Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients

Department of Medicine, Division of Molecular Oncology, Washington University School of Medicine, Campus Box 8069, 660 S. Euclid Ave., St. Louis, MO 63110, USA
*
Author to whom correspondence should be addressed.
Viruses 2011, 3(6), 886-900; https://0-doi-org.brum.beds.ac.uk/10.3390/v3060886
Submission received: 27 April 2011
/
Revised: 7 June 2011
/
Accepted: 9 June 2011
/
Published: 21 June 2011
(This article belongs to the Special Issue Recent Developments in HTLV Research)
Abstract
:Of the millions of HTLV-1 infected carriers worldwide, 3–5% will develop an aggressive T-cell neoplasm that is highly refractory to conventional therapy. The virus carries the Tax oncogene which constitutively activates the NFκB pathway. This co-option of signaling through NFκB provides for the HTLV-1 infected cell an escape from cell cycle arrest and apoptosis, a steady source of growth factors, and a mechanism by which the virus can activate its own target cell. Therapies that target the NFκB pathway sensitize adult T-cell leukemia/lymphoma (ATLL) cells to apoptosis. A focus on translational interrogation of NFκB inhibitors in animal models and ATLL patients is needed to advance NFκB-targeted ATLL therapies to the bedside.
1. Introduction
Of the 15–20 million HTLV-1 infected carriers worldwide, more than 500,000 will develop an aggressive T-cell neoplasm that is highly refractory to conventional therapy. Despite intensive efforts to improve the overall survival, adult T-cell leukemia/lymphoma (ATLL) remains one of the hematologic malignancies with the poorest prognosis. ATLL is typically preceded by decades of clinical latency during which infected cells accumulate selectable traits leading to malignant transformation. Host pathways commandeered by the virus can be used as therapeutic targets and a constitutively activated NFκB pathway has emerged as an essential hit in the development of ATLL. Research into the mechanisms underlying HTLV-1 modulation of the NFκB pathway has revealed the extent to which HTLV-1 oncogenesis is dependent on the NFκB pathway [1,2]. Mouse models of ATLL have also been created to recapitulate the virus induced, NFκB-dependent leukemias and lymphomas. Translational investigations of NFκB inhibitors in mouse models and ATLL patients are necessary to bring this work from bench to bedside.
2. HTLV-1 Oncogenesis Depends on NFκB
During decades of clinical latency HTLV-1 modulates host signaling pathways to promote proliferation of infected cells. Among the genes carried in the viral genome, the 40 kDa viral transcriptional transactivator (Tax) is sufficient to promote cellular transformation. The mechanism utilized by Tax to promote cellular transformation is multi-faceted, broadly involving activation of proliferation, dysregulation of cell cycle checkpoints, and promotion of genetic instability. While the affects of Tax expression in a cell are diverse, Tax depends on a central signaling pathway for its transforming activity. Mouse models in which Tax is expressed in the lymphocyte compartment have constitutively active NFκB and reproduce many aspects of HTLV-1 pathogenesis (Table 1). Tax expression in the absence of other viral factors is sufficient to activate the NFκB pathway and cause leukemia, lymphoma, solid tumors, splenomegaly, and osteolytic disease. More than 20 years ago Nerenberg identified a role for NFκB in tumor growth in a transgenic mouse model in which Tax, under the regulation of the HTLV-1 LTR, developed neurofibromas [3]. However, LTR-driven Tax expression in mouse models most often resulted in disorders associated with chronic inflammation, another NFκB-mediated process [4,5,6,7]. Other promoters were used in transgenic constructs to restrict Tax expression to the lymphoid compartment and better model ATLL-like malignancies. In one mouse model, the granzyme B promoter was used to drive Tax expression in activated T-cells and NK cells. These mice developed leukemia and lymphoma and tumors associated with high levels of NFκB and NFκB-regulated genes [8]. Tax driven by the Lck promoter also causes an ATLL-like malignancy in transgenic mice in which NFκB is constitutively elevated [9,10]. The necessity of NFκB in Tax-mediated transformation was confirmed using HTLV-1 molecular clones in which a single point mutation in the Tax oncogene that disrupts its ability to activate the NFκB pathway (Figure 1) also eliminates the virus’ ability to transform primary cells [11,12]. These data establish that constitutive activation of the NFκB pathway is essential in the process of Tax-mediated oncogenesis.
3. The NFκB Signaling Pathway
The NFκB family of transcription activators are involved in many biological processes including cell survival, stress responses, and development [13]. NFκB is also a central regulator of immune effectors including cytokine and chemokine secretion, receptor expression, antigen presentation, cellular proliferation, and programmed cell death. The NFκB family contains five members (RelA, RelB, cRel, p50, and p52) that can form 15 different homo or heterodimers with a variety of activities and tissue specificities. Latent or unstimulated cells retain NFκB proteins in the cytoplasm bound to various inhibitors of kappa B (IκB) proteins. Upon activation, IκB kinases (IKKs) release NFκB complexes which then translocate to the nucleus through two distinct pathways, termed canonical and non-canonical. These arms are distinct but retain extensive mechanisms of cross-regulation [13]. The canonical pathway, which is activated by pro-inflammatory signals, is IKKγ dependent, while the non-canonical pathway functions independent of IKKγ, instead depending on NFκB inducing kinase (NIK). The non-canonical pathway is activated in response to tumor necrosis factor (TNF) receptors during lymphoid development. It is because NFκB proteins regulate these myriad pathways that cancers and viral pathogens exploit NFκB to enhance proliferation, cell survival, and evasion of immune responses (Figure 2). Consequently, this array of human cancers could also all be targeted by NFκB directed therapies.
4. Tax Targets the NFκB Pathway
Lymphoma is commonly associated with constitutive NFκB activity and oncogenic human viruses, including Epstein-Barr virus (EBV), Human papillomavirus (HPV), Hepatitis B virus (HBV), and Hepatitis C virus (HCV) all carry viral factors that directly target and co-opt the NFκB pathway during oncogenesis. HTLV-1 uses Tax in multiple strategies to efficiently activate both the canonical and non-canonical NFκB pathways (Figure 2).
Tax directly interacts with several NFκB members, including RelA, p50, p52, IKKγ, and the NFκB precursor protein p100 such that both arms of the NFκB pathway are involved in tumorigenesis [1,14]. Rapid and reversible, the canonical pathway of NFκB is transient and strictly regulated. Tax activates the canonical pathway through a variety of mechanisms including binding to IKKγ, activating the Akt pathway which signals through IKKα, or by promoting phosphorylaton of p65 via activation of pro-inflammatory cytokines [15,16,17,18]. HTLV-1 uses the canonical pathway, through Bcl-xl, to prevent intrinsic apoptosis [20]. The non-canonical arm of NFκB, which involves new protein synthesis of p100 and RelB downstream of NIK, is slower and irreversible. Tax-induced p100 processing or its physical interaction with Tax in the nucleus contribute to Tax activation of the non-canonical NFκB pathway as well. The non-canonical pathway has a broader anti-apoptotic effect on both apoptotic pathways through BCL-XL, FLIP, XIAP, and CIAP, making it a more powerful therapeutic target [20]. In fact, the difference in transforming potential between HTLV-1 and HTLV-2 has been linked to the ability of HTLV-1 Tax to activate the non-canonical arm of the NFκB pathway [21].
