
Tsv-N1: A Novel DNA Algal Virus that Infects Tetraselmis striata

Abstract
:1. Introduction
2. Materials and Methods
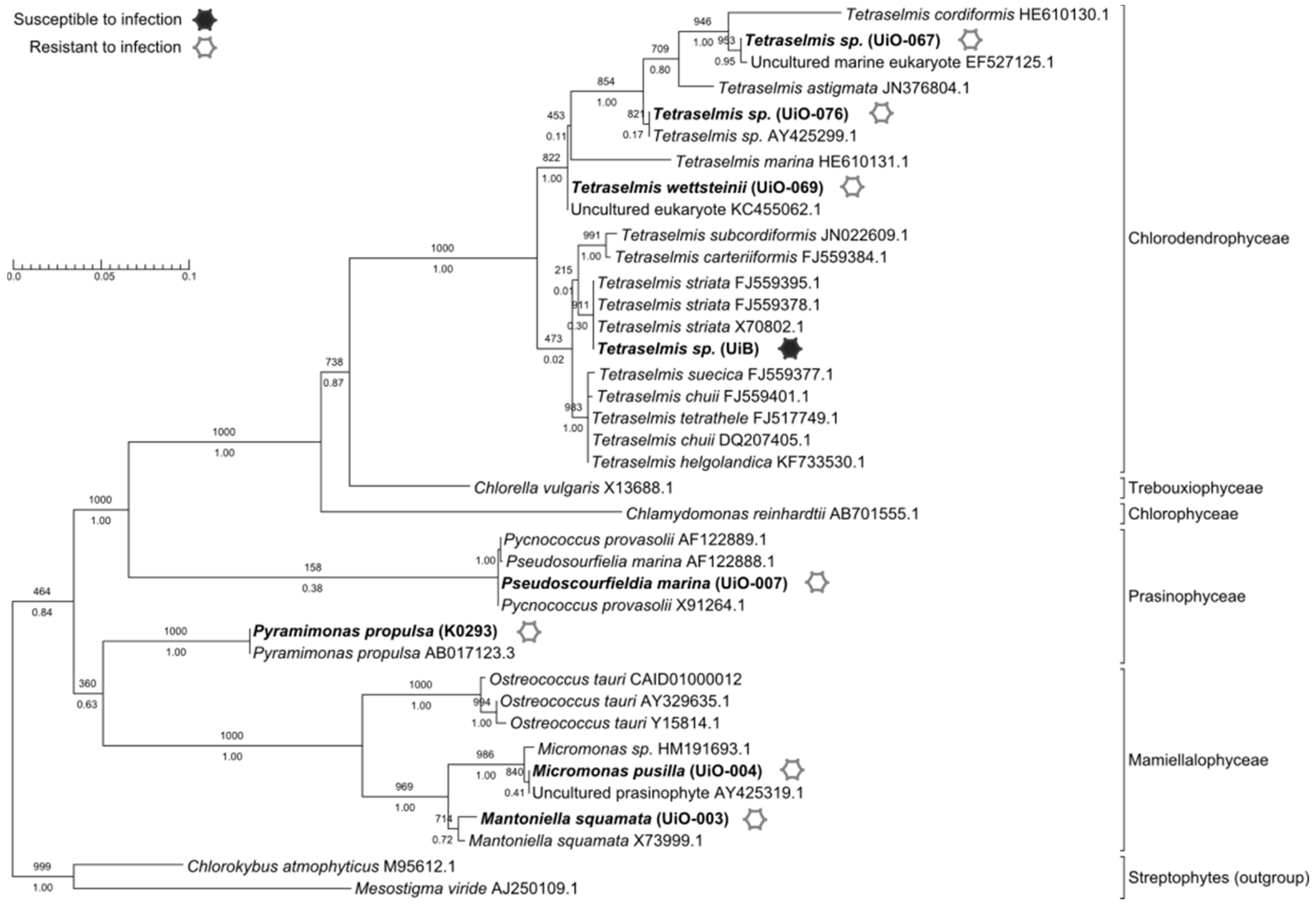
2.1. Testing Host Range

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | F/R | Sequence (5′-3′) | Targeted Gene | Hybridization Temperature for qPCR (°C) |
|---|---|---|---|---|
| Euk1a | F | CTGGTTGATCCTGCCAG | 18S rDNA | n/a |
| Euk516r | R | ACCAGACTTGCCCTCC | ||
| CDP_F339 | F | AAACATCACCGTGCCAACAC | TsV_14 Putative Capsid Decoration Protein | 58 |
| CDP_R450 | R | ATGAGCTTTCTTCGCGTACC | ||
| DNAPol_F83 | F | ATAAGCCCGAGCCGAAAAAG | TsV_19 Putative DNA Polymerase | 59 |
| DNAPol_R203 | R | AATCCGAGATTCAGCTCCAGTG |
2.2. Infection Dynamics
2.3. TsV-N1 Genomic Analysis
2.4. Environmental Metagenomic Analysis
3. Results
3.1. Host Range

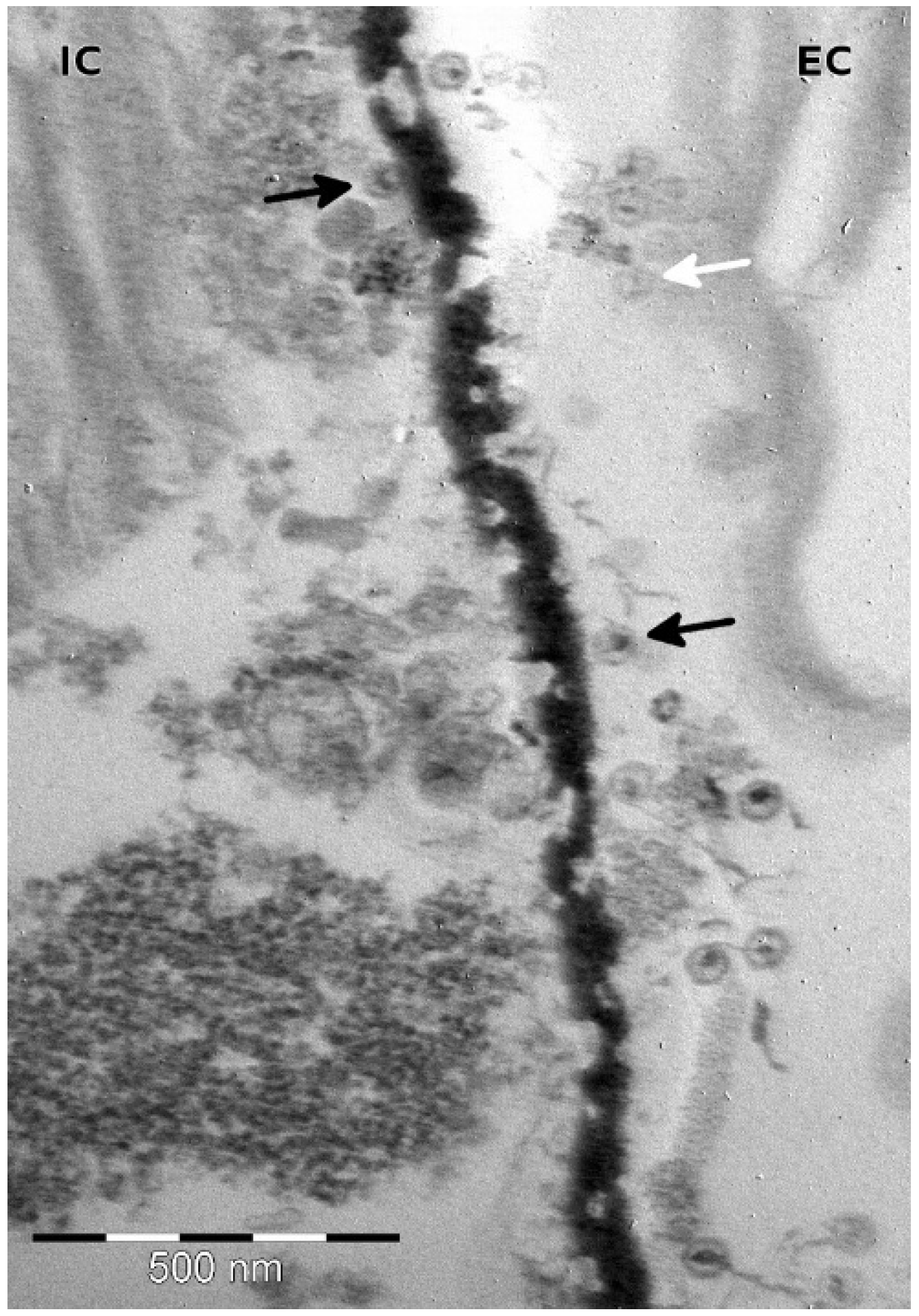
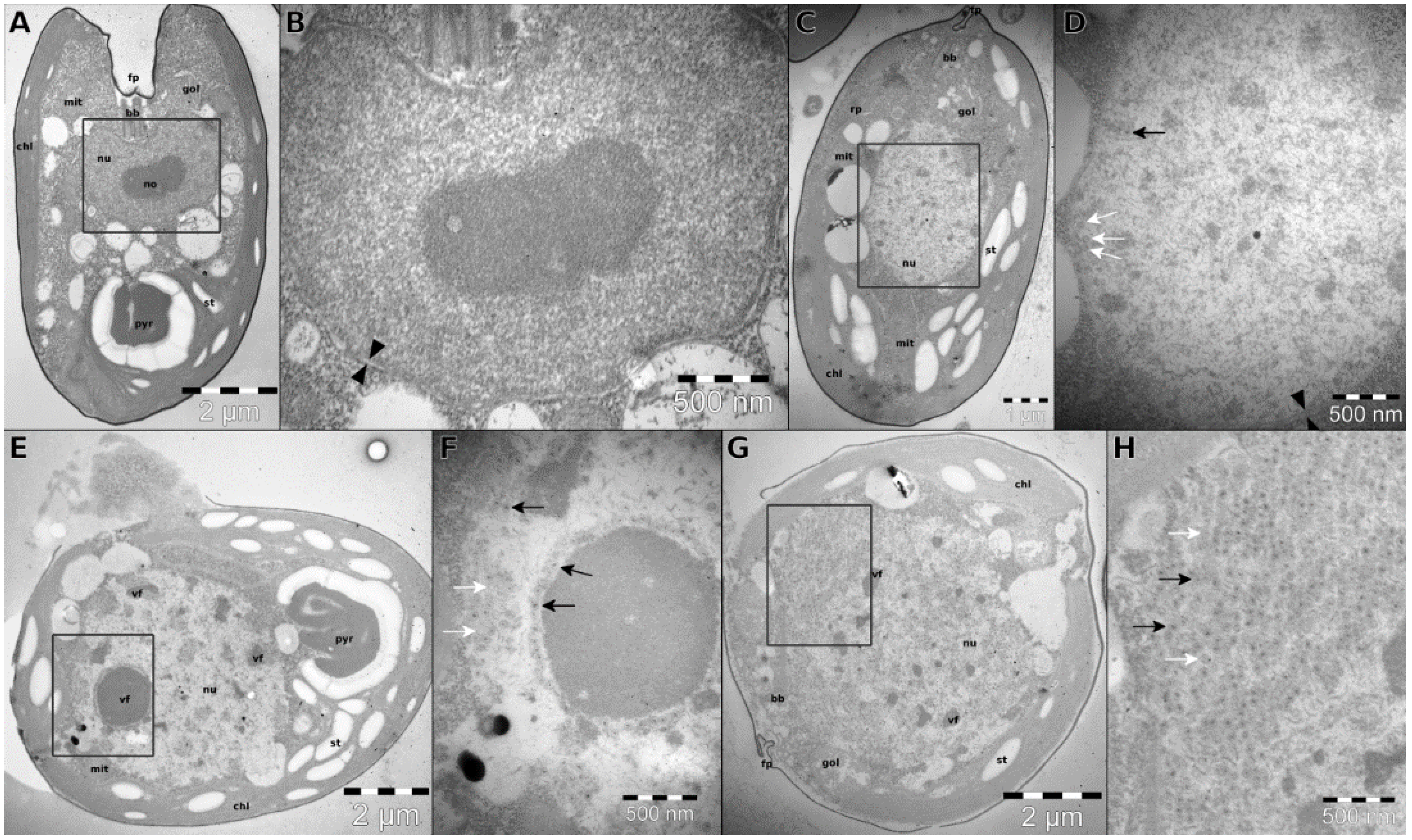
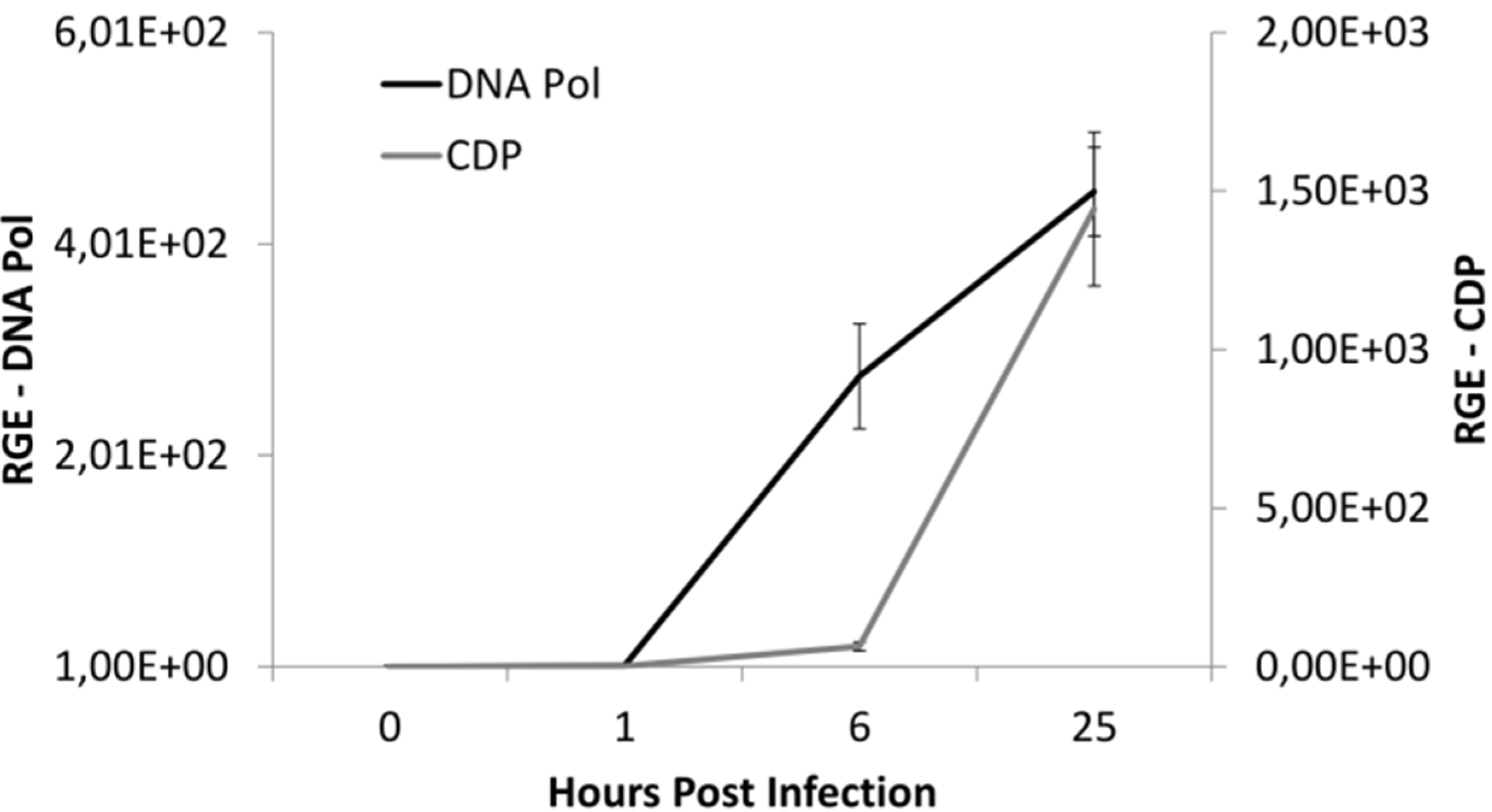
3.2. Infection Dynamics



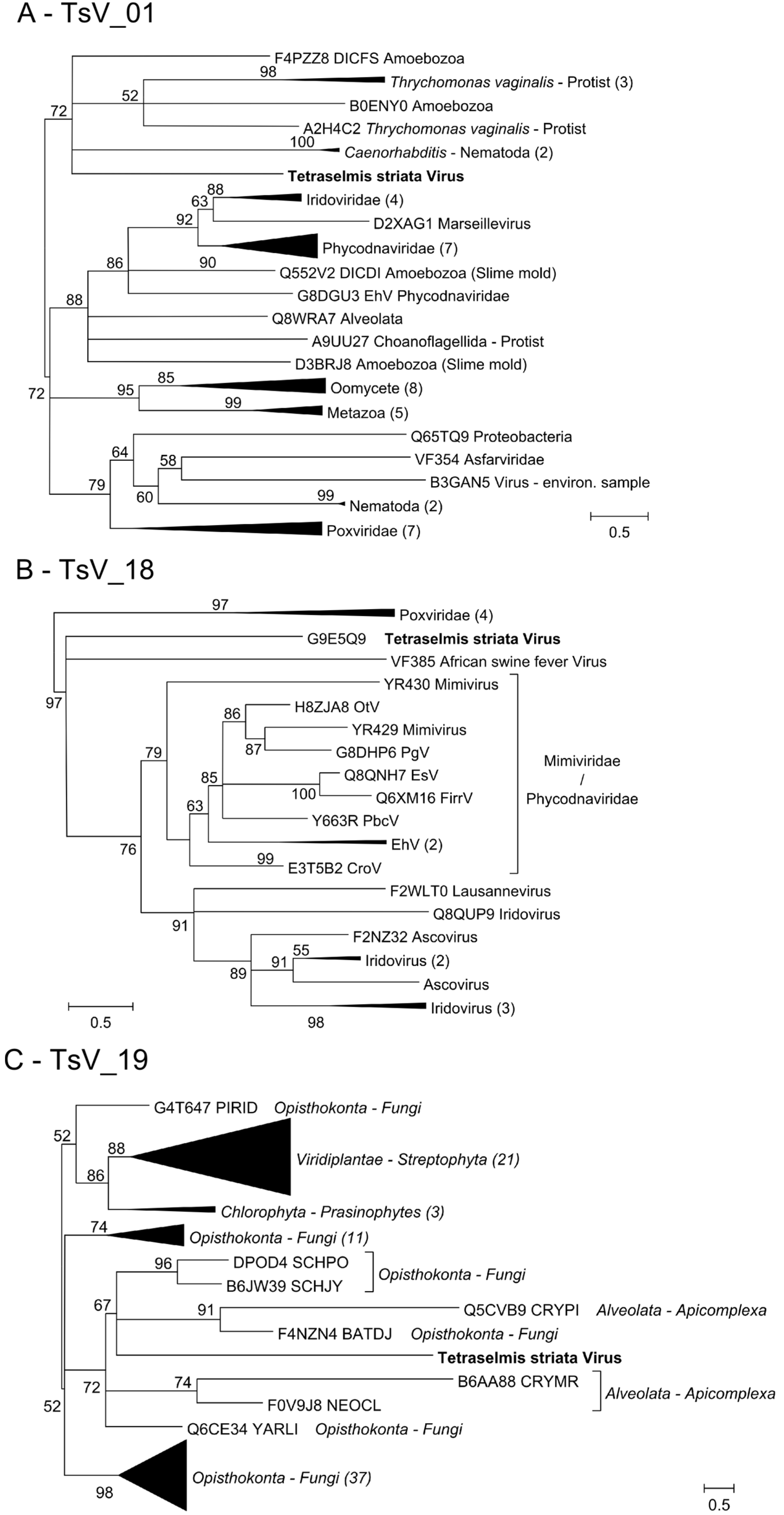
3.3. TsV-N1 Genomic Analysis
3.4. Detection of TV-N1 and Tetraselmis striata Reads in Environmental Datasets

| Gene | Conserved Domains | Pfam | Consistent Phylogeny? | Monophyletic Phycodnavirus | Closest Homology | Psi-Blast Top Hit | ||
