
Recombinant Pseudorabies Virus (PRV) Expressing Firefly Luciferase Effectively Screened for CRISPR/Cas9 Single Guide RNAs and Antiviral Compounds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Viruses
2.2. Generation of the HR Plasmid and CRISPR/Cas9 sgRNA Plasmids
2.3. Recombination and Purification of the Luciferase Tagged PRV
2.4. In Vitro Growth Properties
2.5. Transfection and Western Blot
2.6. CRISPR/Cas9 sgRNA Screening and Antiviral Assay
3. Results
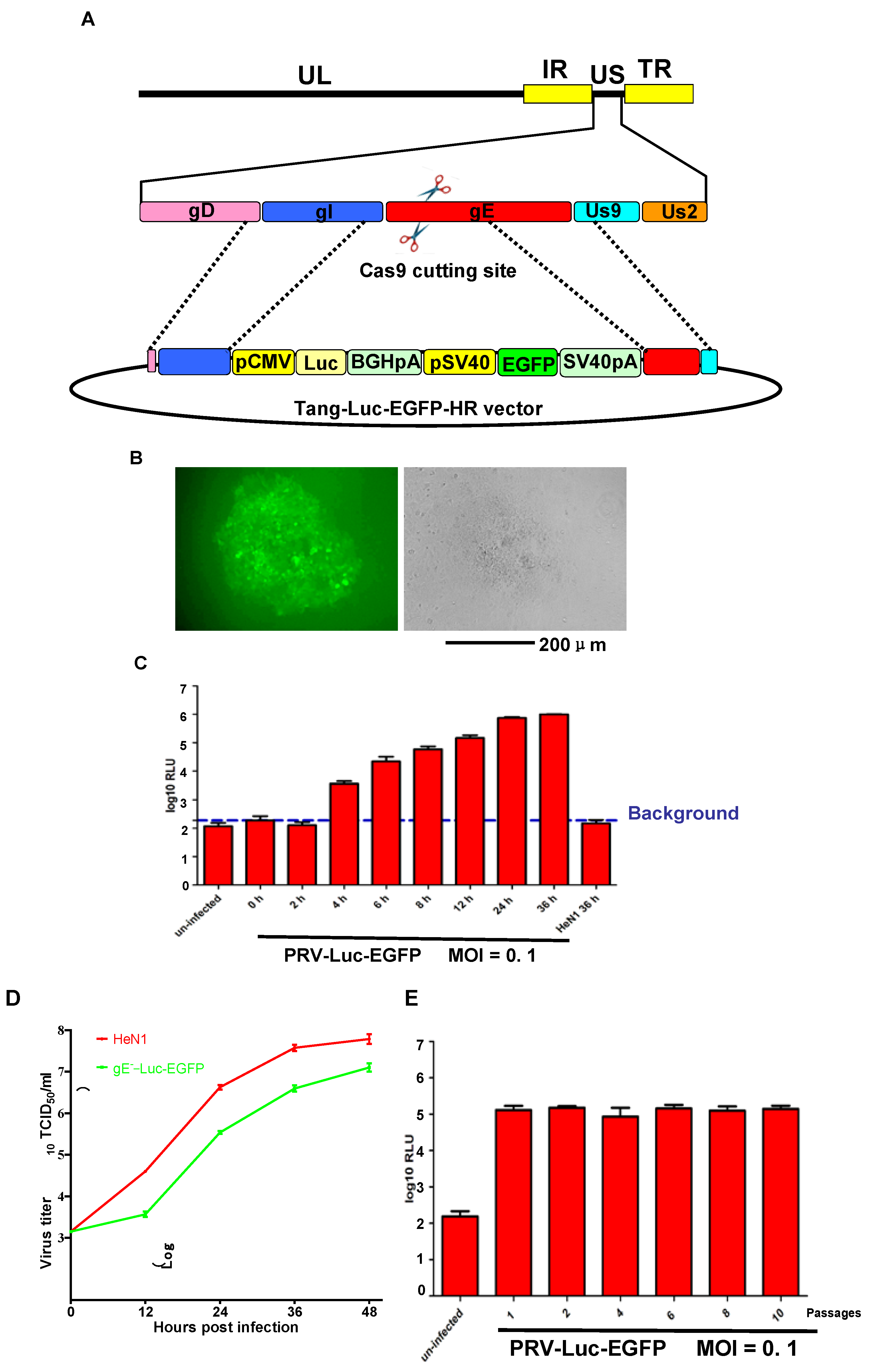
3.1. PRV-Luc-EGFP was Successfully Constructed
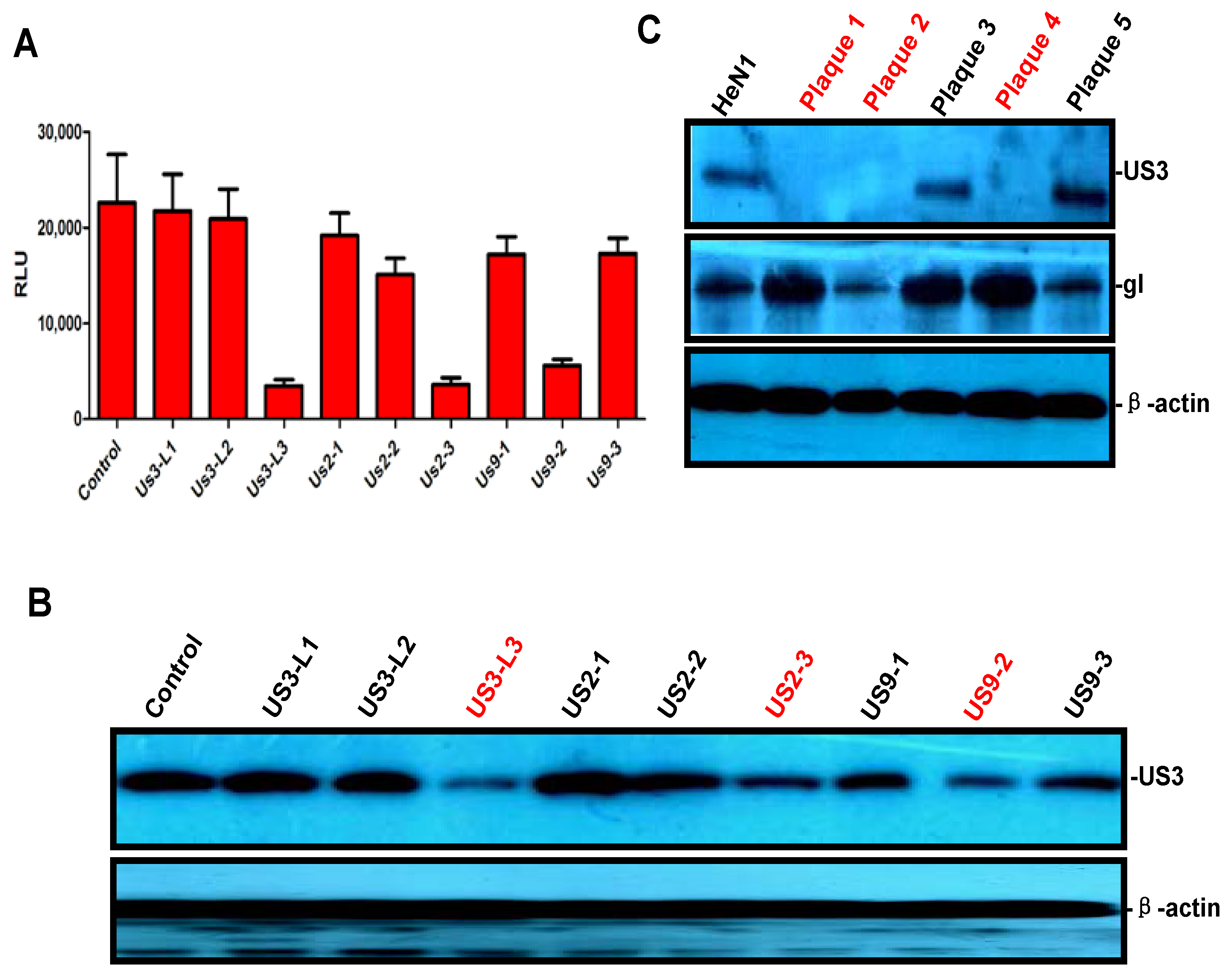
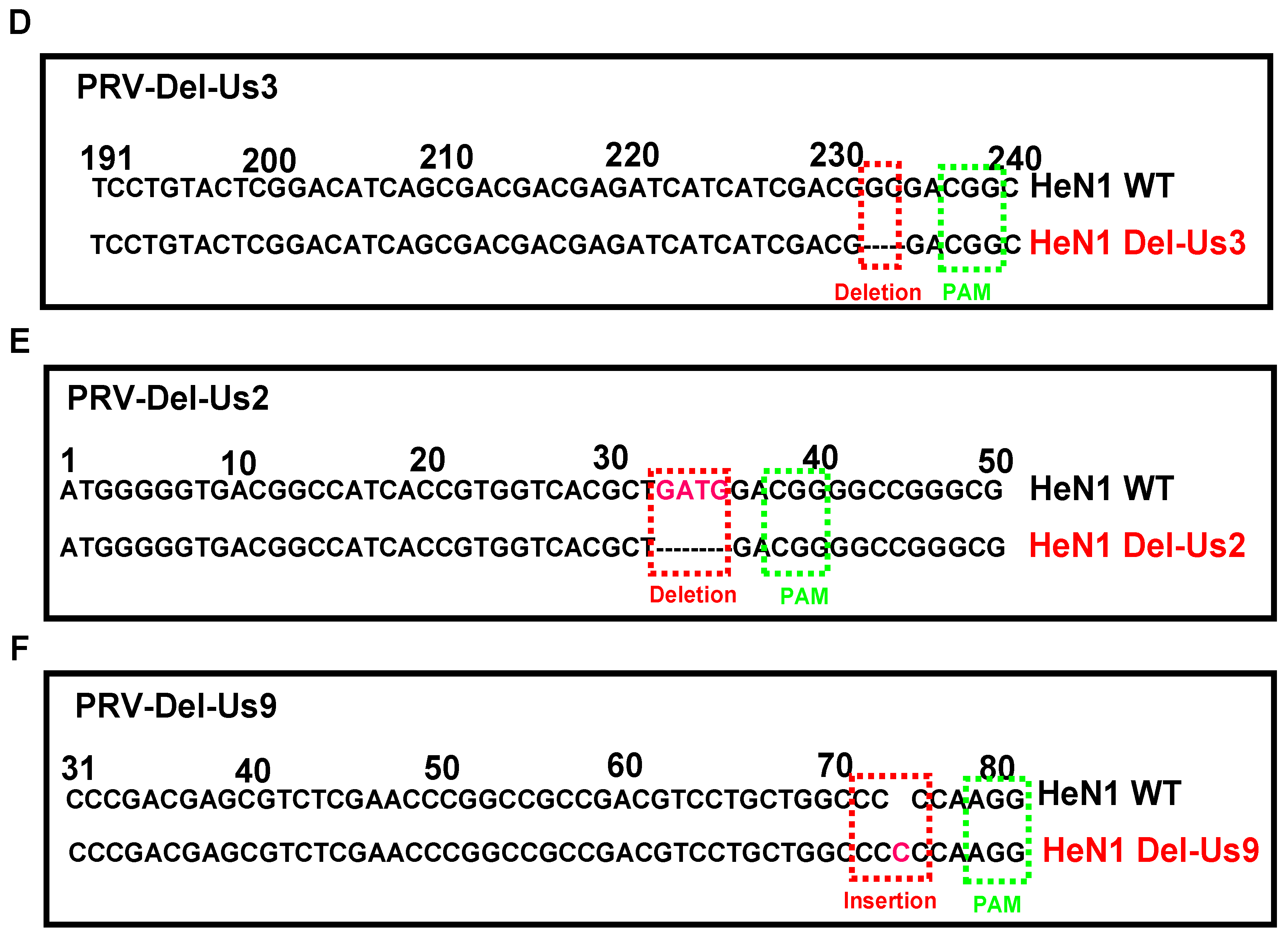
3.2. PRV-Luc-EGFP Was an Effective Tool for CRISPR/Cas9 sgRNA Screening
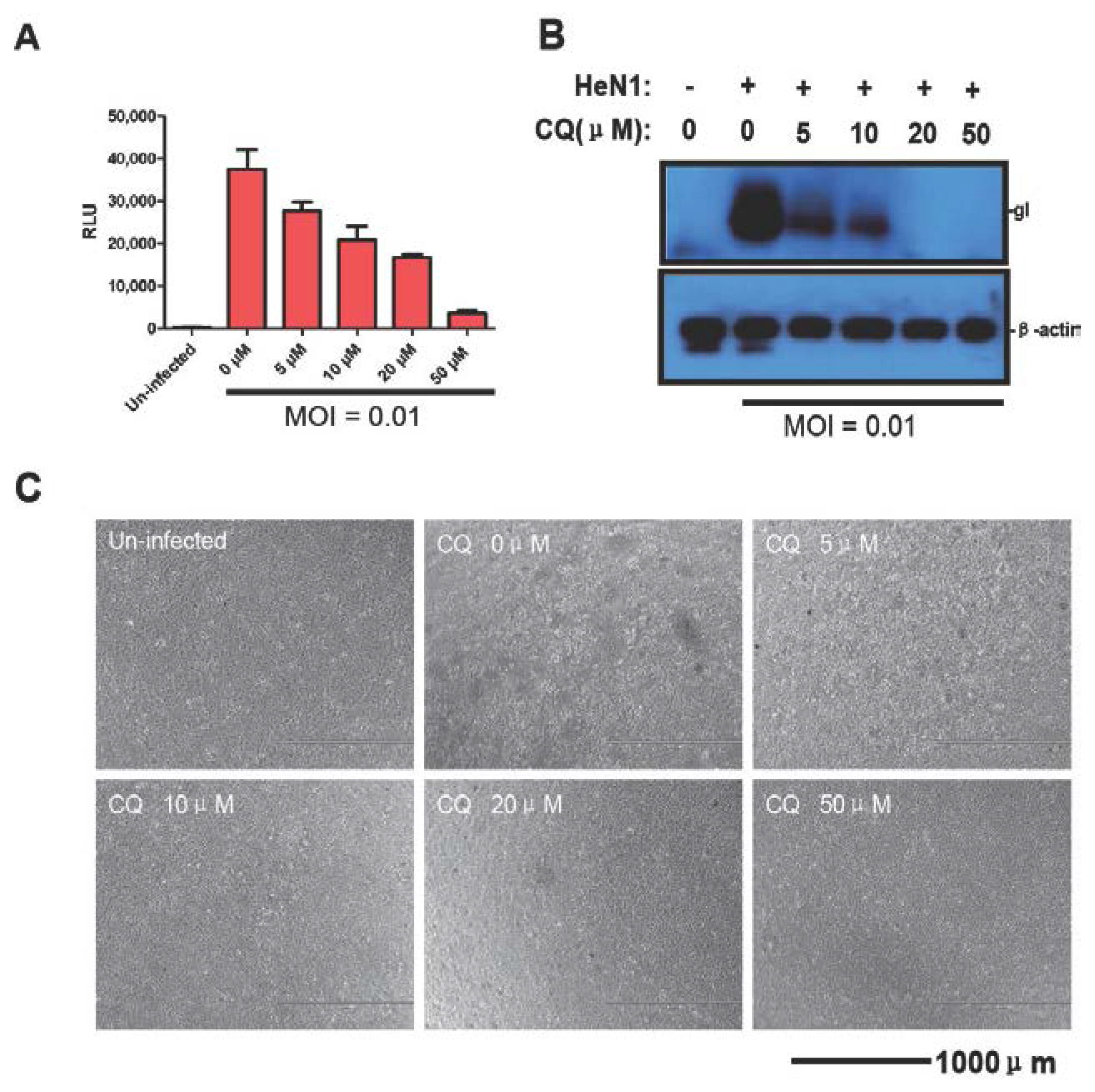
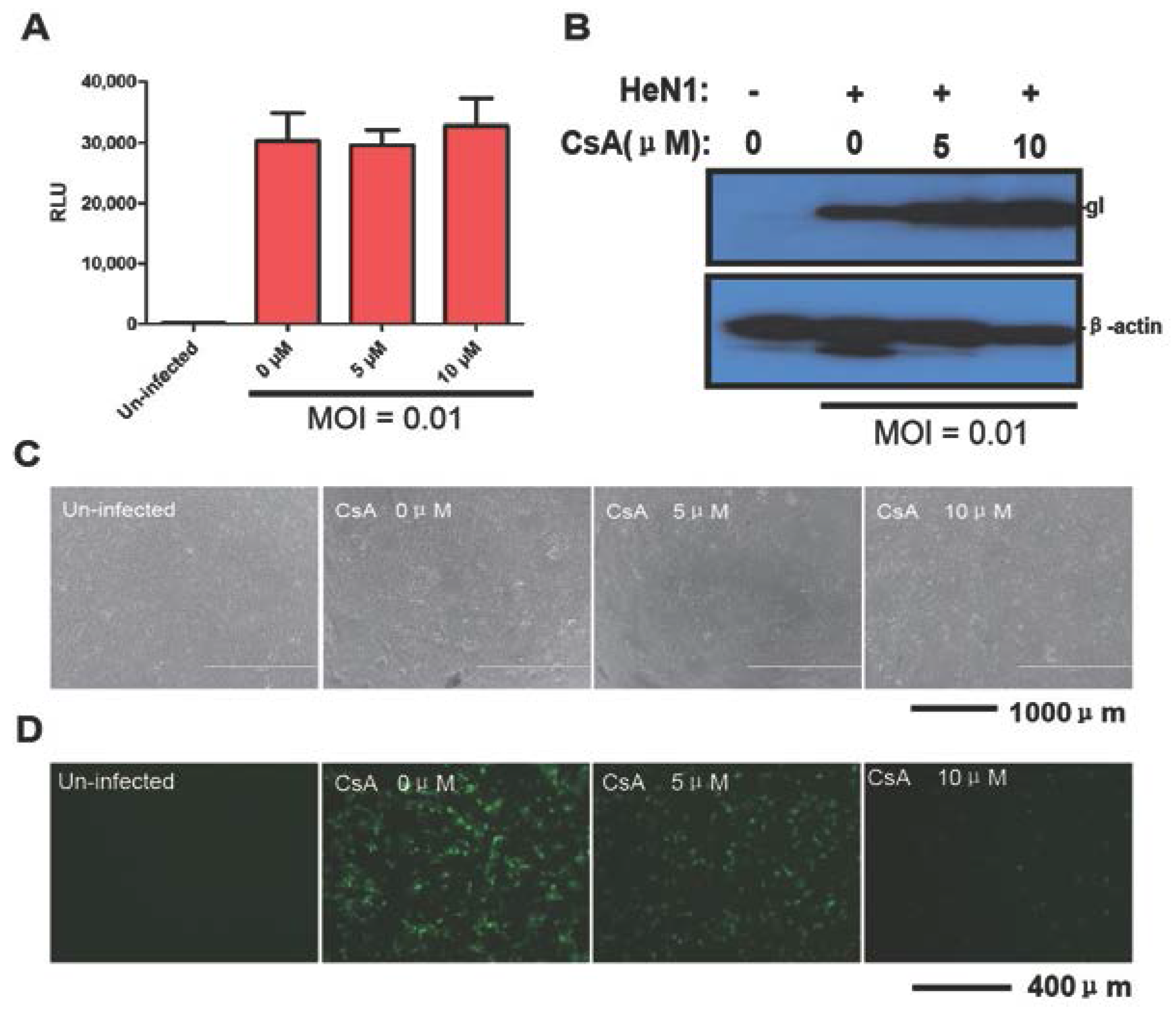
3.3. Evaluating the Antiviral Activity of Chloroquine and CsA
