
Interactions between the Prophage 919TP and Its Vibrio cholerae Host: Implications of gmd Mutation for Phage Resistance, Cell Auto-Aggregation, and Motility

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
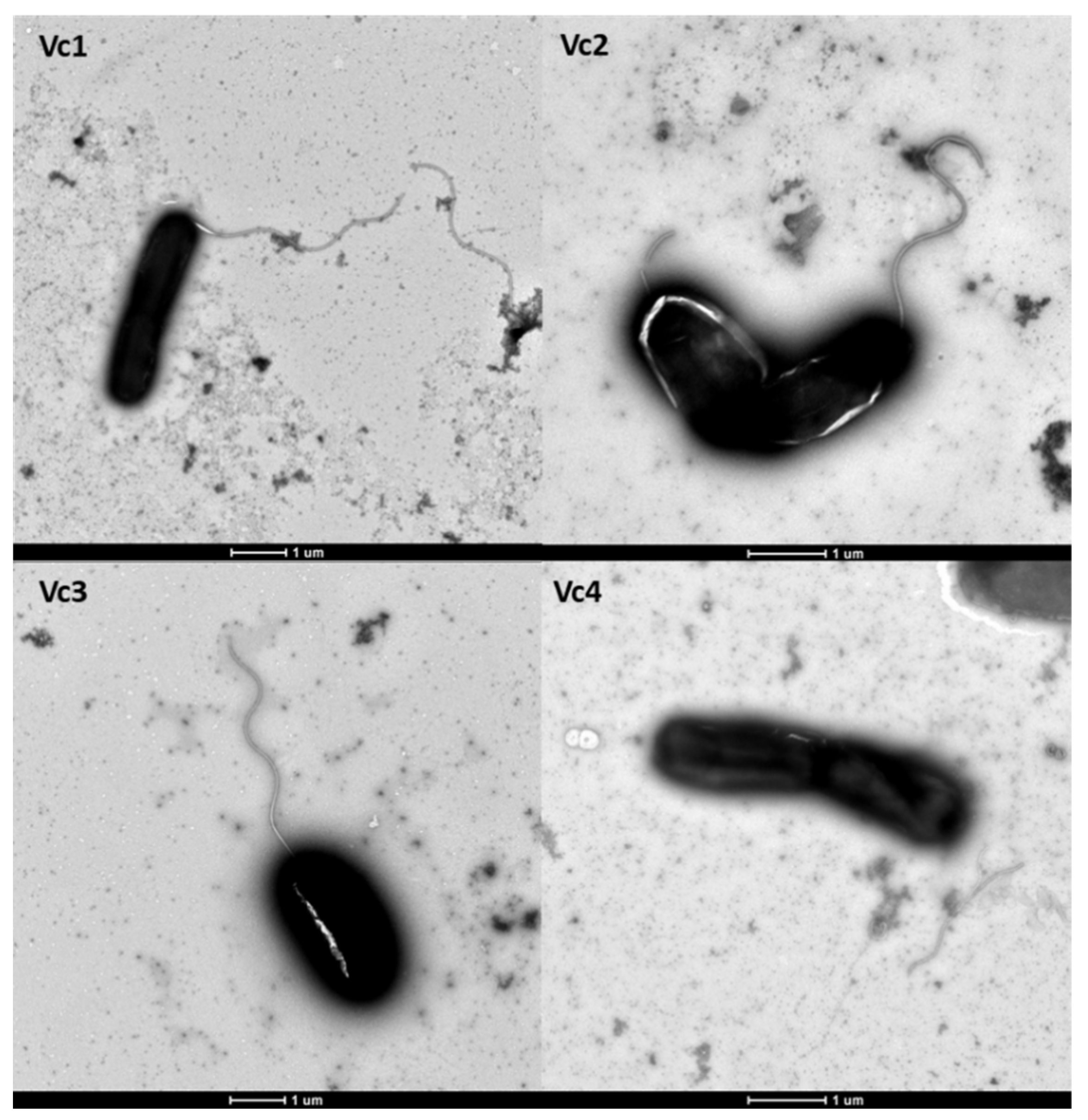
2.1. Vibrio cholerae Strains, Bacteriophages, Plasmids, Oligos, and Growth Conditions
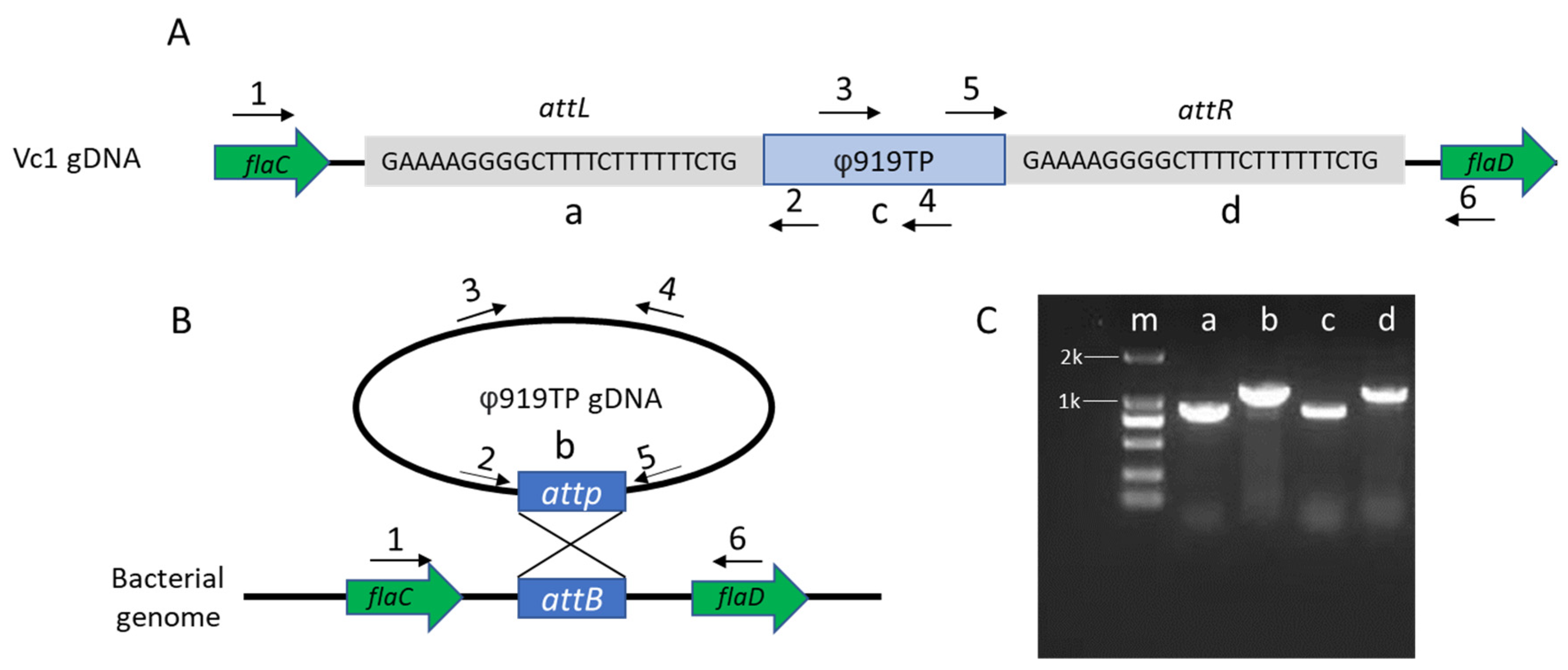
2.2. Mapping of the Phage Attachment Site on the Vibrio Chromosome
2.3. DNA Manipulations
2.4. Isolation of Phage 919TP cI--Resistant Variants
2.5. Adsorption Assays and Efficiency of Plating (EOP)
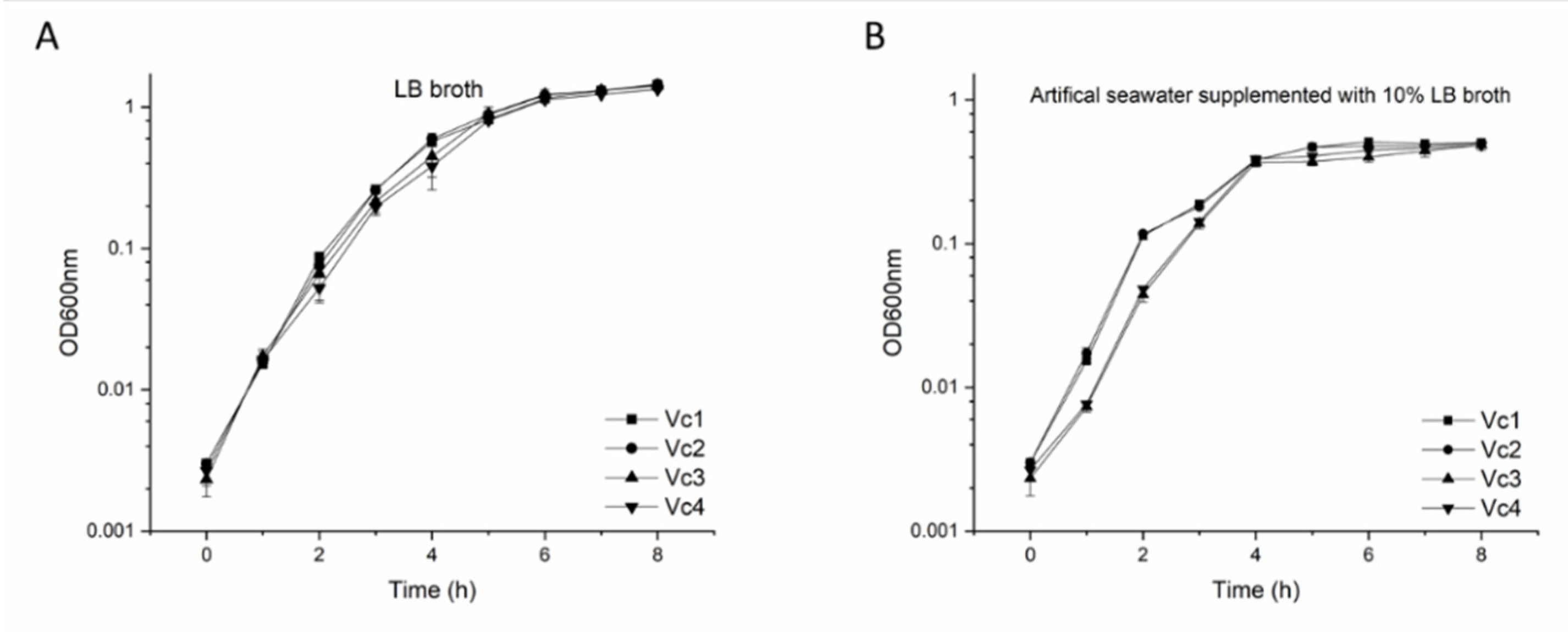
2.6. Growth Curves of V. cholerae Strains
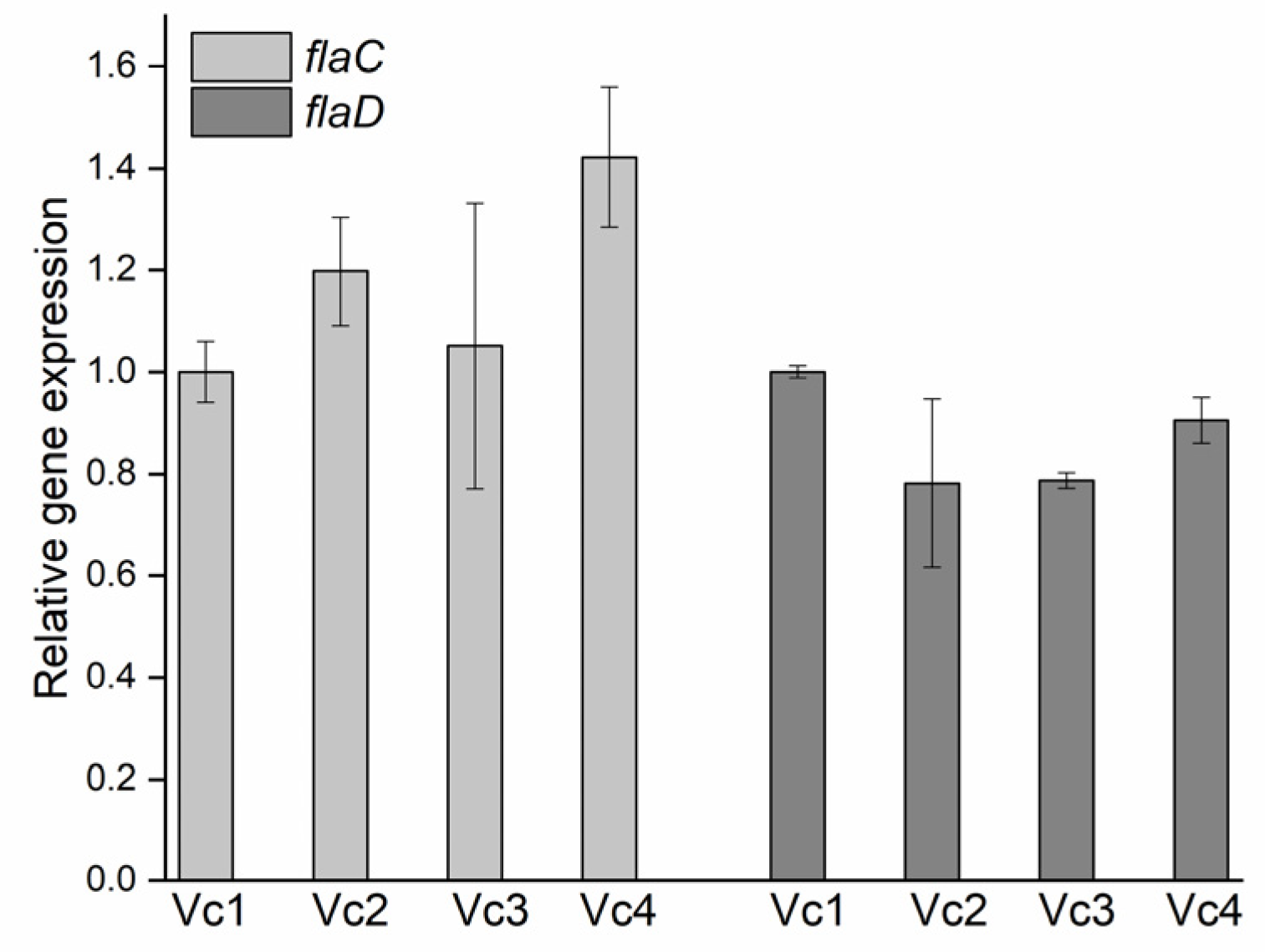
2.7. Quantification of flaC and flaD mRNA Levels
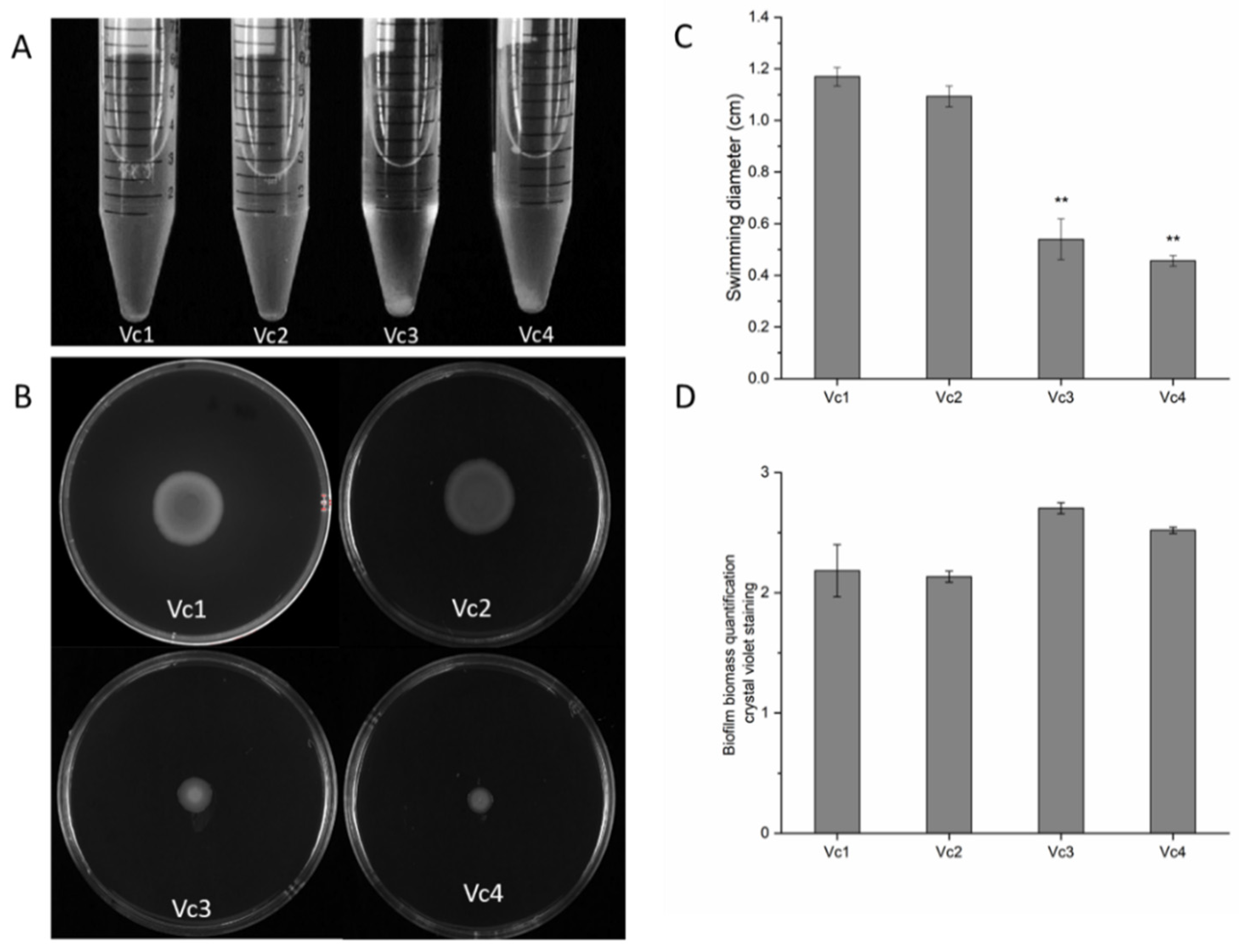
2.8. Motility Assay and Crystal Violet Biofilm Formation Quantification Analysis
2.9. Transmission Electron Microscopy (TEM)
2.10. Statistical Analysis
3. Results
3.1. Prophage Location and Integration Site in the V. cholerae Vc1 Genome
3.2. Development of V. cholerae Vc1 as a Platform for Studying Prophage 919TP
3.3. The Function of Wild-Type cI-Like Repressor in Strain Vc1
3.4. Comparative Genomics and Functional Analysis of Phage φ919TP cI- Resistant Mutant
3.5. Mutation in O-antigen Biosynthesis Gene gmd Suppresses the Phage Infection
3.6. Prophage 919TP Provides Immunity to Phage KVP40
3.7. The Absence of Gene gmd Reduces Motility and Affects Biofilm Formation
4. Discussion
4.1. Development of V. cholerae Vc1 as a Platform for Studying the Roles of Prophage 919TP
4.2. Implications of Phage Resistance for Bacterial Physiological Properties