5. NFκB and Apoptosis
Constitutive activation of NFκB family members and downstream effectors provides several selective advantages to malignant cells. One selective advantage offered by unregulated NFκB signaling is the ability to bypass checkpoint controls and p53-mediated apoptosis. In transformed cell lines as well as freshly isolated ATLL samples, NFκB activation supports the survival and proliferation of HTLV-1 infected cells. Moreover, lymphoma cell lines with constitutively activated NFκB are resistant to a variety of inducers of apoptosis including gamma-irradiation, etoposide, and combinations of cycloheximide and TNF or TRAIL, and resist the activation of both the intrinsic and extrinsic apoptotic pathways [20]. Like ATLL cells, malignant cells arising in mouse models are also resistant to chemotherapy and radiation-induced apoptosis [22]. Although mutations deleting or inactivating p53 are common in ATLL, Tax is capable of bypassing p53-dependent cell-cycle checkpoints through constitutive activation of the NFκB pathway [22]. Tax is known to suppress a wide range of pro-apoptotic factors and stimulate expression of factors acting as apoptosis inhibitors [14]. The necessity of NFκB in Tax-mediated resistance to apoptosis is revealed when overexpression of IκB or loss of p65 represses the NFκB pathway, and re-sensitizes Tax-transformed cells to inducers of p53-mediated apoptosis [23].
6. NFκB and Inflammation
HTLV-1 in humans is also associated with chronic inflammation which is mediated by NFκB. Chronic inflammation resulting from Tax-mediated canonical NFκB activity leads to HAM/TSP in some HTLV-1 carriers [24], but may also contribute to development of ATLL. Chronic inflammation promotes cancer through complex mechanisms involving cytokine mediated proliferation, stromal activation, immune modulation, and release of DNA damage promoting agents [25,26]. Genes activated by Tax in transgenic mice are directly or indirectly regulated by NFκB inducible cytokines that promote inflammation and immune cell infiltration (Figure 3). Tax tumor cells express IL-6, M-CSF, IL-1, TNF-α, and Tax expression enhances IL-6 and TNF-α expression in vitro and in vivo [27,28]. Moreover, malignant cells express NFκB inducible cytokines and stimulate cytokine production in tumor infiltrating cells and stroma. Factors produced by Tax-tumor cells cause splenomegaly, neutrophilia, elevated white count and anemia in transgenic mice and SCID recipients of tumor allografts [27,29]. Preceding tumorigenesis in transgenic mice, Tax and the NFκB pathway promote a state of chronic inflammation in which Tax-induced malignancies can thrive [30].
This inflammation-associated malignancy is Tax-dependent and promoter-dependent since SV40 large T under the same promoter does not reproduce the inflammation promoting nature of Tax tumors [29,31]. Interestingly, chronic inflammation is a complicating factor because it carries both tumor-promoting and tumor-repressing effects. IFNγ, an NFκB inducible gene found in chronic inflammation, causes inhibition of tumor angiogenesis and represses tumor growth [32]. The mechanism by which Tax promotes inflammation and tumorigenesis is coupled by its regulation of the NFκB pathway.
7. NFκB and T-Cells
HTLV-1 infects and transforms CD4+ T-cells. The NFκB pathway is essential for T-cell functions including T-cell development, activation, gene expression, cell cycle progression, survival, cytokine production, and apoptosis [13]. While Tax activates NFκB in T-cells, most freshly isolated ATLL cells do not express detectable levels of Tax [33]. This has been explained by the discovery that Tax is a primary target of cytotoxic T lymphocyte (CTL) attack, and HTLV-1 infected T-cells that express high levels of Tax are destroyed [34]. Is ATLL Tax-independent at the time of clinical presentation? A mouse model in which Tax drives expression of firefly-luciferase, allowed non-invasive, real-time detection of Tax activity using bioluminescence imaging (Figure 3). This model revealed that Tax activity, which was normally undetectable, was occasionally punctuated by short bursts of intense expression. Moreover, these stochastic bursts of Tax expression preceded advancing stages of tumorigenesis [31]. In addition, a triple-transgenic mouse strain carrying an ovalbumin-inducible T-cell receptor (TCR) transgene demonstrated that systemic T-cell activation accelerated the development of Tax-induced lymphoma [35]. Determining whether ATLL ever attains Tax independence awaits an inducible-Tax mouse in which Tax expression can be repressed late in malignancy. Importantly, in ATLL cells the NFκB pathway remains activated when Tax expression is repressed. Thus NFκB remains a therapeutic target even when Tax is not expressed.
8. Targeting NFκB in vivo
NFκB regulates the expression of a wide variety of genes implicated in proliferation, angiogenesis, invasion, and metastasis and the dependence of HTLV-1 oncogenesis on the NFκB pathway makes it an ideal target for therapeutic attack. Repression of the NFκB pathway could make ATLL cells sensitive to apoptosis, slow their proliferation, or repress aspects of the immune response that promote malignancy. In tissue culture and mouse models, non-specific inhibitors of the NFκB pathway like sodium salicylate or cyclopentenone prostaglandins can increase the sensitivity of Tax-tumor cells to apoptosis and repress NFκB-inducible cytokines IL-6, IL-10, IL-15, and IFN-γ [28]. Bortezomib is another non-specific inhibitor of the NFκB pathway that is capable of inhibiting proliferation of Tax tumors cells ex vivo and sensitizing cells to apoptosis [36]. Bortezomib treatment slowed tumor growth in an allograft model by increasing apoptosis, but toxicity constraints limited the efficacy of the treatment [36]. Bay11-7082, an IKK inhibitor, inhibits the NFκB pathway in ATLL cells and sensitizes HTLV-1 infected cells lines as well as primary ATLL cells to apoptosis [37]. Over the past six years several additional studies have therapeutically targeted the NFκB pathway in order to kill ATLL cells [38]. Oridonin, NIK-333, curcumin, fucoidan, histone-deacetylase inhibitors, and a derivative of epoxyquinomicin C have all been reported to induce apoptosis in ATLL cells by repressing the NFκB pathway [39,40,41,42,43,44]. These findings serve as sufficient proof of principle that NFκB-targeted therapies show great promise against ATLL. The field now awaits successful clinical trials in vivo.
9. Targeting NFκB in ATLL Patients
The majority of ATLL patients present with acute or lymphomatous ATLL, which results in a median survival of 0.5–2.0 years, despite intensive chemotherapy treatment [45,46]. To determine if NFκB blockade is tolerated in these patients, and whether or not it improves response rates and overall survival, our current multicenter trial combines infusional chemotherapy (EPOCH) with bortezomib (Figure 4). In addition, this clinical trial includes treatment with integrase inhibitor raltegravir, which was found to inhibit HTLV-1 integration in tissue culture [47]. The addition of an antiviral agent to this ATLL treatment regimen is based on our previous clinical trial in which chemotherapy was found to markedly enhance virus expression in a subset of patients [48].
10. Conclusions
Host pathways can be used as chemotherapeutic targets when they confer an essential trait to the cancer cell. A constitutively activated NFκB pathway represents such a target in the case of HTLV-1 mediated ATLL. The NFκB pathway provides an escape from cell cycle arrest and apoptosis, a steady source of growth factors, and a mechanism by which the virus can activate its own target cell. Accumulating evidence supports the concept that NFκB targeted therapies sensitize ATLL cells to apoptosis. Research in ATLL therapies should now focus on translational interrogation of NFκB inhibitors in animal models and ATLL patients.
Acknowledgments
Funding for D.R. includes grants ACS-IRG 58-010-53 and BJHF 7329-33. Funding for L.R. includes CA 10073, CA94056, CA63417, and LLS 6067-10.
Conflict of Interest
The authors declare no conflict of interest.
