Conserved Structural Motifs at the C-Terminus of Baculovirus Protein IE0 are Important for its Functions in Transactivation and Supporting hr5-mediated DNA Replication

Abstract
:1. Introduction
2. Results
2.1. Functions of IE0


2.2. IE0 Localizes to the Cell Nucleus

2.3. C-Terminal Mutants of IE0




{kind=link}
{kind=link}
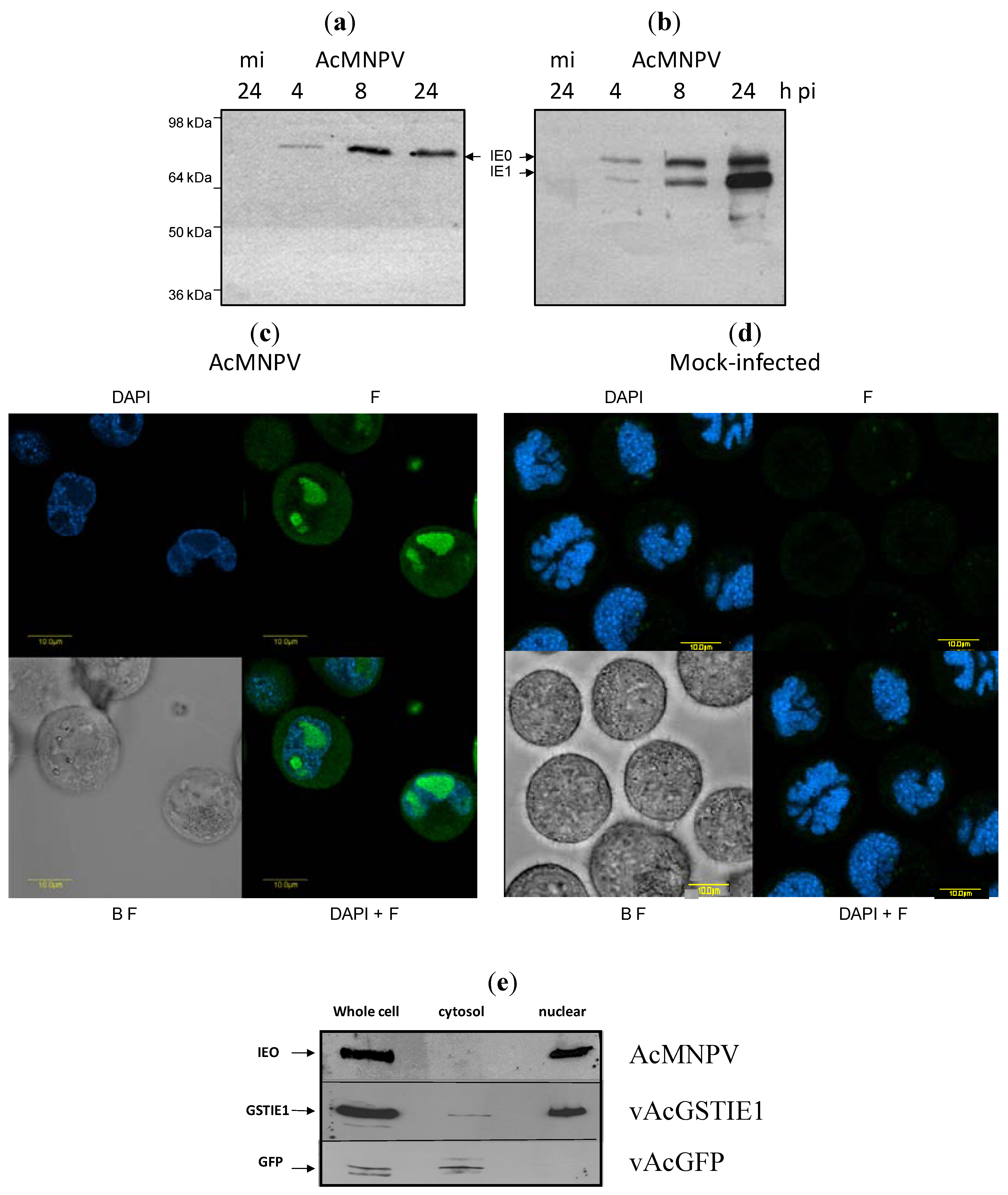{kind=link}
{kind=link}
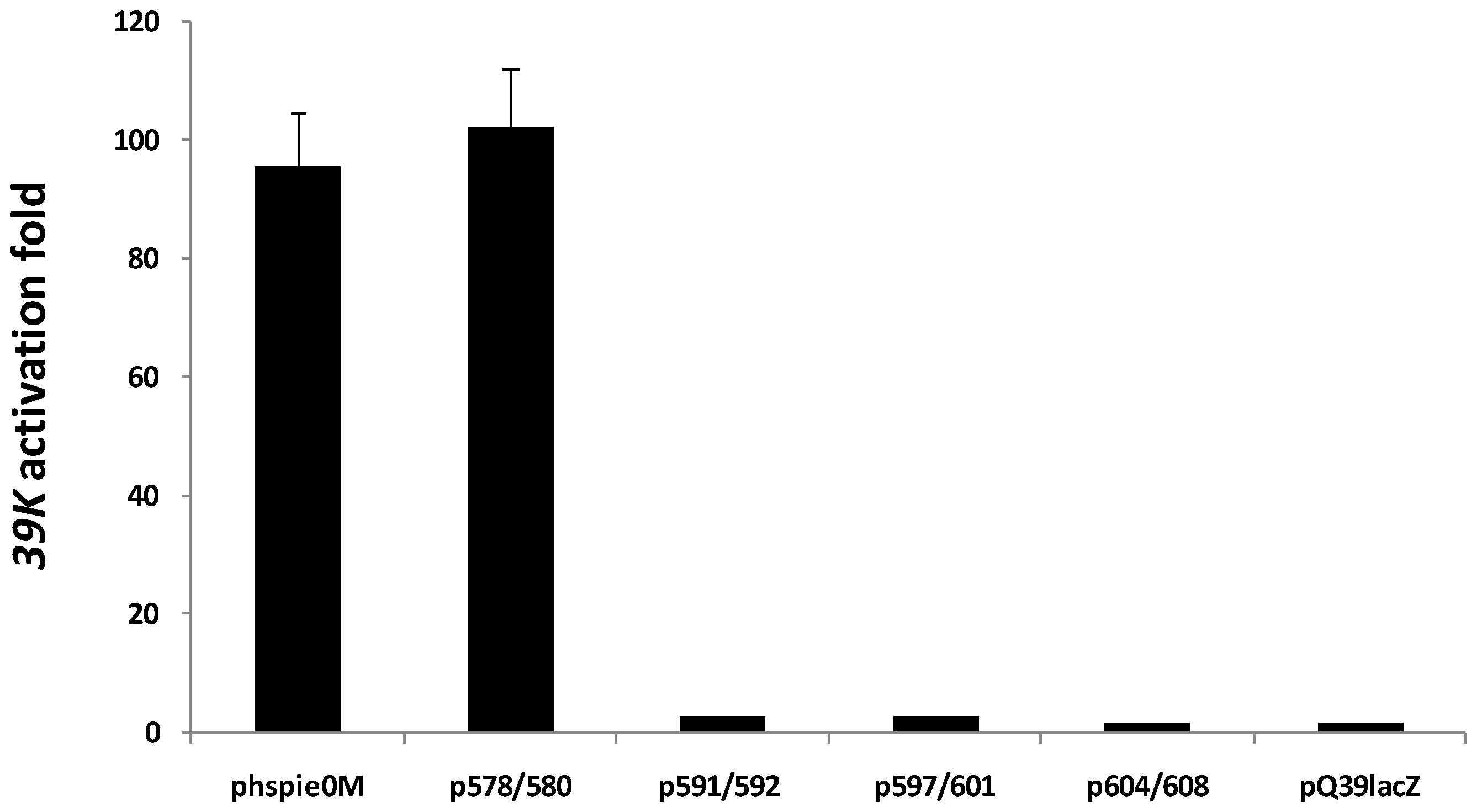{kind=link}