4.3. Effects of O-antigen Mutation on Biofilm Formation in V. cholerae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faruque, S.M.; Albert, M.J.; Mekalanos, J.J. Epidemiology, genetics, and ecology of toxigenic Vibrio cholerae. Microbiol. Mol. Biol. Rev. 1998, 62, 1301–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudry, B.; Fasano, A.; Ketley, J.; Kaper, J.J.I. Cloning of a gene (zot) encoding a new toxin produced by Vibrio cholerae. Immunity 1992, 60, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Zhang, J.; Xu, J.; Du, P.; Pang, B.; Li, J.; Kan, B. The resistance of Vibrio cholerae O1 El Tor strains to the typing phage 919TP, a member of K139 phage family. Front. Microbiol. 2016, 7, 726. [Google Scholar] [CrossRef] [Green Version]
- Oppenheim, A.B.; Kobiler, O.; Stavans, J.; Court, D.L.; Adhya, S. Switches in bacteriophage lambda development. Annu. Rev. Genet. 2005, 39, 409–429. [Google Scholar] [CrossRef]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef] [Green Version]
- Nesper, J.; Lauriano, C.M.; Klose, K.E.; Kapfhammer, D.; Kraiß, A.; Reidl, J.J. Characterization of vibrio cholerae O1 El TorgalU and galE mutants: Influence on lipopolysaccharide structure, colonization, and biofilm formation. Immunity 2001, 69, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Szymczak, P.; Rau, M.H.; Monteiro, J.M.; Pinho, M.G.; Filipe, S.R.; Vogensen, F.K.; Zeidan, A.A.; Janzen, T. A comparative genomics approach for identifying host-range determinants in Streptococcus thermophilus bacteriophages. Sci. Rep. 2019, 9, 7991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milton, D.L.; O’Toole, R.; Horstedt, P.; Wolf-Watz, H. Flagellin A is essential for the virulence of Vibrio anguillarum. J. Bacteriol. 