, R.; Pérez, L.J.; Moreno, A.; et al. The new emerging ovine pestivirus can infect pigs and confers strong protection against classical swine fever virus. Transbound. Emerg. Dis. 2021, 00, 1–17. [Google Scholar] [CrossRef]
- Coronado, L.; Bohórquez, J.A.; Muñoz-González, S.; Perez, L.J.; Rosell, R.; Fonseca, O.; Delgado, L.; Perera, C.L.; Frías, M.T.; Ganges, L. Investigation of chronic and persistent classical swine fever infections under field conditions and their impact on vaccine efficacy. BMC Veter- Res. 2019, 15, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronado, L.; Perera, C.; Rios, L.; Frías, M.; Pérez, L. A Critical Review about Different Vaccines against Classical Swine Fever Virus and Their Repercussions in Endemic Regions. Vaccines 2021, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- van Oirschot, J. Vaccinology of classical swine fever: From lab to field. Vet. Microbiol. 2003, 96, 367–384. [Google Scholar] [CrossRef]
- Meuwissen, M.P.; Horst, S.H.; Huirne, R.B.; A Dijkhuizen, A. A model to estimate the financial consequences of classical swine fever outbreaks: Principles and outcomes. Prev. Veter- Med. 1999, 42, 249–270. [Google Scholar] [CrossRef]
- Kortekaas, J.; Vloet, R.P.M.; Weerdmeester, K.; Ketelaar, J.; van Eijk, M.; Loeffen, W.L. Rational design of a classical swine fever C-strain vaccine virus that enables the differentiation between infected and vaccinated animals. J. Virol. Methods 2010, 163, 175–185. [Google Scholar] [CrossRef]
- Terpstra, C.; Wensvoort, G. The protective value of vaccine-induced neutralising antibody titres in swine fever. Veter- Microbiol. 1988, 16, 123–128. [Google Scholar] [CrossRef]
- Jelsma, T.; Post, J.; Born, E.; Segers, R.; Kortekaas, J. Assessing the Protective Dose of a Candidate DIVA Vaccine against Classical Swine Fever. Vaccines 2021, 9, 483. [Google Scholar] [CrossRef]
- Holinka, L.; Fernandez-Sainz, I.; O’Donnell, V.; Prarat, M.; Gladue, D.; Lu, Z.; Risatti, G.; Borca, M. Development of a live attenuated antigenic marker classical swine fever vaccine. Virology 2009, 384, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Sozzi, E.; Bohórquez, J.A.; Alberch, M.; Pujols, J.; Cantero, G.; Gaffuri, A.; Lelli, D.; Rosell, R.; Bensaid, A.; et al. Decrypting the Origin and Pathogenesis in Pregnant Ewes of a New Ovine Pestivirus Closely Related to Classical Swine Fever Virus. Viruses 2020, 12, 775. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Wernike, K.; Reimann, I.; König, P.; Moß, C.; Beer, M. A decade of research into classical swine fever marker vaccine CP7_E2alf (Suvaxyn® CSF Marker): A review of vaccine properties. Vet. Res. 2017, 48, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Bohórquez, J.A.; Hinojosa, Y.; Muñoz-González, S.; Gerber, M.; Coronado, L.; Perera, C.L.; Liniger, M.; Ruggli, N.; Ganges, L. Abrogation of the RNase activity of Erns in a low virulence classical swine fever virus enhances the humoral immune response and reduces virulence, transmissibility, and persistence in pigs. Virulence 2021, 12, 2037–2049. [Google Scholar] [CrossRef] [PubMed]
- Kortekaas, J.; Ketelaar, J.; Vloet, R.P.M.; Loeffen, W.L. Protective efficacy of a Classical swine fever virus C-strain deletion mutant and ability to differentiate infected from vaccinated animals. Vet. Microbiol. 2011, 147, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Bruderer, U.; van de Velde, J.; Frantzen, I.; De Bortoli, F. Discrimination within epitope specific antibody populations against Classical swine fever virus is a new means of differentiating infection from vaccination. J. Immunol. Methods 2015, 420, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Coronado, L.; Rios, L.; Frías, M.T.; Amarán, L.; Naranjo, P.; Percedo, M.I.; Perera, C.L.; Prieto, F.; Fonseca-Rodriguez, O.; Perez, L.J. Positive selection pressure on E2 protein of classical swine fever virus drives variations in virulence, pathogenesis and antigenicity: Implication for epidemiological surveillance in endemic areas. Transbound. Emerg. Dis. 2019, 66, 2362–2382. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bohórquez, J.A.; Defaus, S.; Rosell, R.; Pérez-Simó, M.; Alberch, M.; Gladue, D.P.; Borca, M.V.; Andreu, D.; Ganges, L. Development of a Dendrimeric Peptide-Based Approach for the Differentiation of Animals Vaccinated with FlagT4G against Classical Swine Fever from Infected Pigs. Viruses 2021, 13, 1980. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101980
Bohórquez JA, Defaus S, Rosell R, Pérez-Simó M, Alberch M, Gladue DP, Borca MV, Andreu D, Ganges L. Development of a Dendrimeric Peptide-Based Approach for the Differentiation of Animals Vaccinated with FlagT4G against Classical Swine Fever from Infected Pigs. Viruses. 2021; 13(10):1980. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101980
Chicago/Turabian StyleBohórquez, José Alejandro, Sira Defaus, Rosa Rosell, Marta Pérez-Simó, Mònica Alberch, Douglas P. Gladue, Manuel V. Borca, David Andreu, and Llilianne Ganges. 2021. "Development of a Dendrimeric Peptide-Based Approach for the Differentiation of Animals Vaccinated with FlagT4G against Classical Swine Fever from Infected Pigs" Viruses 13, no. 10: 1980. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101980