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Muller, T.; Hahn, E.C.; Tottewitz, F.; Kramer, M.; Klupp, B.G.; Mettenleiter, T.C.; Freuling, C. Pseudorabies virus in wild swine: A global perspective. Arch. Virol. 2011, 156, 1691–1705. [Google Scholar] [CrossRef] [PubMed]
- Mettenleiter, T.C. Aujeszky’s disease (pseudorabies) virus: The virus and molecular pathogenesis—State of the art, June 1999. Vet. Res. 2000, 31, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Liu, F.; Zheng, H.; Liang, C.; Zhou, Y.J.; Jiang, Y.F.; Shan, T.L.; Gao, F.; Li, G.X.; Tong, G.Z. Emergence of a Pseudorabies virus variant with increased virulence to piglets. Vet. Microbiol. 2015, 181, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, N.; Cong, X.; Wang, C.H.; Du, M.; Li, L.; Zhao, B.; Yuan, J.; Liu, D.D.; Li, S.; et al. Pathogenicity and genomic characterization of a pseudorabies virus variant isolated from Bartha-K61-vaccinated swine population in China. Vet. Microbiol. 2014, 174, 107–115. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Peng, J.M.; Tian, Z.J.; Zhao, H.Y.; Li, N.; Liu, Y.M.; Chen, J.Z.; Leng, C.L.; Sun, Y.; Chang, D.; et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012. Emerg. Infect. Dis. 2013, 19, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhang, Z.; Lv, L.; Xiao, Y.; Qu, Y.; Ma, H.; Niu, Y.; Wang, G.; Liu, S. Outbreak of variant pseudorabies virus in Bartha-K61-vaccinated piglets in central Shandong Province, China. J. Vet. Diagn. Investig. 2015, 27, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zhang, Q.Z.; Tian, Z.J.; Zheng, H.; Zhao, K.; Liu, F.; Guo, J.C.; Tong, W.; Jiang, C.G.; Wang, S.J.; et al. Genomic characterization of emergent pseudorabies virus in China reveals marked sequence divergence: Evidence for the existence of two major genotypes. Virology 2015, 483, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Yuan, J.; Qin, H.Y.; Luo, Y.; Cong, X.; Li, Y.; Chen, J.; Li, S.; Sun, Y.; Qiu, H.J. A novel gE-deleted pseudorabies virus (PRV) provides rapid and complete protection from lethal challenge with the PRV variant emerging in Bartha-K61-vaccinated swine population in China. Vaccine 2014, 32, 3379–3385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Guo, L.; Jia, X.; Wang, T.; Wang, J.; Sun, Z.; Wang, L.; Li, X.; Tan, F.; Tian, K. Construction of a triple gene-deleted Chinese Pseudorabies virus variant and its efficacy study as a vaccine candidate on suckling piglets. Vaccine 2015, 33, 2432–2437. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, T.; Kohyama, M.; Hirayasu, K.; Arase, H. Engineering large viral DNA genomes using the CRISPR-Cas9 system. Microbiol. Immunol. 2014, 58, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., III. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Wyman, C.; Kanaar, R. DNA double-strand break repair: All’s well that ends well. Annu. Rev. Genet. 2006, 40, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Urnov, F.D.; Rebar, E.J.; Holmes, M.C.; Zhang, H.S.; Gregory, P.D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genet. 2010, 11, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Bogdanove, A.J.; Voytas, D.F. TAL effectors: Customizable proteins for DNA targeting. Science 2011, 333, 1843–1846. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; Zhang, F. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Sun, L.; Gao, D.; Ding, C.; Li, Z.; Li, Y.; Cun, W.; Li, Q. High-efficiency targeted editing of large viral genomes by RNA-guided nucleases. PLoS Pathog. 2014, 10, e1004090. [Google Scholar] [CrossRef] [PubMed]
- Ebina, H.; Misawa, N.; Kanemura, Y.; Koyanagi, Y. Harnessing the CRISPR/Cas9 system to disrupt latent HIV-1 provirus. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.K.; Gu, Y.; Diaz, A.; Marlett, J.; Takahashi, Y.; Li, M.; Suzuki, K.; Xu, R.; Hishida, T.; Chang, C.J.; et al. Use of the CRISPR/Cas9 system as an intracellular defense against HIV-1 infection in human cells. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Qin, C.; Lang, Y.; Wang, M.; Lin, M.; Li, C.; Zhang, R.; Tang, J. A simple and rapid approach to manipulate pseudorabies virus genome by CRISPR/Cas9 system. Biotechnol. Lett. 2015, 37, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Mei, D.Y.; Liu, Q.N.; Qiao, X.H.; Ruan, W.M.; Huang, T.; Cao, G.S. Research of methods to detect genomic mutations induced by CRISPR/Cas systems. J. Biotechnol. 2015, 214, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Blue Heron Biotechnology. Available online: https://wwws.blueheronbio.com/external/tools/gRNASrc.jsp (accessed on 1 September 2015).