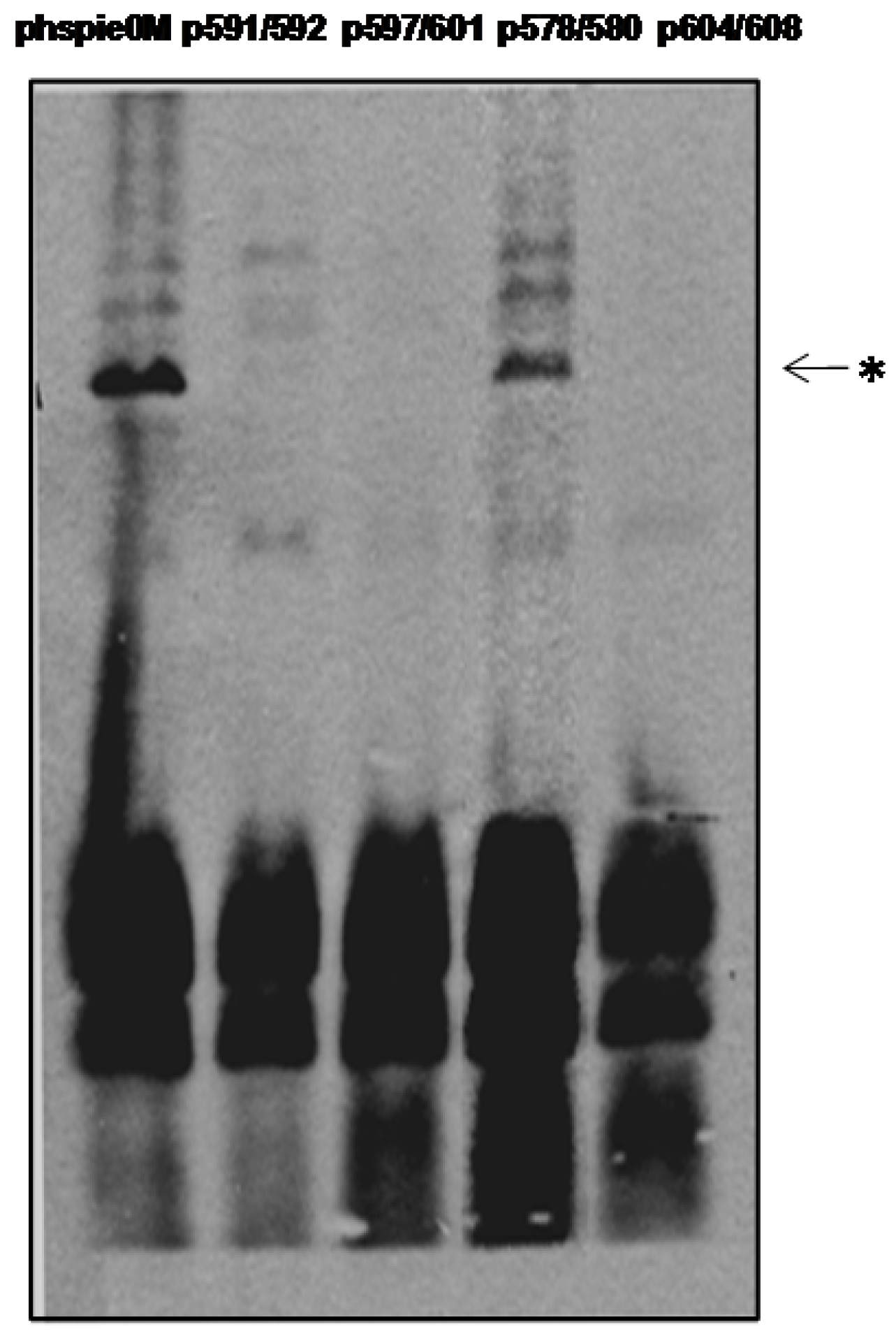{kind=link}
{kind=link}
| IE0-Mutant | Cellular localization | Transactivation of 39K promoter | Replication of hr5-plasmid |
|---|---|---|---|
| R578A/K580A (p578/580) | Nucleus | + | + |
| R591A/R592A (p591/592) | Cytoplasm | − | − |
| L597D/L601E (p597/601) | Cytoplasm | − | − |
| L604D/I608E (p604/608) | Cytoplasm | − | − |

3. Experimental Section
3.1. Cell Lines and Viruses
3.2. Plasmids and Site Directed Mutagenesis
3.3. Preparation of Anti-IE0 Specific Antiserum
3.4. Transient Transfection Assays
3.5. Western Blot Analysis
3.6. Confocal Microscopy
3.7. Biochemical Fractionation
4. Discussion and Conclusions
- Two charged to neutral mutations at the residues 578 and 580 (mutant p578/580), and 591–592 (mutant p591/592) of the BDII motif and,
- Two hydrophobic to charged mutations at residues 597 and 601 (p597/601) and 604 and 608 (p604/608) of the HLH motif.