References and Notes
- Peloponese, J.; Yeung, M.; Jeang, K. Modulation of nuclear factor-kB by human T-cell leukemia virus type 1 tax protein: Implications for oncogenesis and inflammation. Immunol. Res. 2006, 34, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Horie, R. NF-kB in pathogenesis and treatment of adult T-cell leukemia/lymphoma. Int. Rev. Immunol. 2007, 26, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Nerenberg, M.I. An HTLV-I Transgenic Mouse Model: Role of the tax gene in pathogenesis in multiple organ systems. Curr. Top. Microbiol. Immunol. 1990, 160, 121–128. [Google Scholar] [PubMed]
- Iwakura, Y.; Tosu, M.; Yoshida, E.; Takiguchi, M.; Sato, K.; Kitajima, I.; Nishioka, K.; Yamamoto, K.; Takeda, T.; Hatanaka, M.; et al. Induction of inflammatory arthropathy resembling rheumatoid arthritis in mice transgenic for HTLV-I. Science 1991, 253, 1026–1028. [Google Scholar] [CrossRef] [PubMed]
- Iwakura, Y.; Saijo, S.; Kioka, Y.; Nakayama-Yamada, J.; Itagaki, K.; Tosu, M.; Asano, M.; Kanai, Y.; Kakimoto, K. Autoimmunity induction by human T-cell leukemia virus type 1 in transgenic mice that develop chronic inflammatory arthropathy resembling rheumatoid arthritis in humans. J. Immunol. 1995, 155, 1588–1598. [Google Scholar] [CrossRef]
- Yamamoto, H.; Sekiguchi, T.; Itagaki, K.; Saijo, S.; Iwakura, Y. Inflammatory polyarthritis in mice transgenic for human T-cell leukemia virus type I. Arthritis Rheum. 1993, 36, 1612–1620. [Google Scholar] [CrossRef]
- Fujisawa, K.; Asahara, H.; Okamoto, K.; Aono, H.; Hasunuma, T.; Kobata, T.; Iwakura, Y.; Yonehara, S.; Sumida, T.; Nishioka, K. Therapeutic effect of the anti-Fas antibody on arthritis in HTLV-1 tax transgenic mice. J. Clin. Invest. 1996, 98, 271–278. [Google Scholar] [CrossRef]
- Grossman, W.; Kimata, J.; Wong, F.; Zutter, M.; Ley, T.; Ratner, L. Development of leukemia in mice transgenic for the tax gene of human T-cell leukemia virus type I. Proc. Natl. Acad. Sci. U. S. A. 1995, 92, 1057–1061. [Google Scholar] [CrossRef]
- Hasegawa, H.; Sawa, H.; Lewis, M.; Orba, Y.; Sheehy, N.; Yamamoto, Y.; Ichinohe, T.; Tsunetsugu-Yokota, Y.; Katano, H.; Takahashi, H.; et al. Thymus-derived leukemia-lymphoma in mice transgenic for the Tax gene of human T-lymphotropic virus type I. Nat. Med. 2006, 12, 466–472. [Google Scholar] [CrossRef]
- Watters, K.; Dean, J.; Hasegawa, H.; Sawa, H.; Hall, W.; Sheehy, N. Cytokine and growth factor expression by HTLV-1 Lck-tax transgenic cells in SCID mice. AIDS Res. Hum. Retroviruses 2010, 26, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Kimata, J.; Wong, F.; Wang, J.; Ratner, L. Construction and characterization of infectious human T-cell leukemia virus type 1 molecular clones. Virology 1994, 204, 656–664. [Google Scholar] [CrossRef]
- Robek, M.; Ratner, L. Immortalization of CD4(+) and CD8(+) T lymphocytes by human T-cell leukemia virus type 1 Tax mutants expressed in a functional molecular clone. J. Virol. 1999, 73, 4856–4865. [Google Scholar] [CrossRef] [PubMed]
- Shih, V.; Tsui, R.; Caldwell, A.; Hoffmann, A. A single NFκB system for both canonical and non-canonical signaling. Cell Res. 2011, 21, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Saggioro, D.; Silic-Benussi, M.; Biasiotto, R.; D'Agostino, D.; Ciminale, V. Control of cell death pathways by HTLV-1 proteins. Front. Biosci. 2009, 14, 3338–3351. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Ballard, D. Persistent activation of NFκB by the tax transforming protein of HTLV-1; hijacking cellular IkappaB kinases. Oncogene 1999, 18, 6948–6958. [Google Scholar] [CrossRef]
- Jeong, S.; Pise-Masison, C.; Radonovich, M.; Park, H.; Brady, J. Activated AKT regulates NF-kappaB activation, p53 inhibition and cell survival in HTLV-1-transformed cells. Oncogene 2005, 24, 6719–6728. [Google Scholar] [CrossRef]
- Tanaka, H.; Fujita, N.; Tsuruo, T. 3-Phosphoinositide-dependent protein kinase-1-mediated IkappaB kinase beta (IkkB) phosphorylation activates NF-kappaB signaling. J. Biol. Chem. 2005, 280, 40965–40973. [Google Scholar] [CrossRef]
- O'Mahony, A.; Montano, M.; Van Beneden, K.; Chen, L.; Greene, W. Human T-cell lymphotropic virus type 1 tax induction of biologically active NF-kappaB requires IkappaB kinase-1-mediated phosphorylation of RelA/p65. J. Biol. Chem. 2004, 279, 18137–18145. [Google Scholar] [CrossRef]
- Xiao, G.; Cvijic, M.; Fong, A.; Harhaj, E.; Uhlik, M.; Waterfield, M.; Sun, S. Retroviral oncoprotein Tax induces processing of NF-kappaB2/p100 in T cells: Evidence for the involvement of IKKalpha. EMBO J. 2001, 20, 6805–6815. [Google Scholar] [CrossRef]
- Bernal-Mizrachi, L.; Lovly, C.; Ratner, L. The role of NF-{kappa}B-1 and NF-{kappa}B-2-mediated resistance to apoptosis in lymphomas. Proc. Natl. Acad. Sci. U S A. 2006, 103, 9220–9225. [Google Scholar] [CrossRef]
- Shoji, T.; Higuchi, M.; Kondo, R.; Takahashi, M.; Oie, M.; Tanaka, Y.; Aoyagi, Y.; Fujii, M. Identification of a novel motif responsible for the distinctive transforming activity of human T-cell leukemia virus (HTLV) type 1 Tax1 protein from HTLV-2 Tax2. Retrovirology 2009, 6, 83. [Google Scholar] [CrossRef] [PubMed]