1996, 178, 1310–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.; Zhang, Y.; Qin, J.; Le, S.; Gu, J.; Chen, L.-k.; Guo, X.; Zhu, T. A frameshift mutation in wcaJ associated with phage resistance in Klebsiella pneumoniae. Microorganisms 2020, 8, 378. [Google Scholar] [CrossRef] [Green Version]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar]
- Tan, D.; Svenningsen, S.L.; Middelboe, M. Quorum sensing determines the choice of antiphage defense strategy in Vibrio anguillarum. mBio 2015, 6, e00627. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.; Hansen, M.F.; de Carvalho, L.N.; Røder, H.L.; Burmølle, M.; Middelboe, M.; Svenningsen, S.L. High cell densities favor lysogeny: Induction of an H20 prophage is repressed by quorum sensing and enhances biofilm formation in Vibrio anguillarum. ISME J. 2020, 14, 1731–1742. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Dahl, A.; Middelboe, M.J.A. Vibriophages differentially influence biofilm formation by Vibrio anguillarum strains. Microbiology 2015, 81, 4489–4497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamorano-Sánchez, D.; Xian, W.; Lee, C.K.; Salinas, M.; Thongsomboon, W.; Cegelski, L.; Wong, G.C.; Yildiz, F. Functional specialization in Vibrio cholerae diguanylate cyclases: Distinct modes of motility suppression and c-di-GMP production. mBio 2019, 10, e01960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, K.A.; Freedman, O.; Ritchings, B.W.; DuBow, M.S. Characterization of the lysogenic repressor (c) gene of the Pseudomonas aeruginosa transposable