1. Introduction
Chlamydia trachomatis is the most common cause of curable bacterial sexually transmitted infection worldwide because of their prevalence and potentially devastating reproductive consequences, including pelvic inflammatory disease, infertility, and ectopic pregnancy. The number of chlamydial-infected citizens reached 1,422,976 in the United States in 2012 [
1]. In China, the prevalence of chlamydial infection has also increased, and was reported to be 2.6 and 2.1 per 100 individuals for women and men, respectively [
2]. Although the basic biology of the pathogen is well established, the pathogenic mechanisms are still unclear. Even worse, the condition of antibiotics resistance is becoming more and more serious. Therefore, further exploration of the
Chlamydia infection mechanisms is sorely needed to develop new treatment methods to overcome the current public health challenges. Given the condition of a blocked process of developing a vaccine for
Chlamydia trachomatis, use of a specific chlamydiaphage may be a new solution.
Chlamydiae have an obligate intracellular developmental cycle that alternates between the infectious elementary body and the replicative reticulate body. Six bacteriophages have been isolated from the Chlamydiae: Chp1, Chp2, Chp3, Chp4, φCPG1, and φCPAR39. All of the chlamydiaphages share similar features; they are small, icosahedral T = 1 particles containing circular, single-stranded DNA genomes, and molecular characterization revealed that they belong to the virus family Microviridae [
3,
4,
5]. φCPG1 is a lytic phage specific to
Chlamydia caviae, which is a natural parasite of the guinea pig. The genome of φCPG1 contains five open reading frames, which encode the capsid proteins Vp1, Vp2, and Vp3. Capsid proteins, especially Vp1, play essential roles in adhesion and invasion. Our research group has compared the Vp1 of all
Chlamydia phages found until now and recovered nearly 98% comparability among them. Even though a specific bacteriophage of
C. trachomatis has not yet been detected, the high homology among the known chlamydiaphages allows for the reasonable assumption of cross-reaction among species. The phage host range is specified by a peptidic loop of Vp1, and sequence variation within the loop is consistent with the observed overlapping host ranges of the phages [
6]. Vp1 also plays an important role in the interaction between
Chlamydia and its host cell. Our research group successfully expressed and purified Vp1, which was incubated with
C. trachomatis and confirmed to inhibit its growth [
7]. However, the specific mechanism underlying this inhibition effect has not yet been explored.
The mechanisms of
C. trachomatis infection remain mysterious. Most research conducted in this field thus far has considered the inflammation-induced pathological damage [
8,
9]. The
C. trachomatis-induced inflammation reaction is mediated by signaling pathways in the host cell that are triggered after infection, including the mitogen-activated protein kinase (MAPK) pathway, Janus-activated kinase-signal transducer and activator of transcription (JAK/STAT) pathway, and nuclear factor-kappa B (NF-κB) pathway, among others. In particular, the MAPK pathway plays a major role in post-infection signal transduction, and the growth of
C. trachomatis depends on the MAPK/ERK (mitogen-activated protein kinase / extracellular regulated protein kinases) pathway. During the course of incubation in the host,
C. trachomatis enters into its own growth cycle, which induces increased phosphorylation levels of ERK protein in the host cell. Once the growth cycle is blocked with U0126, a specific inhibitor of MEK, the infection rate decreases, the major outer membrane protein (MOMP) level of
C. trachomatis is reduced, and the phosphorylation level of ERK1/2 of the host is downregulated [
10]. These facts reveal that
C. trachomatis modulates the host MEK/ERK pathway and relies on it for its growth. In response to the increase in p-ERK, the host cell will release pro-inflammatory factors such as interleukin (IL)-1, IL-8, and tumor necrosis factor (TNF)-α [
11]. IL-1 and IL-8 are considered the most significant factors contributing to host cell inflammatory injury in
C. trachomatis infection [
12,
13].
Azithromycin has been the recommended antibiotic for treatment of
C. trachomatis infection according to the guideline. Previous studies reported that azithromycin inhibits cytokine production, which may contribute to its therapeutic effect in the treatment of chronic
C. trachomatis infection [
14]. In addition, phosphorylation of ERK was found to be inhibited after azithromycin treatment in epithelial cells obtained from women with recurrent infection [
15]. Traditionally, bacteriolysis is considered the main mechanism by which a bacteriophage inhibits the growth of its host. However, there may be alternative mechanisms that have not yet been explored, which could provide useful insights for the development of new treatments. In the present study, we aimed to address the following remaining questions related to the mechanism of
Chlamydia growth inhibition by Vp1. In particular, we investigated whether Vp1 might exert its inhibition effect via the MAPK pathway similar to azithromycin, and compared the inhibition effect of Vp1 and azithromycin.