- Portis, T.; Grossman, W.; Harding, J.; Hess, J.; Ratner, L. Analysis of p53 inactivation in a human T-cell leukemia virus type 1 Tax transgenic mouse model. J. Virol. 2001, 75, 2185–2193. [Google Scholar] [CrossRef] [PubMed]
- Pise-Masison, C.; Mahieux, R.; Jiang, H.; Ashcroft, M.; Radonovich, M.; Duvall, J.; Guillerm, C.; Brady, J. Inactivation of p53 by human T-cell lymphotropic virus type 1 Tax requires activation of the NF-kappaB pathway and is dependent on p53 phosphorylation. Mol. Cell. Biol. 2000, 10, 3377–3386. [Google Scholar] [CrossRef] [PubMed]
- Oh, U.; McCormick, M.; Datta, D.; Turner, R.; Bobb, K.; Monie, D.; Sliskovic, D.; Tanaka, Y.; Zhang, J.; Meshulam, J.; et al. Inhibition of immune activation by a novel nuclear factor-kappa B inhibitor in HTLV-I-associated neurologic disease. Blood 2011, 117, 3363–3369. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Schäfer, M.; Werner, S. Cancer as an overhealing wound: An old hypothesis revisited. Nat. Rev. Mol. Cell. Biol. 2008, 9, 628–638. [Google Scholar] [CrossRef]
- Gao, L.; Deng, H.; Zhao, H.; Hirbe, A.; Harding, J.; Ratner, L.; Weilbaecher, K. HTLV-1 Tax transgenic mice develop spontaneous osteolytic bone metastases prevented by osteoclast inhibition. Blood 2005, 106, 4294–4302. [Google Scholar] [CrossRef]
- Portis, T.; Harding, J.; Ratner, L. The contribution of NF-kappa B activity to spontaneous proliferation and resistance to apoptosis in human T-cell leukemia virus type 1 Tax-induced tumors. Blood 2001, 98, 1200–1208. [Google Scholar] [CrossRef]
- Grossman, W.; Ratner, L. Cytokine expression and tumorigenicity of large granular lymphocytic leukemia cells from mice transgenic for the tax gene of human T-cell leukemia virus type I. Blood 1997, 90, 783–794. [Google Scholar] [CrossRef]
- Gross, S.; Gammon, S.; Moss, B.; Rauch, D.; Harding, J.; Heinecke, J.; Ratner, L.; Piwnica-Worms, D. Bioluminescence imaging of myeloperoxidase activity in vivo. Nat. Med. 2009, 15, 455–461. [Google Scholar] [CrossRef]
- Rauch, D.; Gross, S.; Harding, J.; Niewiesk, S.; Lairmore, M.; Piwnica-Worms, D.; Ratner, L. Imaging spontaneous tumorigenesis: inflammation precedes development of peripheral NK tumors. Blood 2009, 113, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Mitra-Kaushik, S.; Harding, J.; Hess, J.; Schreiber, R.; Ratner, L. Enhanced tumorigenesis in HTLV-1 tax-transgenic mice deficient in interferon-gamma. Blood 2004, 104, 3305–3311. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, Y.; Osame, M.; Kubota, R.; Tara, M.; Yoshida, M. Human T-cell leukemia virus type-1 (HTLV-1) Tax is expressed at the same level in infected cells of HTLV-1-associated myelopathy or tropical spastic paraparesis patients as in asymptomatic carriers but at a lower level in adult T-cell leukemia cells. Blood 1995, 85, 1865–1870. [Google Scholar] [CrossRef] [PubMed]
- Bangham, C.; Osame, M. Cellular immune response to HTLV-1. Oncogene 2005, 24, 6035–6046. [Google Scholar] [CrossRef] [PubMed]
- Rauch, D.; Gross, S.; Harding, J.; Bokhari, S.; Niewiesk, S.; Lairmore, M.; Piwnica-Worms, D.; Ratner, L. T-cell activation promotes tumorigenesis in inflammation-associated cancer. Retrovirology 2009, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- Mitra-Kaushik, S.; Harding, J.; Hess, J.; Ratner, L. Effects of the proteasome inhibitor PS-341 on tumor growth in HTLV-1 Tax transgenic mice and Tax tumor transplants. Blood 2004, 104, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Yamada, Y.; Ikeda, S.; Yamasaki, Y.; Tsukasaki, K.; Tanaka, Y.; Tomonaga, M.; Yamamoto, N.; Fujii, M. Bay 11–7082 inhibits transcription factor NF-kappaB and induces apoptosis of HTLV-I-infected T-cell lines and primary adult T-cell leukemia cells. Blood 2002, 100, 1828–1834. [Google Scholar] [CrossRef]
- Horie, R. NF-kappaB in pathogenesis and treatment of adult T-cell leukemia/lymphoma. Int. Rev. Immunol. 2007, 26, 269–281. [Google Scholar] [CrossRef]
- Ikezoe, T.; Yang, Y.; Bandobashi, K.; Saito, T.; Takemoto, S.; Machida, H.; Togitani, K.; Koeffler, H.; Taguchi, H. Oridonin, a diterpenoid purified from Rabdosia rubescens, inhibits the proliferation of cells from lymphoid malignancies in association with blockade of the NF-kappa B signal pathways. Mol. Cancer Ther. 2005, 4, 578–586. [Google Scholar] [CrossRef]
- Okudaira, T.; Tomita, M.; Uchihara, J.; Matsuda, T.; Ishikawa, C.; Kawakami, H.; Masuda, M.; Tanaka, Y.; Ohshiro, K.; Takasu, N.; et al. NIK-333 inhibits growth of human T-cell leukemia virus type I-infected T-cell lines and adult T-cell leukemia cells in association with blockade of nuclear factor-kappaB signal pathway. Mol. Cancer Ther. 2006, 5, 704–712. [Google Scholar] [CrossRef]