Acknowledgments
Conflict of Interest
References and Notes
- Carson, D.D.; Guarino, L.A.; Summers, M.D. Functional mapping of an AcNPV immediately early gene which augments expression of the IE-1 trans-activated 39K gene. Virology 1988, 162, 444–451. [Google Scholar]
- Choi, J.; Guarino, L.A. Expression of the IE1 transactivator of Autographa californica nuclear polyhedrosis virus during viral infection. Virology 1995, 209, 99–107. [Google Scholar]
- Friesen, P.D. Regulation of baculovirus early gene expression. In The Baculoviruses; Miller, L.K., Ed.; Plenum: New York, NY, USA, 1997; pp. 141–166. [Google Scholar]
- Guarino, L.A.; Dong, W. Functional dissection of the Autographa californica nuclear polyhedrosis virus enhancer element hr5. Virology 1994, 200, 328–335. [Google Scholar] [CrossRef]
- Huijskens, I.; Li, L.L.; Willis, L.G.; Theilmann, D.A. Role of AcMNPV IE0 in baculovirus very late gene activation. Virology 2004, 323, 120–130. [Google Scholar]
- Kovacs, G.R.; Guarino, L.A.; Summers, M.D. Novel regulatory properties of the IE1 and IE0 transactivators encoded by the baculovirus Autographa californica multicapsid nuclear polyhedrosis virus. J. Virol. 1991, 65, 5281–5288. [Google Scholar]
- Kremer, A.; Knebel-Morsdorf, D. The early baculovirus he65 promoter: On the mechanism of transcriptional activation by IE1. Virology 1998, 249, 336–351. [Google Scholar]
- Passarelli, A.L.; Miller, L.K. Three baculovirus genes involved in late and very late gene expression: ie-1, ie-n, and lef-2. J. Virol. 1993, 67, 2149–2158. [Google Scholar]
- Schultz, K.L.W.; Wetter, J.A.; Fiore, D.C.; Friesen, P.D. Transactivator IE1 is required for baculovirus early replication events that trigger apoptosis in permissive and nonpermissive cells. J. Virol. 2009, 83, 262–272. [Google Scholar]
- Kool, M.; Ahrens, C.H.; Goldbach, R.W.; Rohrmann, G.F.; Vlak, J.M. Identification of genes involved in DNA replication of the Autographa californica baculovirus. Proc. Natl. Acad. Sci. U. S. A. 1994, 91, 11212–11216. [Google Scholar]
- Pathakamuri, J.A.; Theilmann, D.A. The acidic activation domain of the baculovirus transactivator IE1 contains a virus specific domain essential for DNA-replication. J. Virol. 2002, 76, 5598–5604. [Google Scholar]
- Rodems, S.M.; Friesen, P.D. Transcriptional enhancer activity of hr5 requires dual-palindrome half sites that mediate binding of a dimeric form of the baculovirus transregulator IE1. J. Virol. 1995, 69, 5368–5375. [Google Scholar]
- Lu, A.; Miller, L.K. The roles of eighteen baculovirus late expression factor genes in transcription and DNA replication. J. Virol. 1995, 69, 975–982. [Google Scholar]
- Olson, V.A.; Wetter, J.A.; Friesen, P.D. The highly conserved basic domain I of baculovirus IE1 is required for hr enhancer DNA binding and hr-dependent transactivation. J. Virol. 2003, 77, 5668–5677. [Google Scholar]
- Rodems, S.M.; Pullen, S.S.; Friesen, P.D. DNA-dependent transregulation by IE1 of Autographa californica nuclear polyhedrosis virus: IE1 domains required for transactivation and DNA binding. J. Virol. 1997, 71, 9270–9277. [Google Scholar]