bacteriophage D3112. Virology 2000, 272, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesper, J.; Blaß, J.; Fountoulakis, M.; Reidl, J. Characterization of the major control region of vibrio cholerae bacteriophage K139: Immunity, exclusion, and integration. J. Bacteriol. 1999, 181, 2902–2913. [Google Scholar] [CrossRef] [Green Version]
- Sturla, L.; Bisso, A.; Zanardi, D.; Benatti, U.; De Flora, A.; Tonetti, M. Expression, purification and characterization of GDP-D-mannose 4, 6-dehydratase from Escherichia coli. FEBS Lett. 1997, 412, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Nesper, J.; Kapfhammer, D.; Klose, K.E.; Merkert, H.; Reidl, J. Characterization of vibrio cholerae O1 antigen as the bacteriophage K139 receptor and identification of IS1004Insertions aborting O1 antigen biosynthesis. J. Bacteriol. 2000, 182, 5097–5104. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, J.; Lu, X.; Liang, W.; Zhang, L.; Kan, B. O antigen is the receptor of Vibrio cholerae serogroup O1 El Tor typing phage VP4. J. Bacteriol. 2013, 195, 798–806. [Google Scholar] [CrossRef] [Green Version]
- Bondy-Denomy, J.; Qian, J.; Westra, E.R.; Buckling, A.; Guttman, D.S.; Davidson, A.R.; Maxwell, K.L. Prophages mediate defense against phage infection through diverse mechanisms. ISME J. 2016, 10, 2854–2866. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Matsuzaki, S.; Tanaka, S. A 26-kDa outer membrane protein, OmpK, common to Vibrio species is the receptor for a broad-host-range vibriophage, KVP40. FEMS Microbiol. Lett. 1995, 125, 101–105. [Google Scholar] [CrossRef]