- Tomita, M.; Kawakami, H.; Uchihara, J.; Okudaira, T.; Masuda, M.; Takasu, N.; Matsuda, T.; Ohta, T.; Tanaka, Y.; Ohshiro, K.; et al. Curcumin (diferuloylmethane) inhibits constitutive active NF-kappaB, leading to suppression of cell growth of human T-cell leukemia virus type I-infected T-cell lines and primary adult T-cell leukemia cells. Int. J. Cancer 2006, 118, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Haneji, K.; Matsuda, T.; Tomita, M.; Kawakami, H.; Ohshiro, K.; Uchihara, J.; Masuda, M.; Takasu, N.; Tanaka, Y.; Ohta, T.; et al. Fucoidan extracted from Cladosiphon okamuranus Tokida induces apoptosis of human T-cell leukemia virus type 1-infected T-cell lines and primary adult T-cell leukemia cells. Nutr. Cancer 2005, 52, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, C.; Ikezoe, T.; Yang, J.; Komatsu, N.; Bandobashi, K.; Taniguchi, A.; Kuwayama, Y.; Togitani, K.; Koeffler, H.; Taguchi, H. Histone deacetylase inhibitors induce growth arrest and apoptosis of HTLV-1-infected T-cells via blockade of signaling by nuclear factor kappaB. Leuk. Res. 2008, 32, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Horie, R.; Kumasaka, T.; Ishida, A.; Ishida, T.; Yamaguchi, K.; Watanabe, T.; Umezawa, K.; Urano, T. In vivo antitumor activity of the NF-kappaB inhibitor dehydroxymethylepoxyquinomicin in a mouse model of adult T-cell leukemia. Carcinogenesis 2005, 26, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, M. Diagnostic criteria and classification of clinical subtypes of adult T-cell leukaemia-lymphoma. A report from the Lymphoma Study Group (1984–87). Br. J. Haematol. 1991, 79, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Tsukasaki, K.; Hermine, O.; Bazarbachi, A.; Ratner, L.; Ramos, J.; Harrington, W., Jr.; O'Mahony, D.; Janik, J.; Bittencourt, A.; Taylor, G.; et al. Definition, prognostic factors, treatment, and response criteria of adult T-cell leukemia-lymphoma: A proposal from an international consensus meeting. J. Clin. Oncol. 2009, 27, 453–459. [Google Scholar] [CrossRef]
- Seegulam, M.; Ratner, L. Integrase Inhibitors Effective Against Human T-Cell Leukemia Virus Type 1. Antimicrob. Agents Chemother. 2011, 55, 2011–2017. [Google Scholar] [CrossRef]
- Ratner, L.; Harrington, W.; Feng, X.; Grant, C.; Jacobson, S.; Noy, A.; Sparano, J.; Lee, J.; Ambinder, R.; Campbell, N.; et al. AIDS Malignancy Consortium. Human T-cell leukemia virus reactivation with progression of adult T-cell leukemia-lymphoma. Plos ONE 2009, 4, e4420. [Google Scholar] [CrossRef]
- Nerenberg, M.; Hinrichs, S.; Reynolds, R.; Khoury, G.; Jay, G. The tat Gene of Human T-Lymphotropic Virus Type 1 Induces Mesenchymal Tumors in Transgenic Mice. Science 1987, 237, 1324–1329. [Google Scholar] [CrossRef]
- Kim, S.; Winokur, T.; Lee, H.; Danielpour, D.; Kim, K.; Geiser, A.; Chen, L.; Sporn, M.; Roberts, A.; Jay, G. Overexpression of transforming growth factor-beta in transgenic mice carrying the human T-cell lymphotropic virus type I tax gene. Mol. Cell. Biol. 1991, 11, 5222–5228. [Google Scholar] [CrossRef]
- Kitajima, I.; Shinohara, T.; Bilakovics, J.; Brown, D.; Xu, X.; Nerenberg, M. Ablation of transplanted HTLV-I Tax-transformed tumors in mice by antisense inhibition of NF-kappa B. Science 1992, 258, 1792–1795. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, S.; Nerenberg, M.; Reynolds, R.; Khoury, G.; Jay, G. A transgenic mouse model for human neurofibromatosis. Science 1987, 237, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Baird, A.; Hinrichs, S.; Klintworth, G.; Jay, G. Adrenal medullary tumors and iris proliferation in a transgenic mouse model of neurofibromatosis. Am. J. Pathol. 1992, 140, 1401–1410. [Google Scholar] [PubMed]
- Feigenbaum, L.; Fujita, K.; Collins, F.; Jay, G. Repression of the NF1 gene by Tax may explain the development of neurofibromas in human T-lymphotropic virus type 1 transgenic mice. J. Virol. 1996, 70, 3280–3285. [Google Scholar] [CrossRef]
- Green, J. Trans activation of nerve growth factor in transgenic mice containing the human T-cell lymphotropic virus type I tax gene. Mol. Cell. Biol. 1991, 11, 4635–4641. [Google Scholar]
- Nerenberg, M.; Wiley, C. Degeneration of oxidative muscle fibers in HTLV-1 tax transgenic mice. Am. J. Pathol. 1989, 135, 1025–1033. [Google Scholar]
- Green, J.; Hinrichs, S.; Vogel, J.; Jay, G. Exocrinopathy resembling Sjogren's syndrome in HTLV-1 tax transgenic mice. Nature 1989, 341, 72–74. [Google Scholar] [CrossRef]
- Peebles, R.; Maliszewski, C.; Sato, T.; Hanley-Hyde, J.; Maroulakou, I.; Hunziker, R.; Schneck, J.; Green, J. Abnormal B-cell function in HTLV-I-tax transgenic mice. Oncogene 1995, 10, 1045–1051. [Google Scholar]
- Ruddle, N.; Li, C.; Horne, W.; Santiago, P.; Troiano, N.; Jay, G.; Horowitz, M.; Baron, R. Mice transgenic for HTLV-I LTR-tax exhibit tax expression in bone, skeletal alterations, and high bone turnover. Virology 1993, 197, 196–204. [Google Scholar] [CrossRef]
- Bieberich, C.; King, C.; Tinkle, B.; Jay, G. A transgenic model of transactivation by the Tax protein of HTLV-I. Virology 1993, 196, 309–318. [Google Scholar] [CrossRef]