- Slack, J.M.; Blissard, G.W. Identification of two independent transcriptional activation domains in the Autographa californica multicapsid nuclear polyhedrosis virus IE1 protein. J. Virol. 1997, 71, 9579–9587. [Google Scholar]
- Okano, K.; Vanarsdall, A.L.; Mikhailov, V.S.; Rohrmann, G.F. Conserved molecular systems of the Baculoviridae. Virology 2006, 344, 77–87. [Google Scholar]
- Kang, W.Y.; Suzuki, M.; Zemskov, E.; Okano, K.; Maeda, S. Characterization of baculovirus repeated open reading frames (bro) in Bombyx mori nucleopolyhedrovirus. J. Virol. 1999, 73, 10339–10345. [Google Scholar]
- Okano, K.; Mikhailov, V.S.; Maeda, S. Colocalization of baculovirus IE-1 and two DNA-binding proteins, DBP and LEF-3, to viral replication factories. J. Virol. 1999, 73, 110–119. [Google Scholar]
- Olson, V.A.; Wetter, J.A.; Friesen, P.D. Oligomerization mediated by a helix-loop-helix-like domain of baculovirus IE1 is required for early promoter transactivation. J. Virol 2001, 75, 6042–6051. [Google Scholar]
- Olson, V.A.; Wetter, J.A.; Friesen, P.D. Baculovirus transregulator IE1 requires a dimeric nuclear localization element for nuclear import and promoter activation. J. Virol. 2002, 76, 9505–9515. [Google Scholar]
- Theilmann, D.A.; Willis, L.G.; Bosch, B.J.; Forsythe, I.J.; Li, Q.J. The baculovirus transcriptional transactivator ie0 produces multiple products by internal initiation of translation. Virology 2001, 290, 211–223. [Google Scholar]
- Leisy, D.J.; Rasmussen, C.; Kim, H.T.; Rohrmann, G.F. The Autographa californica nuclear polyhedrosis virus homologous region 1a: Identical sequences are essential for DNA replication activity and transcriptional enhancer function. Virology 1995, 208, 742–752. [Google Scholar] [CrossRef]
- Nagamine, T.; Kawasaki, Y.; Iizuka, T.; Matsumoto, S. Focal distribution of baculovirus IE1 triggered by its binding to the hr DNA elements. J. Virol. 2005, 79, 39–46. [Google Scholar]
- Chisholm, G.E.; Henner, D.J. Multiple early transcripts and splicing of the Autographa californica nuclear polyhedrosis virus IE-1 gene. J. Virol. 1988, 62, 3193–3200. [Google Scholar]
- Kovacs, G.R.; Guarino, L.A.; Graham, B.L.; Summers, M.D. Identification of spliced baculovirus RNAs expressed late in infection. Virology 1991, 185, 633–643. [Google Scholar]
- Dai, X.J.; Willis, L.G.; Huijskens, I.; Palli, S.R.; Theilmann, D.A. The acidic activation domains of the baculovirus transactivators IE1 and IE0 are functional for transcriptional activation in both insect and mammalian cells. J. Gen. Virol. 2004, 85, 573–582. [Google Scholar]
- Lu, L.Q.; Rivkin, H.; Chejanovsky, N. The immediate-early protein IE0 of the Autographa californica nucleopolyhedrovirus is not essential for viral replication. J. Virol. 2005, 79, 10077–10082. [Google Scholar] [CrossRef]
- Stewart, T.M.; Huijskens, I.; Willis, L.G.; Theilmann, D.A. The Autographa californica multiple nucleopolyhedrovirus ie0-ie1 gene complex is essential for wild-type virus replication, but either IE0 or IE1 can support virus growth. J. Virol. 2005, 79, 4619–4629. [Google Scholar]
- Lu, L.Q.; Du, Q.S.; Chejanovsky, N. Reduced expression of the immediate-early protein IE0 enables efficient replication of Autographa californica multiple nucleopolyhedrovirus in poorly permissive Spodoptera littoralis cells. J. Virol. 2003, 77, 535–545. [Google Scholar] [CrossRef]