|---|---|---|---|---|---|---|---|---|
| Top Hit | E-value | Taxonomy | ||||||
| TsV_01 | Poxvirus A32 protein | PF04665 | yes | yes | Trichomonas vaginalis Eukaryota | — | — | — |
| TsV_18 | Poxvirus Late Transcription Factor VLTF3 like | PF04947 | yes | yes | Eukaryotic Virus | — | — | |
| TsV_19 | DNA polymerase delta, subunit 4 | PF04081 | yes | yes | Fungi-Eukaryota | — | — | — |
| TsV_04 | Apolipoprotein L | PF05461 | no | — | Mammalia | Microtus ochrogaster | 2.0 × 10−11 | Eukaryota-Mammalia |
| TsV_07 | GP16 | — | no | — | — | Mycobacterium Phage Marcell | 1.1 × 10−2 | Virus-Caudovirales |
| TsV_08 | ATP-dependent DNA helicase PIF1 | PF13604 | no | — | — | Candida albicans | 1.2 × 10−2 | Eukaryota-Fungi |
| TsV_010 | — | — | no | — | — | Chlorella variabillis | 1.0 × 10−3 | Eukaryota-Chlorophyta |
| TsV_14 | Capsid decoration protein | PF02924 | no | — | — | Prochlorococcus phage P-SSM4 | 1.0 × 10−2 | Virus-Caudovirales |
| TsV_25 | Coagulation factor 5/8 C-terminal type domain | PF00754 | no | — | — | Pectobacterium carotovorum | 9.0 × 10−8 | Bacteria Enterobacteriaceae |
4. Discussion
Perspectives and Implications
Supplementary Files