2. Materials and Methods
2.1. McCoy Cell Culture
McCoy cells (mouse fibroblast L cell origin) were purchased from the Chinese Academy of Medical Sciences, and stored in the Tianjin Institute of Sexually Transmitted Diseases, China. They were grown in minimal essential medium supplemented with 10% fetal bovine serum for Chlamydia culture. When the cells were distributed compactly and uniformly as a single layer, they were transferred to 6-well plates for continuous incubation and Chlamydia infection.
2.2. C. trachomatis Infection
The monolayer McCoy cells were pretreated with 30 μg/mL of DEAE-D for 30 min to increase the susceptibility of infection. The C. trachomatis strain used in this study was the E serotype strain that is maintained in our laboratory. The strain was pretreated with two freeze-thawing cycles, oscillation, and centrifugation at 500× g for 5 min. The plates were further centrifuged at 500× g at 32 °C for 1 h to facilitate Chlamydia adhesion. After 2 h, the wells of the plate were overlaid with culture medium containing 1 mg/L cycloheximide, and incubated for 44–48 h. The majority of cells were collected in transport culture medium, and the remaining cells were fixed with methanol and stained with iodine dye to observe the residual inclusions to verify that the infective rate was over 90%.
2.3. Vp1 Expression, Identification, and Purification
The stored
E. coli bacteria that contained Vp1-pET30a(+) [
16] were incubated in kanamycin-resistant Luria-Bertani medium for 12–16 h. The bacteria were amplified in the shaker and Vp1 expression was induced with 0.03 mM isopropyl-β-
d-thiogalactopyranoside (IPTG) at 30 °C for 3 h until reaching the logarithmic phase (OD 0.6–0.8 in 600 nm). The bacterial solution was centrifuged and the sediment was treated with 4 mg/mL lysozyme and 3% Triton-X100. The suspension was ultrasonicated and centrifuged. The sediment was blended in PBS with 6 mol/L urea and identified by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The purification of Vp1 was achieved with the PAGE recovery method, renaturated by dialysis and quantified for further use. LPS (Lipopolysaccharides) was neutralized less than 0.1 EU/mL by ToxinEraser
TM endotoxin removal resin kit from GenScript (Piscataway, NJ, USA). Vp1 was confirmed without toxic effect on McCoy cells by MTT (3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide) method.
2.4. Treatments and Experimental Design
The McCoy cells were incubated with or without
C. trachomatis, Vp1 and azithromycin, and classified into 6 experimental groups (
Table 1).
C. trachomatis was infected at 3 × 10
5 IFU/mL after purification and quantification. The inoculation loads of Vp1 and azithromycin were 60 μg/mL and 25 μg/mL, according to previous reports
12 and the results of preliminary experiments. After 48 h, the inclusions were counted under the microscope and stained with iodine dye.
