
Cross- and Co-Packaging of Retroviral RNAs and Their Consequences



Abstract
:1. Introduction
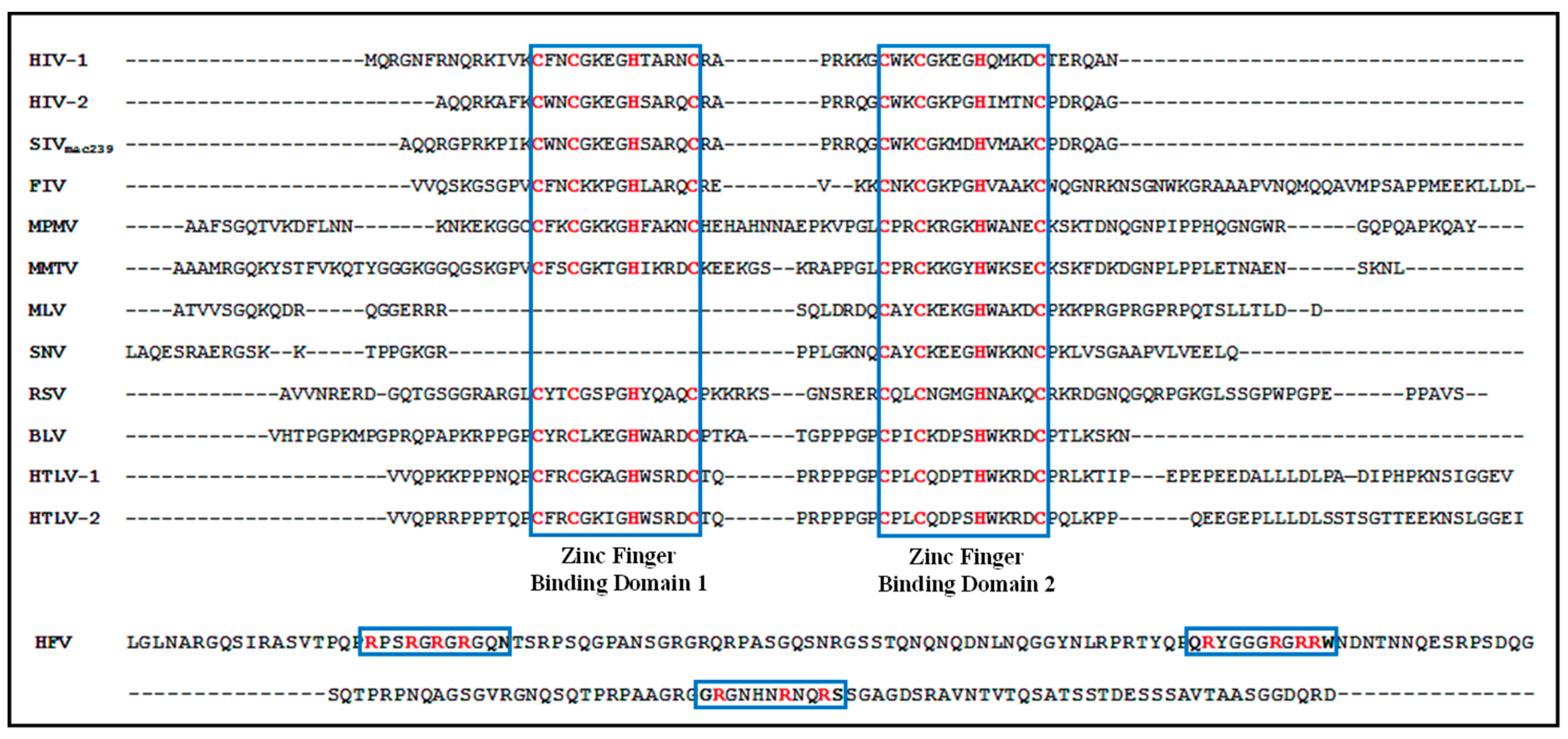
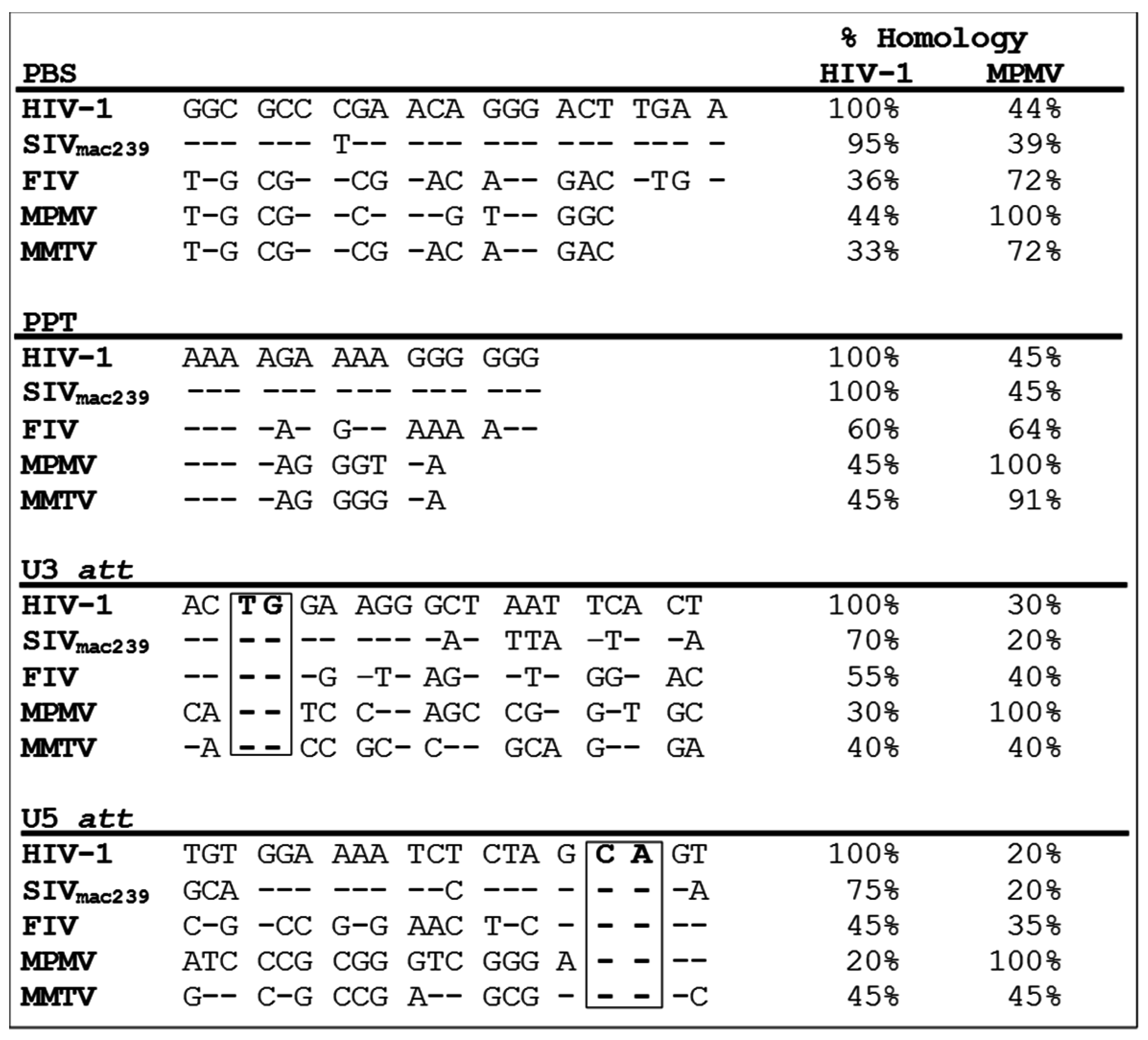
2. How Similar Is the Structural Organization of Packaging Elements within Various Retroviruses?
3. What Determines the Specificity of gRNA Packaging among Retroviruses?
4. How Do Retroviruses Cross-Package Each Other’s Genomes?
5. What Determines the Propagation Capabilities of the Cross-Packaged RNAs?
6. Packaging of Non-Viral RNAs into Retroviral Particles
7. Packaging of Cellular RNAs into Retroviral Particles
8. Co-packaging and Consequences of Co-Packaging among Diverse Retroviruses
8.1. Role of Dimerization in Co-Packaging
8.2. Recombination Potential of Retroviruses
9. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rulli, S.J.; Hibbert, C.S.; Mirro, J.; Pederson, T.; Biswal, S.; Rein, A. Selective and Nonselective Packaging of Cellular RNAs in Retrovirus Particles. J. Virol. 2007, 81, 6623–6631. [Google Scholar] [CrossRef] [PubMed]
- Coffin, J.M.; Hughes, S.H.; Varmus, H.E. (Eds.) Retroviruses; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997.