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.A.; Enquist, L.W. Construction and transposon mutagenesis in Escherichia coli of a full-length infectious clone of pseudorabies virus, an alphaherpesvirus. J. Virol. 1999, 73, 6405–6014. [Google Scholar] [PubMed]
- Quint, W.; Gielkens, A.; Van Oirschot, J.; Berns, A.; Cuypers, H.T. Construction and characterization of deletion mutants of pseudorabies virus: A new generation of “live” vaccines. J. Gen. Virol. 1987, 68, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef] [PubMed]
- Savarino, A.; Boelaert, J.R.; Cassone, A.; Majori, G.; Cauda, R. Effects of chloroquine on viral infections: An old drug against today's diseases? Lancet Infect. Dis. 2003, 3, 722–727. [Google Scholar] [CrossRef]
- Koyama, A.H.; Uchida, T. Inhibition of multiplication of herpes simplex virus type 1 by ammonium chloride and chloroquine. Virology 1984, 138, 332–335. [Google Scholar] [CrossRef]
- Borel, J.F.; Feurer, C.; Magnee, C.; Stahelin, H. Effects of the new anti-lymphocytic peptide cyclosporin A in animals. Immunology 1977, 32, 1017–1025. [Google Scholar] [PubMed]
- De Wilde, A.H.; Li, Y.; van der Meer, Y.; Vuagniaux, G.; Lysek, R.; Fang, Y.; Snijder, E.J.; van Hemert, M.J. Cyclophilin inhibitors block arterivirus replication by interfering with viral RNA synthesis. J. Virol. 2013, 87, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Sakamoto, N.; Tanabe, Y.; Koyama, T.; Itsui, Y.; Takeda, Y.; Chen, C.H.; Kakinuma, S.; Oooka, S.; Maekawa, S.; et al. Suppression of hepatitis C virus replication by cyclosporin a is mediated by blockade of cyclophilins. Gastroenterology 2005, 129, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.T.; Stein, D.S.; Schnittman, S.M. Primary human immunodeficiency virus type 1 infection: Review of pathogenesis and early treatment intervention in humans and animal retrovirus infections. J. Infect. Dis. 1993, 168, 1490–1501. [Google Scholar] [CrossRef] [PubMed]
- Watashi, K.; Hijikata, M.; Hosaka, M.; Yamaji, M.; Shimotohno, K. Cyclosporin A suppresses replication of hepatitis C virus genome in cultured hepatocytes. Hepatology 2003, 38, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Sakamoto, N.; Enomoto, N.; Tanabe, Y.; Kanazawa, N.; Koyama, T.; Kurosaki, M.; Maekawa, S.; Yamashiro, T.; Chen, C.H.; et al. Specific inhibition of hepatitis C virus replication by cyclosporin A. Biochem. Biophys. Res. Commun. 2004, 313, 42–47. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, R.C.; Epand, R.M.; Johnson, D.C. Cyclosporine A inhibits herpes simplex virus-induced cell fusion but not virus penetration into cells. Virology 1987, 159, 1–9. [Google Scholar] [CrossRef]