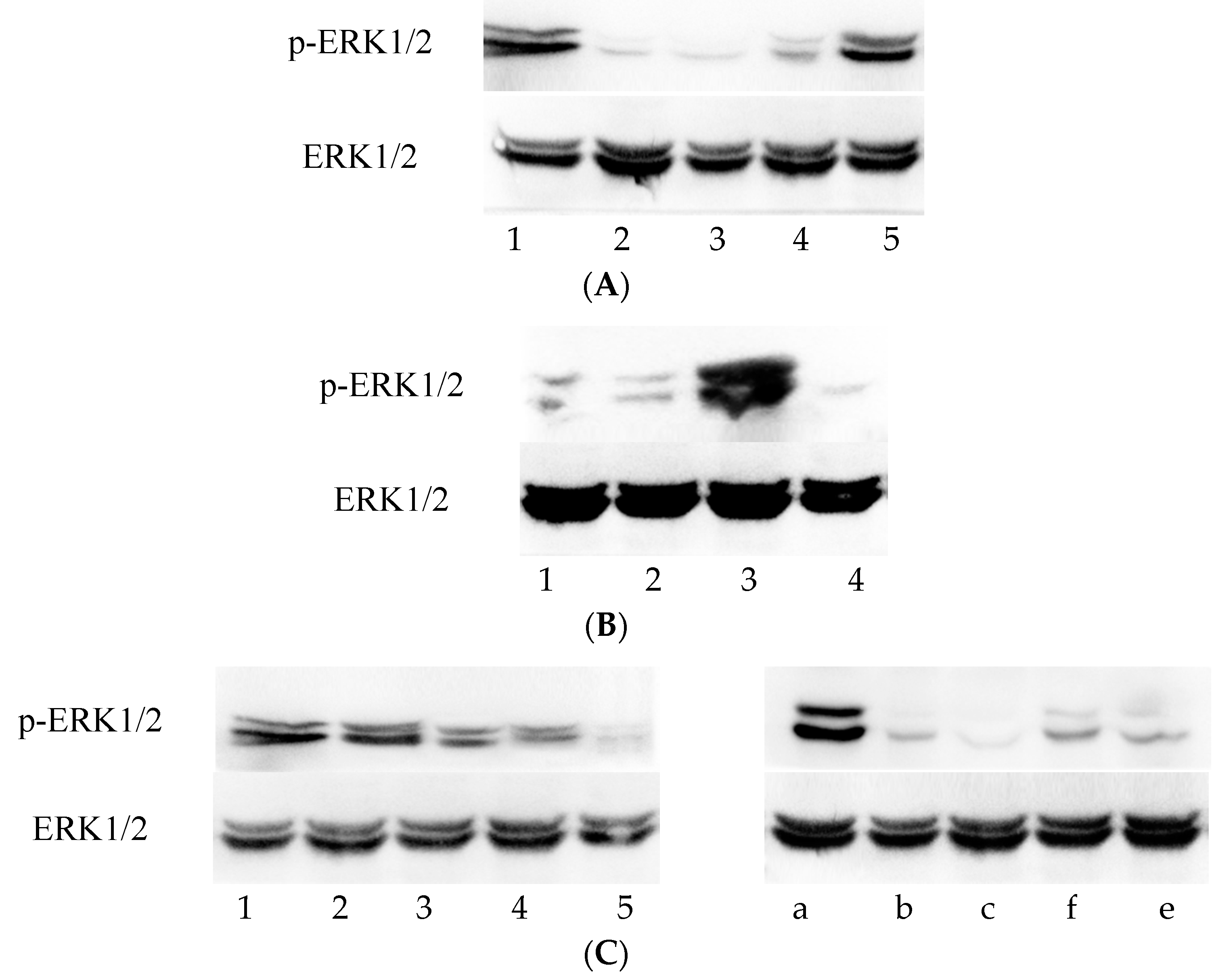
2.5. Western Blotting
The cells were incubated at 0, 12 h, 24 h, 36 h, or 48 h, according to treatment, and the cellular proteins was extracted by the RIPA (Radio Immunoprecipitation Assay) method. The total proteins were separated by SDS-PAGE. The polyvinylidene fluoride (PVDF) membrane (Millipore Immobilon-P, Darmstadt, Germany). The monoclonal antibodies of p-ERK1/2 and total ERK1/2 (Cell Signaling Technology, Danvers, MA, USA) were added to the relevant treatment groups at a dilution of 1:1000. The corresponding secondary antibodies (Cell Signaling Technology) were added at dilutions of 1:5000–1:10000. The PVDF membrane was exposed and photographed after development, and the grey value ratio was used for evaluation determined with Gel-Pro Analyzer Software (Media Cybernetics, Rockville, MD, USA). The p-ERK1/2 level was standardized by the grey value ratio of ERK1/2. The experiment was repeated three times and the results are presented as the mean and standard deviation.
2.6. Real-Time PCR
The PCR protocols were performed according to the methods described by Wang and Seed [
17]. In brief, cells in Groups M, C, C + V, and C + A were incubated for 48 h in 6-well plates, and the cells were dissociated and centrifuged at 500× g for collection according to the treatment groups. Total RNA was extracted by the TRNzol method and its purity and concentration were verified on a spectrophotometer. Reverse transcription was carried out with the PrimeScript RT reagent Kit (Qiagen, Hilden, Germany) with gDNA Eraser (Qiagen) for RT-PCR. The primers were synthesized by Invitrogen (Carlsbad, CA, USA) (
Table 2). The PCR program was carried out at 95 °C 30 s, (95 °C 5 s, 60 °C 40 s) × 45 with SYBR® Premix Ex Taq™ II (Tli RNaseH Plus) (Takara Bio, Dalian, China), ROX plus (Takara Bio) on an ABI 7500 system (Applied Biosystems, Foster City, CA, USA). The results were quantitatively analyzed based on
Actin by the 2
−ΔΔct method.