Supplementary File 1Acknowledgements
Author Contributions
Conflicts of Interest
References
- Short, S.M. The ecology of viruses that infect eukaryotic algae. Environ. Microbiol. 2012, 14, 2253–2271. [Google Scholar] [CrossRef] [PubMed]
- Suttle, C.A. Marine viruses—Major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M. Marine viruses: Truth or dare. Ann. Rev. Mar. Sci. 2012, 4, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, C.P.D.; Wilhelm, S.W.; Thingstad, F.; Weinbauer, M.G.; Bratbak, G.; Heldal, M.; Kimmance, S.A.; Middelboe, M.; Nagasaki, K.; Paul, J.H.; et al. Global-scale processes with a nanoscale drive: The role of marine viruses. Int. Soc. Microb. Ecol. 2008, 2, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, F.; Thurber, R.V. Viruses manipulate the marine environment. Nature 2009, 459, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Monier, A.; Claverie, J.M.; Ogata, H. Taxonomic distribution of large DNA viruses in the sea. Genome Biol. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Williamson, S.J.; Allen, L.Z.; Lorenzi, H.A.; Fadrosh, D.W.; Brami, D.; Thiagarajan, M.; McCrow, J.P.; Tovchigrechko, A.; Yooseph, S.; Venter, J.C. Metagenomic exploration of viruses throughout the Indian Ocean. PLoS ONE 2012, 7, e42047. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Grimsley, N.; Escande, M.L.; Subirana, L.; Derelle, E.; Moreau, H. Acquisition and maintenance of resistance to viruses in eukaryotic phytoplankton populations. Environ. Microbiol. 2011, 13, 1412–1420. [Google Scholar] [CrossRef] [PubMed]
- Avrani, S.; Lindell, D. Convergent evolution toward an improved growth rate and a reduced resistance range in prochlorococcus strains resistant to phage. Proc. Natl. Acad. Sci. USA 2015, 112, 2191–2200. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, S.; Liu, T.; Sun, J.; Chi, S.; Liu, C.; Li, X.; Yin, J.; Wang, X.; Yu, J. Endogenous viral elements in algal genomes. Acta Ocean. Sin. 2014, 33, 102–107. [Google Scholar] [CrossRef]
- Suttle, C. The viriosphere: The greatest biological diversity on earth and driver of global processes. Environ. Microbiol. 2005, 7, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Monier, A.; Pagarete, A.; de Vargas, C.; Allen, M.J.; Read, B.; Claverie, J.-M.; Ogata, H. Horizontal gene transfer of an entire metabolic pathway between a eukaryotic alga and its DNA virus. Genome Res. 2009, 19, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Grimsley, N.H.; Thomas, R.; Kegel, J.U.; Jacquet, S.; Moreau, H.; Desdevises, Y. Genomics of algal host-virus interactions. In Advances in Botanical Research; Piganeau, G., Ed.; Academic Press Elsevier Science LTD: San Diego, CA, USA, 2012; Volume 64, pp. 343–381. [Google Scholar]
- Bellec, L.; Clerissi, C.; Edern, R.; Foulon, E.; Simon, N.; Grimsley, N.; Desdevises, Y. Cophylogenetic interactions between marine viruses and eukaryotic picophytoplankton. BMC Evol. Biol. 2014, 14, e59. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.E.; Chan, A.M.; Suttle, C.A. A novel virus (HaNIV) causes lysis of the toxic bloom-forming alga Heterosigma akashiwo (Raphidophyceae). J. Phycol. 2001, 37, 216–222. [Google Scholar] [CrossRef]
- Lee, H.J.; Hur, S.B. Genetic relationships among multiple strains of the genus Tetraselmis based on partial 18S rDNA sequences. Algae 2009, 24, 205–212. [Google Scholar] [CrossRef]
- Mani, A.; Anil, A.C.; Leliaert, F.; Delany, J.; Mesbahi, E. Tetraselmis indica (Chlorodendrophyceae, Chlorophyta), a new species isolated from salt pans in Goa, India. Eur. J. Phycol. 2013, 48, 61–78. [Google Scholar]