- Heilmann, S.; Sneppen, K.; Krishna, S. Coexistence of phage and bacteria on the boundary of self-organized refuges. Proc. Natl. Acad. Sci. USA 2012, 109, 12828–12833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140. [Google Scholar] [CrossRef] [PubMed]
- Charles, R.C.; Kelly, M.; Tam, J.M.; Akter, A.; Hossain, M.; Islam, K.; Biswas, R.; Kamruzzaman, M.; Chowdhury, F.; Khan, A.I. Humans surviving cholera develop antibodies against Vibrio cholerae O-specific polysaccharide that inhibit pathogen motility. mBio 2020, 11, e02847. [Google Scholar] [CrossRef]
- Glauert, A.M.; Kerridge, D.; Horne, R.W. The fine structure and mode of attachment of the sheathed flagellum of Vibrio metchnikovii. J. Cell Biol. 1963, 18, 327–336. [Google Scholar] [CrossRef]
- Waldor, M.K.; Mekalanos, J.J. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 1996, 272, 1910–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casjens, S. Prophages and bacterial genomics: What have we learned so far? Mol. Microbiol. 2003, 49, 277–300. [Google Scholar] [CrossRef]
- Singh, K.; Milstein, J.N.; Navarre, W.W. Xenogeneic silencing and its impact on bacterial genomes. Annu. Rev. Microbiol. 2016, 70, 199–213. [Google Scholar] [CrossRef]
- Pfeifer, E.; Michniewski, S.; Gätgens, C.; Münch, E.; Müller, F.; Polen, T.; Millard, A.; Blombach, B.; Frunzke, J. Generation of a prophage-free variant of the fast-growing bacterium Vibrio natriegens. Microbiology 2019, 85, e00853-19. [Google Scholar] [CrossRef] [Green Version]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Steinbacher, S.; Baxa, U.; Miller, S.; Weintraub, A.; Seckler, R.; Huber, R. Crystal structure of phage P22 tailspike protein complexed with Salmonella sp. O-antigen