- Xu, X.; Brown, D.; Kitajima, I.; Bilakovics, J.; Fey, L.; Nerenberg, M. Transcriptional suppression of the human T-cell leukemia virus type I long terminal repeat occurs by an unconventional interaction of a CREB factor with the R region. Mol. Cell. Biol. 1994, 14, 5371–5383. [Google Scholar] [PubMed]
- Furuta, Y.; Aizawa, S.; Suda, Y.; Ikawa, Y.; Kishimoto, H.; Asano, Y.; Tada, T.; Hikikoshi, A.; Yoshida, M.; Seiki, M. Thymic atrophy characteristic in transgenic mice that harbor pX genes of human T-cell leukemia virus type I. J. Virol. 1989, 63, 3185–3189. [Google Scholar] [CrossRef] [PubMed]
- Habu, K.; Nakayama-Yamada, J.; Asano, M.; Saijo, S.; Itagaki, K.; Horai, R.; Yamamoto, H.; Sekiguchi, T.; Nosaka, T.; Hatanaka, M.; et al. The human T cell leukemia virus type I-tax gene is responsible for the development of both inflammatory polyarthropathy resembling rheumatoid arthritis and noninflammatory ankylotic arthropathy in transgenic mice. J. Immunol. 1999, 162, 2956–2963. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Saijo, S.; Arai, M.; Karasawa, S.; Ueda, S.; Kannagi, M.; Iwakura, Y.; Fujii, M.; Yonehara, S. Resistance to fas-mediated apoptosis of peripheral T cells in human T lymphocyte virus type I (HTLV-I) transgenic mice with autoimmune arthropathy. J. Exp. Med. 1997, 186, 57. [Google Scholar] [CrossRef]
- Coscoy, L.; Gonzalez-Dunia, D.; Tangy, F.; Syan, S.; Brahic, M.; Ozden, S. Molecular mechanism of tumorigenesis in mice transgenic for the human T cell leukemia virus Tax gene. Virology 1998, 248, 332–341. [Google Scholar] [CrossRef]
- Benvenisty, N.; Ornitz, D.; Bennett, G.; Sahagan, B.; Kuo, A.; Cardiff, R.; Leder, P. Brain tumours and lymphomas in transgenic mice that carry HTLV-I LTR/c-myc and Ig/tax genes. Oncogene 1992, 7, 2399–2405. [Google Scholar]
- Rauch, D.; Hurchla, M.; Harding, J.; Deng, H.; Shea, L.; Eagleton, M.; Niewiesk, S.; Lairmore, M.; Piwnica-Worms, D.; Rosol, T.; et al. The ARF tumor suppressor regulates bone remodeling and osteosarcoma development in mice. PLoS ONE 2010, 5, e15755. [Google Scholar] [CrossRef]
- Kwon, H.; Ogle, L.; Benitez, B.; Bohuslav, J.; Montano, M.; Felsher, D.; Greene, W. Lethal cutaneous disease in transgenic mice conditionally expressing type I human T cell leukemia virus Tax. J. Biol. Chem. 2005, 280, 35713–35722. [Google Scholar] [CrossRef]
- El Hajj, H.; El-Sabban, M.; Hasegawa, H.; Zaatari, G.; Ablain, J.; Saab, S.; Janin, A.; Mahfouz, R.; Nasr, R.; Kfoury, Y.; et al. Therapy-induced selective loss of leukemia-initiating activity in murine adult T cell leukemia. J. Exp. Med. 2010, 207, 2785–2792. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Orba, Y.; Kimura, T.; Iha, H.; Ogata, M.; Tsuji, T.; Ainai, A.; Sata, T.; Okamoto, T.; Hall, W.; et al. Inhibition of the SDF-1alpha-CXCR4 axis by the CXCR4 antagonist AMD3100 suppresses the migration of cultured cells from ATL patients and murine lymphoblastoid cells from HTLV-I Tax transgenic mice. Blood 2009, 114, 2961–2968. [Google Scholar] [CrossRef]
- Yamazaki, J.; Mizukami, T.; Takizawa, K.; Kuramitsu, M.; Momose, H.; Masumi, A.; Ami, Y.; Hasegawa, H.; Hall, W.; Tsujimoto, H.; et al. Identification of cancer stem cells in a Tax-transgenic (Tax-Tg) mouse model of adult T-cell leukemia/lymphoma. Blood 2009, 114, 2709–2720. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Kumasaka, T.; Okada, S.; Urano, T. The Tax protein of HTLV-1 promotes oncogenesis in not only immature T cells but also mature T cells. Nat. Med. 2007, 13, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Kumasaka, T. Low CD4/CD8 T-cell ratio associated with inflammatory arthropathy in human T-cell leukemia virus type I Tax transgenic mice. PLoS ONE 2011, 6, e18518. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.; Irvine, J.; Blyth, K.; Cameron, E.; Onions, D.; Campbell, M. Tumours derived from HTLV-I tax transgenic mice are characterized by enhanced levels of apoptosis and oncogene expression. J. Pathol. 1998, 186, 209–214. [Google Scholar] [CrossRef]
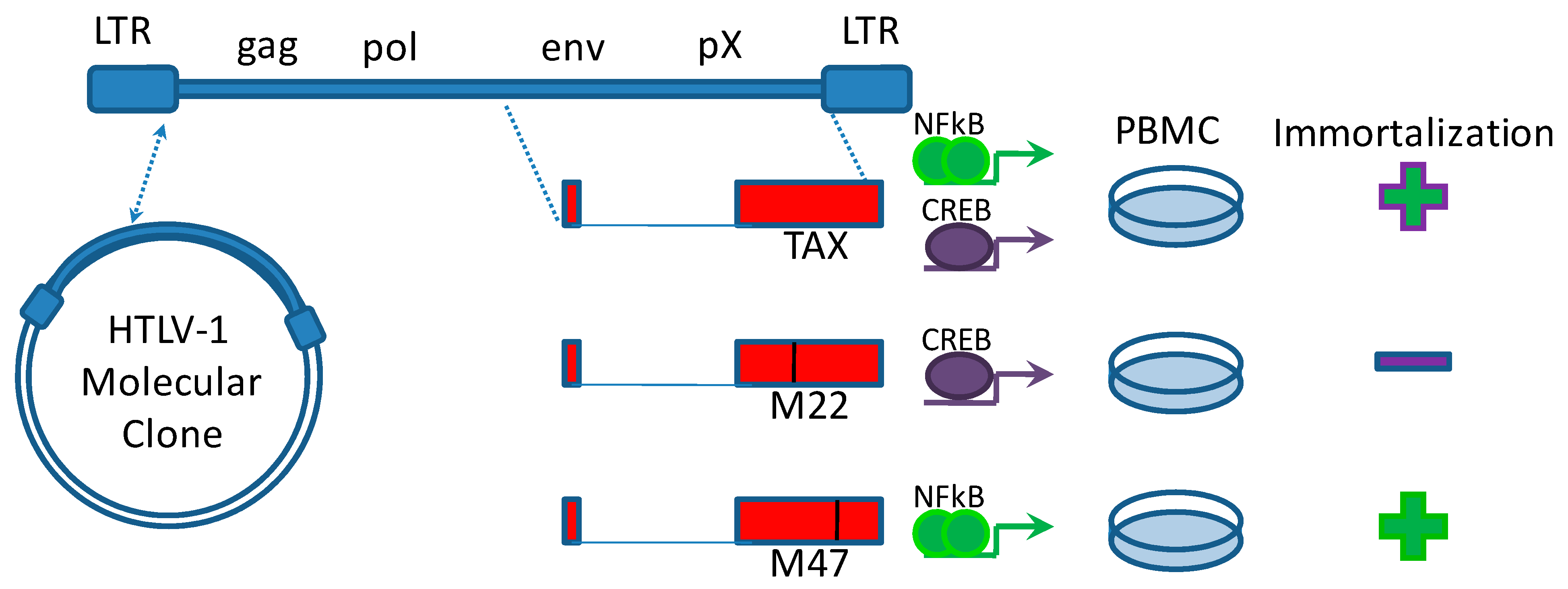
Figure 1.
Tax activation of NFκB is required for immortalization. The creation of a molecular clone of HTLV-1 by Kimata et al. [11] enabled the analysis of Tax mutants for immortalization determinants [12]. Mutations that inhibit Tax activation of the NFκB pathway prevented immortalization of peripheral blood mononuclear cells (PBMC), whereas mutations that inhibit Tax activation of the CREB pathway allowed NFκB activation and Tax-mediated immortalization.
Figure 1.
Tax activation of NFκB is required for immortalization. The creation of a molecular clone of HTLV-1 by Kimata et al. [11] enabled the analysis of Tax mutants for immortalization determinants [12]. Mutations that inhibit Tax activation of the NFκB pathway prevented immortalization of peripheral blood mononuclear cells (PBMC), whereas mutations that inhibit Tax activation of the CREB pathway allowed NFκB activation and Tax-mediated immortalization.

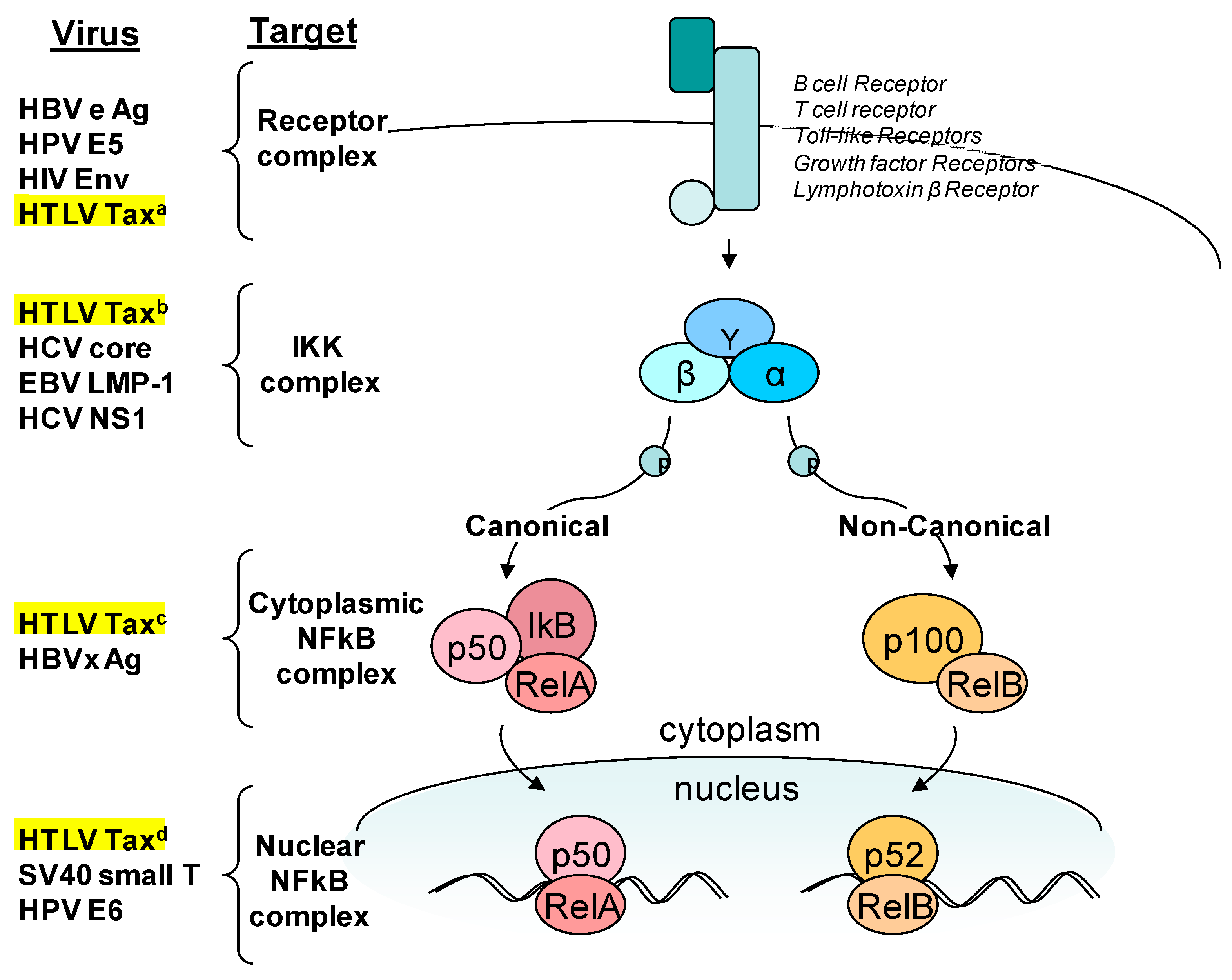
Figure 2.