- Johnson, S.F.; Telesnitsky, A. Retroviral RNA Dimerization and Packaging: The What, How, When, Where, and Why. PLoS Pathog. 2010, 6, e1001007. [Google Scholar] [CrossRef] [PubMed]
- Hussein, I.T.M.; Ni, N.; Galli, A.; Chen, J.; Moore, M.D.; Hu, W.-S. Delineation of the preferences and requirements of the human immunodeficiency virus type 1 dimerization initiation signal by using an in vivo cell-based selection approach. J. Virol. 2010, 84, 6866–6875. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, V.; Summers, M.F. How retroviruses select their genomes. Nat. Rev. Microbiol. 2005, 3, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Lever, A.M.L. HIV-1 RNA packaging. Adv. Pharmacol. 2007, 55, 1–32. [Google Scholar] [PubMed]
- Moore, M.D.; Nikolaitchik, O.A.; Chen, J.; Hammarskjöld, M.-L.; Rekosh, D.; Hu, W.-S. Probing the HIV-1 Genomic RNA Trafficking Pathway and Dimerization by Genetic Recombination and Single Virion Analyses. PLoS Pathog. 2009, 5, e1000627. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Miyake, A.; Nomaguchi, M.; Adachi, A. Structural dynamics of retroviral genome and the packaging. Front. Microbiol. 2011, 2, 264. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.S.; Liang, C.; Wainberg, M.A. Is HIV-1 RNA dimerization a prerequisite for packaging? Yes, no, probably? Retrovirology 2004, 1, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, K.; Heng, X.; Summers, M.F. Structural determinants and mechanism of HIV-1 genome packaging. J. Mol. Biol. 2011, 410, 609–633. [Google Scholar] [CrossRef] [PubMed]
- Hibbert, C.S.; Mirro, J.; Rein, A. mRNA molecules containing murine leukemia virus packaging signals are encapsidated as dimers. J. Virol. 2004, 78, 10927–10938. [Google Scholar] [CrossRef] [PubMed]
- Paillart, J.-C.; Dettenhofer, M.; Yu, X.-F.; Ehresmann, C.; Ehresmann, B.; Marquet, R. First snapshots of the HIV-1 RNA structure in infected cells and in virions. J. Biol. Chem. 2004, 279, 48397–48403. [Google Scholar] [CrossRef] [PubMed]
- Baig, T.T.; Lanchy, J.-M.; Lodmell, J.S. HIV-2 RNA dimerization is regulated by intramolecular interactions in vitro. RNA 2007, 13, 1341–1354. [Google Scholar] [CrossRef] [PubMed]
- Berkhout, B.; van Wamel, J.L. Role of the DIS hairpin in replication of human immunodeficiency virus type 1. J. Virol. 1996, 70, 6723–6732. [Google Scholar] [PubMed]
- Lanchy, J.-M.; Ivanovitch, J.D.; Lodmell, J.S. A structural linkage between the dimerization and encapsidation signals in HIV-2 leader RNA. RNA 2003, 9, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Lanchy, J.-M.; Lodmell, J.S. An extended stem-loop 1 is necessary for human immunodeficiency virus type 2 replication and affects genomic RNA encapsidation. J. Virol. 2007, 81, 3285–3292. [Google Scholar] [CrossRef] [PubMed]
- Laughrea, M.; Jetté, L. HIV-1 Genome Dimerization: Kissing-Loop Hairpin Dictates Whether Nucleotides Downstream of the 5‘ Splice Junction Contribute to Loose and Tight Dimerization of Human Immunodeficiency Virus RNA. Biochemistry 1997, 36, 9501–9508. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.S.; Panganiban, A.T. Position dependence of functional hairpins important for human immunodeficiency virus type 1 RNA encapsidation in vivo. J. Virol. 1997, 71, 2050–2058. [Google Scholar] [PubMed]
- Paillart, J.C.; Skripkin, E.; Ehresmann, B.; Ehresmann, C.; Marquet, R. A loop-loop “kissing” complex is the essential part of the dimer linkage of genomic HIV-1 RNA. Proc. Natl. Acad. Sci. USA 1996, 93, 5572–5577. [Google Scholar] [CrossRef] [PubMed]
- Paillart, J.; Marquet, R.; Skripkin, E.; Ehresmann, C.; Ehresmann, B. Dimerization of retroviral genomic RNAs: structural and functional implications. Biochimie 1996, 78, 639–653. [Google Scholar] [CrossRef]
- Berkowitz, R.; Fisher, J.; Goff, S.P. RNA packaging. Curr. Top. Microbiol. Immunol. 1996, 214, 177–218. [Google Scholar]