receptors. Proc. Natl. Acad. Sci. USA 1996, 93, 10584–10588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seed, K.D.; Faruque, S.M.; Mekalanos, J.J.; Calderwood, S.B.; Qadri, F.; Camilli, A. Phase variable O antigen biosynthetic genes control expression of the major protective antigen and bacteriophage receptor in Vibrio cholerae O1. PLoS Pathog. 2012, 8, e1002917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, K.; Braun, V. Polymannose O-antigens of Escherichia coli, the binding sites for the reversible adsorption of bacteriophage T5+ via the L-shaped tail fibers. J. Virol. 1982, 41, 222–227. [Google Scholar] [CrossRef] [Green Version]
- Mangalea, M.R.; Duerkop, B. Fitness trade-offs resulting from bacteriophage resistance potentiate synergistic antibacterial strategies. Immunity 2020, 88, e009226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Post, D.M.; Yu, L.; Krasity, B.C.; Choudhury, B.; Mandel, M.J.; Brennan, C.A.; Ruby, E.G.; McFall-Ngai, M.J.; Gibson, B.W.; Apicella, M.A. O-antigen and core carbohydrate of Vibrio fischeri lipopolysaccharide: Composition and analysis of their role in Euprymna scolopes light organ colonization. J. Biol. Chem. 2012, 287, 8515–8530. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, R.C.; Adekunle, O.; Yu, H.; Cho, A.; Nyhoff, L.E.; Kelly, M.; Harris, J.B.; Bhuiyan, T.R.; Qadri, F.; Calderwood, S.B.; et al. Impact of immunoglobulin isotype and epitope on the functional properties of Vibrio cholerae O-specific polysaccharide-specific monoclonal antibodies. mBio 2021, 12, e03679. [Google Scholar] [CrossRef]