- Serodio, J.; Silva, R.; Ezequiel, J.; Calado, R. Photobiology of the symbiotic acoel flatworm Symsagittifera roscoffensis: Algal symbiont photoacclimation and host photobehaviour. J. Mar. Biol. Assoc. UK 2011, 91, 163–171. [Google Scholar] [CrossRef]
- Hori, T.; Norris, R.; Chihara, M. Studies on the ultrastructure and taxonomy of the genus Tetraselmis (Prasinophyceae). Bot. Mag. Tokyo 1982, 95, 49–61. [Google Scholar] [CrossRef]
- D’Adamo, S.; Jinkerson, R.E.; Boyd, E.S.; Brown, S.L.; Baxter, B.K.; Peters, J.W.; Posewitz, M.C. Evolutionary and biotechnological implications of robust hydrogenase activity in halophilic strains of Tetraselmis. PLoS ONE 2014, 9, e85812. [Google Scholar] [CrossRef] [PubMed]
- Malik, A. Metal bioremediation through growing cells. Environ. Int. 2004, 30, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, O.A.; Boyko, A.L.; Gordienko, A.I.; Sherban, S.A.; Shevchenko, T.P.; Polischuck, V.P. Characteristics of virus of Tetraselmis viridis norris (Chorophyta, Prasinophycea). Dokl. Akad. Nauk. Ukr. 2005, 1, 158–162. [Google Scholar]
- Sogin, M.L.; Gunderson, J.H. Structural diversity of eukaryotic small subunit ribosomal RNasA. Ann. N. Y. Acad. Sci. 1987, 503, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16s rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The silva ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the www for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [PubMed]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schäffer, A.A.; Yu, Y.-K. Protein database searches using compositionally adjusted substitution matrices. FEBS J. 2005, 272, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. Interproscan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef] [PubMed]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Biegert, A.; Mayer, C.; Remmert, M.; Söding, J.; Lupas, A.N. The MPI Bioinformatics Toolkit for protein sequence analysis. Nucleic Acids Res. 2006, 34, W335–W339. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling expedition: northwest Atlantic through eastern tropical Pacific. PLoS Biol. 2007, 5, e77. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, O.A.; Heldal, M.; Sandaa, R.A.; Bratbak, G. The first isolation of Tetraselmis viridis virus (TvV-N1) from Norway environment. In Proceedings of the V International Conference on Bioresourses and Viruses, Kiev, Ukraine, 10–13 September 2007; p. 93.