2.7. IL-1 and IL-8 Enzyme-Linked Immunosorbent Assay
The supernatants of treated cells from the different groups were collected at various time points of incubation (0, 12 h, 24 h, 36 h, 48 h) and subjected to enzyme-linked immunosorbent assay for detection of cytokine (IL-1 and IL-8) production levels. The experiment was repeated three times and the results are presented as the mean with standard deviation.
2.8. Data Analysis
All of the data were analyzed with SPSS 17.0 software (IBM, Armonk, NY, USA) according to the statistics. The difference was considered statistically significant if the p-value was less than 0.05.
4. Discussion
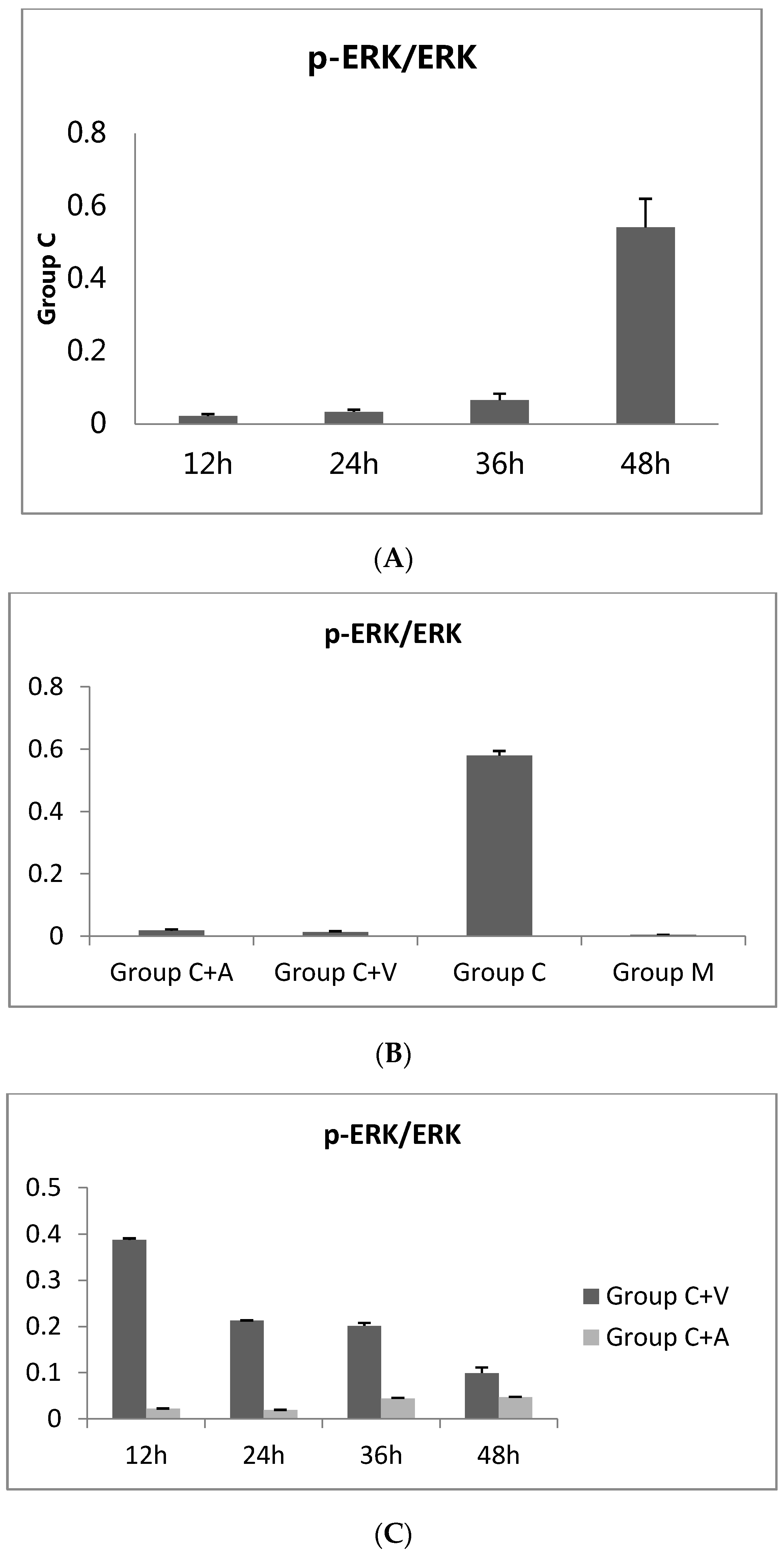
C. trachomatis urogenital tract infection is a sexually transmitted disease that has become increasingly common and is a global health concern. The mechanisms of Chlamydia infection are closely related to inflammation-induced pathologic damage. Accordingly, the MAPK signaling pathways play a major role in the C. trachomatis infection, especially with respect to the growth and production of pro-inflammatory cytokines. To further understand the mechanisms of Chlamydia infection, it is important to investigate the mechanisms underlying the inhibition of C. trachomatis by the bacteriophage capsid protein Vp1. We here report the first attempt at exploring the relationship between C. trachomatis-infected cells treated with Vp1 and the MEK/ERK pathway at the protein and gene levels. Furthermore, we evaluated the effects of Vp1 treatment on the pro-inflammatory cytokines IL-8 and IL-1 to clarify the mechanism of inhibition.