- Islam, M.S.; Drasar, B.S.; Sack, R.B. The aquatic environment as a reservoir of Vibrio cholerae: A review. J. Diarrhoeal. Dis. Res. 1993, 197–206. [Google Scholar]
- Waldor, M.K.; Colwell, R.; Mekalanos, J.J. The Vibrio cholerae O139 serogroup antigen includes an O-antigen capsule and lipopolysaccharide virulence determinants. Proc. Natl. Acad. Sci. USA 1994, 91, 11388–11392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spöring, I.; Felgner, S.; Preuße, M.; Eckweiler, D.; Rohde, M.; Häussler, S.; Weiss, S.; Erhardt, M.J.M. Regulation of flagellum biosynthesis in response to cell envelope stress in Salmonella enterica serovar Typhimurium. mBio 2018, 9, e00736. [Google Scholar] [CrossRef] [Green Version]
- Burrows, L.L. Pseudomonas aeruginosa twitching motility: Type IV pili in action. Annu Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, H.; Ma, X.; Lu, A.; Lux, R.; Zusman, D.; Shi, W. Extracellular polysaccharides mediate pilus retraction during social motility of Myxococcus xanthus. Proc. Natl. Acad. Sci. USA 2003, 100, 5443–5448. [Google Scholar] [CrossRef] [Green Version]
- Marinelli, L.J.; Piuri, M.; Swigoňová, Z.; Balachandran, A.; Oldfield, L.M.; van Kessel, J.C.; Hatfull, G.F. BRED: A simple and powerful tool for constructing mutant and recombinant bacteriophage genomes. PLoS ONE 2008, 3, e3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Hathroubi, S.; Hancock, M.; Bosse, J.; Langford, P.; Tremblay, Y.; Labrie, J.; Jacques, M.J. Surface polysaccharide mutants reveal that absence of O antigen reduces biofilm formation of Actinobacillus pleuropneumoniae. Immunity 2016, 84, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Nanda, A.M.; Thormann, K.; Frunzke, J.J. Impact of spontaneous prophage induction on the fitness of bacterial populations and host-microbe interactions. J. Bacteriol. 