- Guarino, L.A.; Smith, M. Regulation of delayed-early gene transcription by dual TATA boxes. J. Virol. 1992, 66, 3733–3739. [Google Scholar]
- Guarino, L.A.; Summers, M.D. Interspersed homologous DNA of Autographa californica nuclear polyhedrosis virus enhances delayed-early gene expression. J. Virol. 1986, 60, 215–223. [Google Scholar]
- Xu, B.; Yoo, S.; Guarino, L.A. Differential transcription of baculovirus late and very late promoters: Fractionation of nuclear extracts by phosphocellulose chromatography. J. Virol. 1995, 69, 2912–2917. [Google Scholar]
- Passarelli, A.L.; Guarino, L.A. Baculovirus late and very late gene regulation. Curr. Drug Targets 2007, 8, 1103–1115. [Google Scholar]
- Smith, G.E.; Summers, M.D. Analysis of baculovirus genomes with restriction endonucleases. Virology 1978, 89, 517–527. [Google Scholar]
- Summers, M.D.; Smith, G.E. A Manual of Methods for Baculovirus Vectors and Insect Cell-Culture Procedures; Texas Agricultural Experiment Station Bulletin: College Station, TX, USA, 1987. [Google Scholar]
- Rivkin, H.; Kroemer, J.A.; Bronshtein, A.; Belausov, E.; Webb, B.A.; Chejanovsky, N. Response of immunocompetent and immunosuppressed Spodoptera littoralis larvae to baculovirus infection. J. Gen. Virol. 2006, 87, 2217–2225. [Google Scholar] [CrossRef]
- Guarino, L.A.; Summers, M.D. Nucleotide sequence and temporal expression of a baculovirus regulatory gene. J. Virol. 1987, 61, 2091–2099. [Google Scholar]
- Pei, Z.F.; Reske, G.; Huang, Q.H.; Hammock, B.D.; Qi, Y.P.; Chejanovsky, N. Characterization of the apoptosis suppressor protein P49 from the Spodoptera littoralis nucleopolyhedrovirus. J. Biol. Chem. 2002, 277, 48677–48684. [Google Scholar]
- Regev, A.; Rivkin, H.; Gurevitz, M.; Chejanovsky, N. New measures of insecticidal efficacy and safety obtained with the 39K promoter of a recombinant baculovirus. FEBS Lett. 2006, 580, 6777–6782. [Google Scholar] [CrossRef]
- Kool, M.; Ahrens, C.H.; Vlak, J.M.; Rohrmann, G.F. Replication of baculovirus DNA. J. Gen. Virol. 1995, 76, 2103–2118. [Google Scholar]
- Lu, A.; Miller, L.K. Differential requirements for baculovirus late expression factor genes in two cell lines. J. Virol. 1995, 69, 6265–6272. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Luria, N.; Lu, L.; Chejanovsky, N. Conserved Structural Motifs at the C-Terminus of Baculovirus Protein IE0 are Important for its Functions in Transactivation and Supporting hr5-mediated DNA Replication. Viruses 2012, 4, 761-776. https://0-doi-org.brum.beds.ac.uk/10.3390/v4050761
Luria N, Lu L, Chejanovsky N. Conserved Structural Motifs at the C-Terminus of Baculovirus Protein IE0 are Important for its Functions in Transactivation and Supporting hr5-mediated DNA Replication. Viruses. 2012; 4(5):761-776. https://0-doi-org.brum.beds.ac.uk/10.3390/v4050761
Chicago/Turabian StyleLuria, Neta, Liqun Lu, and Nor Chejanovsky. 2012. "Conserved Structural Motifs at the C-Terminus of Baculovirus Protein IE0 are Important for its Functions in Transactivation and Supporting hr5-mediated DNA Replication" Viruses 4, no. 5: 761-776. https://0-doi-org.brum.beds.ac.uk/10.3390/v4050761