First, we found that the p-ERK level increased gradually during
C. trachomatis incubation with McCoy cells, which confirms Du
et al. [
8] in HeLa cells. In addition, Vp1 decreased the p-ERK level and the
ERK1 and
ERK2 mRNA expression levels, showing the same effects as azithromycin treatment. These results demonstrate that Vp1 and azithromycin have the same mechanism of
Chlamydia inhibition by modulating the MEK/ERK pathway and minimizing ERK activation. The only difference was the effective time of the inhibition: the Vp1 is effective in the late infection period, while azithromycin was effective in the early period of
Chlamydia growth.
Second, the
ERK1 mRNA level was upregulated, while the
ERK2 mRNA level was downregulated after
C. trachomatis infection. Although the
ERK1 and
ERK2 genes have 90% sequence identity and share the same target upstream and downstream, they have some different biological functions. For example, ERK2 plays a key role in hepatocyte cell division, while ERK1 improves long-term hepatocyte survival [
18]. In addition, specifically ERK2 but not ERK1 is required for the c-Met-paxillin signaling axis in HGF (Hepatocyte Growth Factor)-mediated motility [
14]. The results of this study suggest that ERK1 might play a more important role in
C. trachomatis growth than ERK2. However, confirmation of this hypothesis and the detailed mechanism await further investigation.
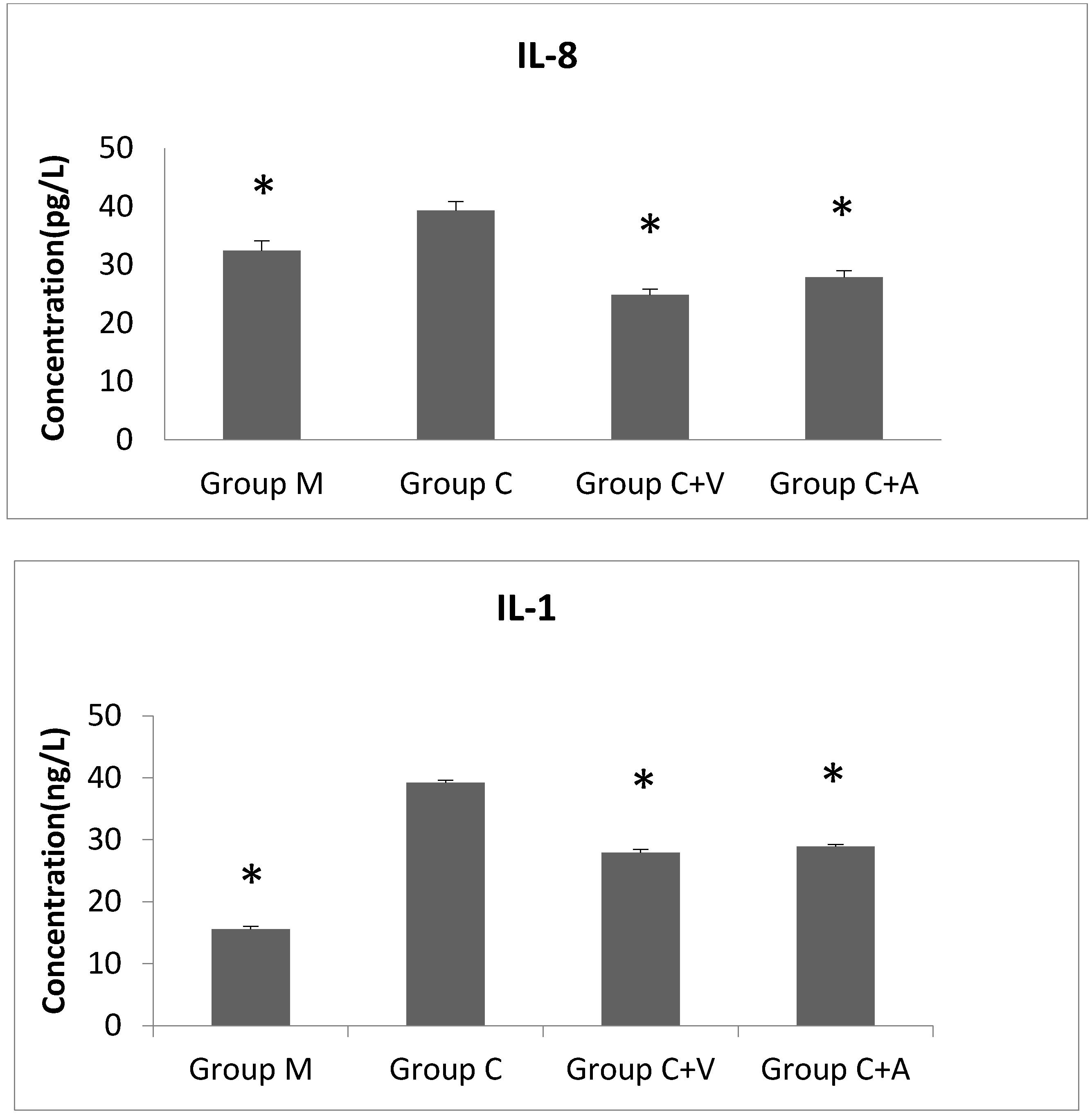
Finally, the pro-inflammatory cytokines IL-8 and IL-1 were found to increase after
C. trachomatis infection, confirming previous research [
11]. These two cytokines are directly induced by ERK pathways. In our study, production of these two cytokines also decreased after infected cells were treated with Vp1 and azithromycin compared with the untreated group. The IL-8 level of the Vp1-treated group decreased earlier than that of the azithromycin-treated group, while the IL-1 of the both groups showed an initial decrease (similar to the untreated infected cells), and then peaked at 24 h and 36 h before decreasing subsequently. The observed variations of IL-8 and IL-1 provide further understanding of the function of related pathways and their relationship to