2015, 197, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, S.; Tanaka, S.; Koga, T.; Kawata, T.J.M. A broad-host-range vibriophage, KVP40, isolated from sea water. Microbiol. Immunol. 1992, 36, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage | Bacterial Strain (EOP) | |||
|---|---|---|---|---|
| Vc1 | Vc2 | Vc3 | Vc4 | |
| KVP40 | 0.03 ± 0.01 | 1.0 | 155.3 ± 57.8 | 188.2 ± 60.7 |
| φ919TP cI- | 6.5 × 10−5 ± 9.01 × 10−6 | 1.0 | - | - |
| φ919TP | - a | 1.0 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, N.; Zeng, Y.; Hu, B.; Zhu, T.; Svenningsen, S.L.; Middelboe, M.; Tan, D. Interactions between the Prophage 919TP and Its Vibrio cholerae Host: Implications of gmd Mutation for Phage Resistance, Cell Auto-Aggregation, and Motility. Viruses 2021, 13, 2342. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122342
Li N, Zeng Y, Hu B, Zhu T, Svenningsen SL, Middelboe M, Tan D. Interactions between the Prophage 919TP and Its Vibrio cholerae Host: Implications of gmd Mutation for Phage Resistance, Cell Auto-Aggregation, and Motility. Viruses. 2021; 13(12):2342. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122342
Chicago/Turabian StyleLi, Na, Yigang Zeng, Bijie Hu, Tongyu Zhu, Sine Lo Svenningsen, Mathias Middelboe, and Demeng Tan. 2021. "Interactions between the Prophage 919TP and Its Vibrio cholerae Host: Implications of gmd Mutation for Phage Resistance, Cell Auto-Aggregation, and Motility" Viruses 13, no. 12: 2342. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122342