- Hurwitz, B.L.; Deng, L.; Poulos, B.T.; Sullivan, M.B. Evaluation of methods to concentrate and purify ocean virus communities through comparative, replicated metagenomics. Environ. Microbiol. 2013, 15, 1428–1440. [Google Scholar] [CrossRef] [PubMed]
- Clerissi, C.; Desdevises, Y.; Grimsley, N. Prasinoviruses of the marine green alga Ostreococcus tauri are mainly species specific. J. Virol. 2012, 86, 4611–4619. [Google Scholar] [CrossRef] [PubMed]
- Johannessen, T.V.; Bratbak, G.; Larsen, A.; Ogata, H.; Egge, E.S.; Edvardsen, B.; Eikrem, W.; Sandaa, R.-A. Characterisation of three novel giant viruses reveals huge diversity among viruses infecting Prymnesiales (Haptophyta). Virology 2015, 476, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Baudoux, A.C.; Brussaard, C.P. Characterization of different viruses infecting the marine harmful algal bloom species Phaeocystis globosa. Virology 2005, 341, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.H.; van Etten, J.L.; Allen, M.J. The phycodnaviridae: The story of how tiny giants rule the world. Curr. Top. Microbiol. Immunol. 2009, 328, 1–42. [Google Scholar] [PubMed]
- Van Etten, J.L.; Graves, M.V.; Muller, D.G.; Boland, W.; Delaroque, N. Phycodnaviridae—Large DNA algal viruses. Arch. Virol. 2002, 147, 1479–1516. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, J.L.; Lane, L.C.; Dunigan, D.D. DNA viruses: The really big ones (giruses). Ann. Rev. Microbiol. 2010, 64, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Aravind, L.; Koonin, E.V. Common origin of four diverse families of large eukaryotic DNA viruses. J. Virol. 2001, 75, 11720–11734. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Balaji, S.; Koonin, E.V.; Aravind, L. Evolutionary genomics of nucleo-cytoplasmic large DNA viruses. Virus Res. 2006, 117, 156–184. [Google Scholar] [CrossRef] [PubMed]
- Tai, V.; Lawrence, J.E.; Lang, A.S.; Chan, A.M.; Culley, A.I.; Suttle, C.A. Characterization of HaRNAV, a single-stranded RNA virus causing lysis of Heterosigma akashiwo (Raphidophyceae). J. Phycol. 2003, 39, 343–352. [Google Scholar] [CrossRef]
- Peters, B.M.; Yano, J.; Noverr, M.C.; Fidel, P.L., Jr. Candida vaginitis: When opportunism knocks, the host responds. PLoS Pathog. 2014, 10, e1003965. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, O.A.; Solovyova, Y.V.; Solovyov, A.V. Results of algae viruses search in human clinical material. Ukr. Bioorganica Acta 2011, 9, 53–56. [Google Scholar]
- Popgeorgiev, N.; Boyer, M.; Fancello, L.; Monteil, S.; Robert, C.; Rivet, R.; Nappez, C.; Azza, S.; Chiaroni, J.; Raoult, D.; et al. Marseillevirus-like virus recovered from blood donated by asymptomatic humans. J. Infect. Dis. 2013, 208, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Yolken, R.H.; Jones-Brando, L.; Dunigan, D.D.; Kannan, G.; Dickerson, F.; Severance, E.; Sabunciyan, S.; Talbot, C.C.; Prandovszky, E.; Gurnon, J.R.; et al. Chlorovirus ATCV-1 is part of the human oropharyngeal virome and is associated with changes in cognitive functions in humans and mice. Proc. Natl. Acad. Sci. USA 2014, 111, 16106–16111. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagarete, A.; Grébert, T.; Stepanova, O.; Sandaa, R.-A.; Bratbak, G. Tsv-N1: A Novel DNA Algal Virus that Infects Tetraselmis striata. Viruses 2015, 7, 3937-3953. https://0-doi-org.brum.beds.ac.uk/10.3390/v7072806
Pagarete A, Grébert T, Stepanova O, Sandaa R-A, Bratbak G. Tsv-N1: A Novel DNA Algal Virus that Infects Tetraselmis striata. Viruses. 2015; 7(7):3937-3953. https://0-doi-org.brum.beds.ac.uk/10.3390/v7072806
Chicago/Turabian StylePagarete, António, Théophile Grébert, Olga Stepanova, Ruth-Anne Sandaa, and Gunnar Bratbak. 2015. "Tsv-N1: A Novel DNA Algal Virus that Infects Tetraselmis striata" Viruses 7, no. 7: 3937-3953. https://0-doi-org.brum.beds.ac.uk/10.3390/v7072806