Tax targets the NFκB pathway. A simplified schematic of NFκB signaling pathways highlighting regulatory proteins commonly targeted by viruses [1,2,13]. HTLV-1 Tax targets the pathway at several points. (a) Tax leads to the activation of receptor-associated kinases that signal through the NFκB pathway [16,17]. (b) Tax directly binds to IKKγ (NEMO) which leads to constitutive phosphorylation and degradation of the NFκB repressor IκB [15]. (c) Tax activates and recruits IKKα to p100 stimulating phosphorylation, ubiquitination, and processing to p52 leading to nuclear translocation [19]. (d) Tax alters binding or recognition of a variety of transcription factors and DNA binding proteins increasing the number of genes regulated by the NFκB pathway [18].
Figure 2.
Tax targets the NFκB pathway. A simplified schematic of NFκB signaling pathways highlighting regulatory proteins commonly targeted by viruses [1,2,13]. HTLV-1 Tax targets the pathway at several points. (a) Tax leads to the activation of receptor-associated kinases that signal through the NFκB pathway [16,17]. (b) Tax directly binds to IKKγ (NEMO) which leads to constitutive phosphorylation and degradation of the NFκB repressor IκB [15]. (c) Tax activates and recruits IKKα to p100 stimulating phosphorylation, ubiquitination, and processing to p52 leading to nuclear translocation [19]. (d) Tax alters binding or recognition of a variety of transcription factors and DNA binding proteins increasing the number of genes regulated by the NFκB pathway [18].

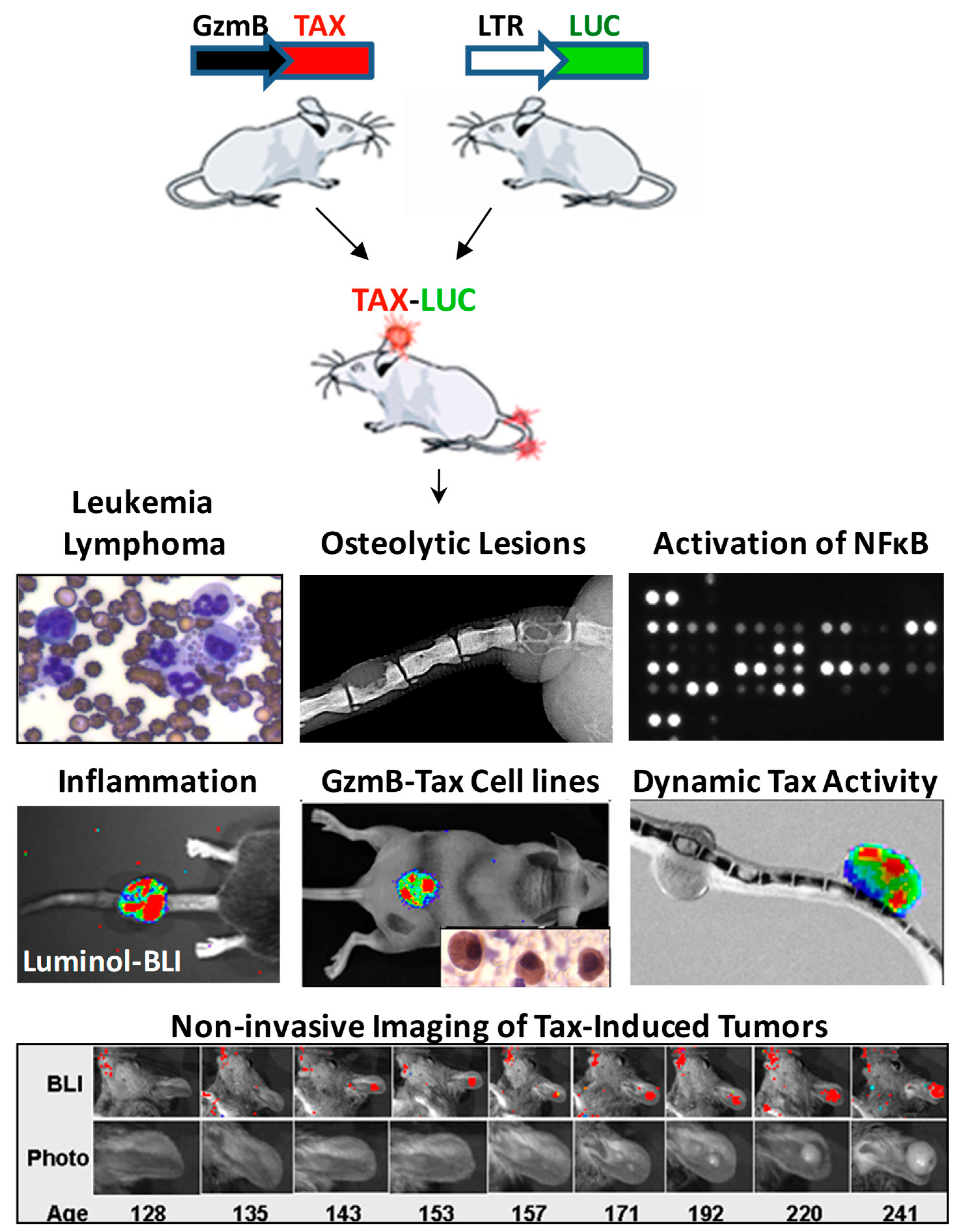
Figure 3.
Tax transgenic mice model ATLL. Several Tax transgenic mouse models of ATLL have demonstrated the sufficiency of Tax as an independent oncogene. Second generation strains, such as the one depicted, have added capabilities, which enable non-invasive interrogation of various Tax activities using bioluminescence imaging [30,31,35].
Figure 3.
Tax transgenic mice model ATLL. Several Tax transgenic mouse models of ATLL have demonstrated the sufficiency of Tax as an independent oncogene. Second generation strains, such as the one depicted, have added capabilities, which enable non-invasive interrogation of various Tax activities using bioluminescence imaging [30,31,35].