Chlamydia inhibition.
5. Conclusions
In conclusion, our research demonstrates that the mechanism of chlamydiaphage φCPG1 capsid protein Vp1 inhibiting C. trachomatis growth is associated with the MAPK pathway, via inhibition of the pro-inflammatory cytokines IL-8 and IL-1. Therefore, we have identified that bacteriophages may also influence their hosts via signaling transduction mediation and not only via bacteriolysis. Thus, we have uncovered a likely preliminary mechanism for how Vp1 inhibits C. trachomatis in cells. Nevertheless, the specific mechanisms determining the relationship between Vp1 and C. trachomatis are still under exploration.
Although azithromycin is still the first-choice recommended treatment for
C. trachomatis [
19], its clinical use has become less frequent because of acquired drug resistance. Although a chlamydia vaccine has been under exploration for several years and several candidate antigens have been screened, vaccine development remains a great challenge [
20]. Therefore, the exploration of infection mechanisms offers new perspective for
Chlamydia treatment. Bacteriophages have unique characteristics and show good potential as a clinical treatment to overcome antibiotic resistance [
21]. The use of bacteriophages would minimize the chance of secondary infections because of their specific combination with hosts, which is a continued risk with antibiotics. In addition to their high specificity, the concentration at the site of infection can be assured so that side effects can be limited. Experts make efforts on phage transformation to get broad-spectrum, long-life and multi-function phages [
22]. Therefore, the application of chlamydiaphages as a biotic therapy shows good future promise and warrants further exploration.







{kind=link}
{kind=link}
{kind=link}
{kind=link}