- Vahlne, A.; Larsson, P.A.; Horal, P.; Ahlmen, J.; Svennerholm, B.; Gronowitz, J.S.; Olofsson, S. Inhibition of herpes simplex virus production in vitro by cyclosporin A. Arch. Virol. 1992, 122, 61–75. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers and sgRNAs | Sequences |
|---|---|
| CreBamHI | 5′-CTTTTGCAAAAAGCTCCCGGGATCCTGTATATCCATTTTCG-3′ |
| 5′-CGAAAATGGATATACAGGATCCCGGGAGCTTTTTGCAAAAG-3′ | |
| CreNotI | 5′-GGATGATCCTCCAGCGGCCGCATCTCATGCTGGAG-3′ |
| 5′-CTCCAGCATGAGATGCGGCCGCTGGAGGATCATCC-3′ | |
| CreEcoRI XhoI | 5′-CTTATCATGTCTGAATTCCGTCGACCTCTAGCTCGAGCTTGG-3′ |
| 5′-CAAGCTCGAGCTAGAGGTCGACGGAATTCAGACATGATAAG-3′ | |
| Luciferase | 5′-CACGCTAGCCACCATGGAAGATGCCAAAAAC-3′ |
| 5′-AGCTTTGTTTAAAC TTACACGGCGATCTTGCCGC-3′ | |
| HR L arm | 5′-ACAAGATCTCCGGTCCGTAGCCTCCGCAGTA-3′ |
| 5′-ACAACGCGTCGAAGCTCGGCCAACGTCATC-3′ | |
| HR R arm | 5′-CCGGAATTCGGGCCGTGTTCTTTGTGGC-3′ |
| 5′-CGGCTCGAGACTCGCTGGGCGTCTCGTTG-3′ | |
| sgRNA-gE1 | 5′-CACCGGGGCAGGAACGTCCAGATCC-3′ |
| 5′-AAACGGATCTGGACGTTCCTGCCCC-3′ | |
| sgRNA-US3-1 | 5′-CACCGCCCCGACGAGATCCTGTACT-3′ |
| 5′-AAACAGTACAGGATCTCGTCGGGGC-3′ | |
| sgRNA-US3-2 | 5′-CACCGGAGATCATCATCGACGGCGA-3′ |
| 5′-AAACTCGCCGTCGATGATGATCTCC-3′ | |
| sgRNA-US3-3 | 5′-CACCGGAGATCATCATCGACGGCGA-3′ |
| 5′-AAACTCGCCGTCGATGATGATCTCC-3′ | |
| sgRNA-US2-1 | 5′-CACCGACCGTGGTCACGCTGATGGA-3′ |
| 5′-AAACTCCATCAGCGTGACCACGGTC-3′ | |
| sgRNA-US2-2 | 5′-CACCGGGGCGCATCCCCGCCTTCGT-3′ |
| 5′-AAACACGAAGGCGGGGATGCGCCCC-3′ | |
| sgRNA-US2-3 | 5′-CACCGGGCGCACCCGGACCTGTGGA-3′ |
| 5′-AAACTCCACAGGTCCGGGTGCGCCC-3′ | |
| sgRNA-US9-1 | 5′-CACCGCGACGTCCTGCTGGCCCCCA-3′ |
| 5′-AAACTGGGGGCCAGCAGGACGTCGC-3′ | |
| sgRNA-US9-2 | 5′-CACCGGCCAGCAGGACGTCGGCGGC-3′ |
| 5′-AAACGCCGCCGACGTCCTGCTGGCC-3′ | |
| sgRNA-US9-3 | 5′-CACCGGGGGTCCCTTGGGGGCCAGC-3′ |
| 5′-AAACGCTGGCCCCCAAGGGACCCCC-3′ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.-D.; Liu, J.-T.; Fang, Q.-Q.; Wang, T.-Y.; Sun, M.-X.; An, T.-Q.; Tian, Z.-J.; Cai, X.-H. Recombinant Pseudorabies Virus (PRV) Expressing Firefly Luciferase Effectively Screened for CRISPR/Cas9 Single Guide RNAs and Antiviral Compounds. Viruses 2016, 8, 90. https://0-doi-org.brum.beds.ac.uk/10.3390/v8040090
Tang Y-D, Liu J-T, Fang Q-Q, Wang T-Y, Sun M-X, An T-Q, Tian Z-J, Cai X-H. Recombinant Pseudorabies Virus (PRV) Expressing Firefly Luciferase Effectively Screened for CRISPR/Cas9 Single Guide RNAs and Antiviral Compounds. Viruses. 2016; 8(4):90. https://0-doi-org.brum.beds.ac.uk/10.3390/v8040090
Chicago/Turabian StyleTang, Yan-Dong, Ji-Ting Liu, Qiong-Qiong Fang, Tong-Yun Wang, Ming-Xia Sun, Tong-Qing An, Zhi-Jun Tian, and Xue-Hui Cai. 2016. "Recombinant Pseudorabies Virus (PRV) Expressing Firefly Luciferase Effectively Screened for CRISPR/Cas9 Single Guide RNAs and Antiviral Compounds" Viruses 8, no. 4: 90. https://0-doi-org.brum.beds.ac.uk/10.3390/v8040090