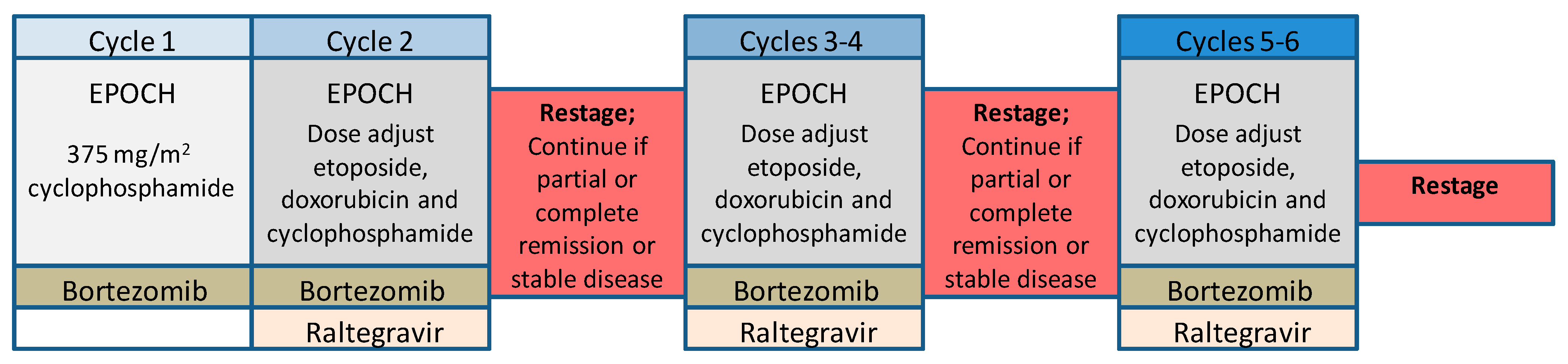
Figure 4.
Schema for an ATLL clinical trial using NFκB targeted therapy. The clinical trial shown has been approved and is currently accepting patients. Clinical trials of NFκB-targeted combination chemotherapies are now beginning to apply the information obtained from primary research to clinical practice.
Figure 4.
Schema for an ATLL clinical trial using NFκB targeted therapy. The clinical trial shown has been approved and is currently accepting patients. Clinical trials of NFκB-targeted combination chemotherapies are now beginning to apply the information obtained from primary research to clinical practice.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of tax transgenic mouse models of adult T-cell leukemia/lymphoma (ATLL).
| Promoter | Gene | Strain | Diseases Observed | Relevance to ATLL | Merits of Model | Weaknesses of Model | Therapies Tested | Ref |
|---|---|---|---|---|---|---|---|---|
| HTLV LTR | Tax | C57BL/6 X DBA/2 X CD1 | Mesenchymal Tumors Thymic atrophy | Not Observed | Overexpression of TGF-beta, activation of NFκB | Tax transforms fibroblasts but not thymocytes | NFκB ODN | [49] [50] [51] |
| Neurofibromatosis Adrenal Tumors | Not Observed | Tax activation of Nerve growth factor and repression of NF1 | Not associated with HTLV associated diseases in humans | None | [52] [53] [54] [55] | |||
| Muscle degeneration | Myositis | High levels of Tax expression in muscle | Incomplete penetrance | None | [56] | |||
| Exocrinopathy Lymphadenopathy Splenomegaly | Sjogren syndrome ocular lesions | NfκB inflammatory disorders associated with Tax | Caused by B not T lymphocytes | None | [57] [58] | |||
| Bone Turnover | Lytic bone lesions | NFkB associated bone lesions | Incomplete penetrance | None | [59] | |||
| Tax βgal | Mesenchymal Tumors | Not Observed | Tax expression in response to tissue damage | Tissue damage not correlated with tumor | None | [60] [61] | ||
| HTLV LTR | pX | C57BL/6 X CD1 | Thymic Atrophy | Not Observed | Effects of pX on thymus independent of promoter used | pX gene expression not detectable | None | [62] |
| Ig-SV40 | ||||||||
| MMTV LTR | ||||||||
| HTLV LTR | pX | C3H/ HeN | Inflammatory Arthropathy Osteogenesis Autoimmunity | Arthritis | IL-1, IL-6, TNFα, TGFβ detected in joints. | No malignancy | Anti-Fas mAb (RK-8) | [4] [5] [7] [63] [64] |
| Tax | ||||||||
| CD4 | Tax | |||||||
| HTLV LTR | Tax | C3H | Mesenchymal Tumors | NFκB mediated malignancy | IκB degradation leads to constitutive NFκB activation | Expression restricted to CNS and testes | None | [65] |
| Ig | Tax | FVB/N | Not Observed | Not Observed | Lymphoma with CNS involvement | Roles of c-Myc and Tax unclear | None | [66] |
| Ig HTLV LTR | Tax c-myc | CD4+ Lymphoma CNS Tumors | CD4+ Lymphoma | |||||
| GzmB | Tax | C57BL/6 | LGL lymphoma Leukemia Osteolytic lesions Splenomegaly Lymphadenopathy Hypercalcemia | Lymphoma Leukemia Lytic bone lesions Hypercalcemia | NFkB mediated leukemia lymphoma | Not a CD4+ T cell malignancy | Bortezomib | [8] [36] |
| Tax IL-2−/− | IL-2 not required for phenotype | Not a CD4+ T cell malignancy | None | [29] | ||||
| Tax IFNγ−/− | Accelerated tumor onset and death | May also affect tumor immunity | None | [32] | ||||
| Tax P53−/− | Accelerated disease progression | Only seen in P53+/− mice | None | [22] | ||||
| GzmB ApoE | Tax OPG | Reduced cancer and bone lesions | Causes osteopetrosis | Zoledronic Acid | [27] | |||
| GzmB HTLV LTR | Tax LUC | C57BL/6 X FVB | Bioluminescent tumors | Not a CD4+ T cell malignancy | None | [31] | ||
| GzmB HTLV LTR | Tax LUC ARF−/− | Lymphoma Bone Turnover Osteosarcoma | ARF−/− is not equivalent to p53−/− | Osteosarcoma not associated with ATLL | Zoledronic Acid | [67] | ||
| GzmB HTLV LTR TCR | Tax LUC TCRova | C57BL/6 X FVB X BALB/c | Leukemia Lymphoma Lymphadenopathy | Tax induced by wounding and T cell activation leads to enhanced tumorigenesis | Primary malignancy not a not a CD4+ T cell malignancy | None | [35] | |
| EμSRα TET TET | tTA Tax M47 | FVB/N | Alopecia Hyperkeratosis Splenomegaly | Skin Lesions | Tet-inducible model allows repression of Tax | No malignancy | None | [68] |
| EμSRα TET | tTA M22 | Not observed | Not Observed | Control establishes role of NFκB in disease | ||||
| Lck-prox | Tax | C57BL/6 | CD4- CD25+ pre-T cell Leukemia Lymphoma | Leukemia Lymphoma | Cancer stem cells derived from these mice recapitulate disease in SCID | Not a CD4+ T cell malignancy | As2O3 + IFN-α AMD3100 | [9] [69] [70] [71] |
| Lck-dis | Tax | C57BL/6 X DBA/2 | CD4+ CD25- mature T cell leukemia lymphoma Arthritis | Leukemia Lymphoma | Mature CD4+ or CD8+ T cell malignancy | Cells lack CD25 | None | [72] [73] |
| CD3-ε | Tax | C57BL/6 X CBA | Mesencymal tumors Mammary Adenoma | Not Observed | Tax associated with apoptosis and p53 | Not a CD4+ T cell malignancy | None | [74] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Rauch, D.A.; Ratner, L. Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients. Viruses 2011, 3, 886-900. https://0-doi-org.brum.beds.ac.uk/10.3390/v3060886
AMA Style
Rauch DA, Ratner L. Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients. Viruses. 2011; 3(6):886-900. https://0-doi-org.brum.beds.ac.uk/10.3390/v3060886
Chicago/Turabian StyleRauch, Daniel A., and Lee Ratner. 2011. "Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients" Viruses 3, no. 6: 886-900. https://0-doi-org.brum.beds.ac.uk/10.3390/v3060886