- Cain, D.; Erlwein, O.; Grigg, A.; Russell, R.A.; McClure, M.O. Palindromic sequence plays a critical role in human foamy virus dimerization. J. Virol. 2001, 75, 3731–3739. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Heng, X.; Garyu, L.; Monti, S.; Garcia, E.L.; Kharytonchyk, S.; Dorjsuren, B.; Kulandaivel, G.; Jones, S.; Hiremath, A.; et al. NMR detection of structures in the HIV-1 5’-leader RNA that regulate genome packaging. Science 2011, 334, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Al Shamsi, I.R.; Al Dhaheri, N.S.; Phillip, P.S.; Mustafa, F.; Rizvi, T.A. Reciprocal cross-packaging of primate lentiviral (HIV-1 and SIV) RNAs by heterologous non-lentiviral MPMV proteins. Virus Res. 2011, 155, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Al Dhaheri, N.S.; Phillip, P.S.; Ghazawi, A.; Ali, J.; Beebi, E.; Jaballah, S.A.; Rizvi, T.A. Cross-packaging of genetically distinct mouse and primate retroviral RNAs. Retrovirology 2009, 6, 66. [Google Scholar] [CrossRef] [PubMed]
- Motomura, K.; Chen, J.; Hu, W.-S. Genetic Recombination between Human Immunodeficiency Virus Type 1 (HIV-1) and HIV-2, Two Distinct Human Lentiviruses. J. Virol. 2008, 82, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.D.; Fu, W.; Nikolaitchik, O.; Chen, J.; Ptak, R.G.; Hu, W.-S. Dimer Initiation Signal of Human Immunodeficiency Virus Type 1: Its Role in Partner Selection during RNA Copackaging and Its Effects on Recombination. J. Virol. 2007, 81, 4002–4011. [Google Scholar] [CrossRef] [PubMed]
- Parveen, Z.; Mukhtar, M.; Goodrich, A.; Acheampong, E.; Dornburg, R.; Pomerantz, R.J. Cross-Packaging of Human Immunodeficiency Virus Type 1 Vector RNA by Spleen Necrosis Virus Proteins: Construction of a New Generation of Spleen Necrosis Virus-Derived Retroviral Vectors. J. Virol. 2004, 78, 6480–6488. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, T.A.; Panganiban, A.T. Simian immunodeficiency virus RNA is efficiently encapsidated by human immunodeficiency virus type 1 particles. J. Virol. 1993, 67, 2681–2688. [Google Scholar] [PubMed]
- White, S.M.; Renda, M.; Nam, N.-Y.; Klimatcheva, E.; Zhu, Y.; Fisk, J.; Halterman, M.; Rimel, B.J.; Federoff, H.; Pandya, S.; et al. Lentivirus Vectors Using Human and Simian Immunodeficiency Virus Elements. J. Virol. 1999, 73, 2832–2840. [Google Scholar] [PubMed]
- Yin, P.D.; Hu, W.S. RNAs from genetically distinct retroviruses can copackage and exchange genetic information in vivo. J. Virol. 1997, 71, 6237–6242. [Google Scholar] [PubMed]
- Jewell, N.A.; Mansky, L.M. In the beginning: genome recognition, RNA encapsidation and the initiation of complex retrovirus assembly. J. Gen. Virol. 2000, 81, 1889–1899. [Google Scholar] [CrossRef] [PubMed]
- Jouvenet, N.; Lainé, S.; Pessel-Vivares, L.; Mougel, M. Cell biology of retroviral RNA packaging. RNA Biol. 2011, 8, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Erlwein, O.; Bieniasz, P.D.; McClure, M.O. Sequences in pol are required for transfer of human foamy virus-based vectors. J. Virol. 1998, 72, 5510–5516. [Google Scholar] [PubMed]
- Heinkelein, M.; Schmidt, M.; Fischer, N.; Moebes, A.; Lindemann, D.; Enssle, J.; Rethwilm, A. Characterization of a cis-acting sequence in the Pol region required to transfer human foamy virus vectors. J. Virol. 1998, 72, 6307–6314. [Google Scholar] [PubMed]
- Wu, M.; Chari, S.; Yanchis, T.; Mergia, A. cis-Acting sequences required for simian foamy virus type 1 vectors. J. Virol. 1998, 72, 3451–3454. [Google Scholar] [PubMed]
- Sorge, J.; Ricci, W.; Hughes, S.H. cis-Acting RNA packaging locus in the 115-nucleotide direct repeat of Rous sarcoma virus. J. Virol. 1983, 48, 667–675. [Google Scholar] [PubMed]
- Aschoff, J.M.; Foster, D.; Coffin, J.M. Point mutations in the avian sarcoma/leukosis virus 3’ untranslated region result in a packaging defect. J. Virol. 1999, 73, 7421–7429. [Google Scholar] [PubMed]
- Yu, S.S.; Kim, J.M.; Kim, S. The 17 nucleotides downstream from the env gene stop codon are important for murine leukemia virus packaging. J. Virol. 2000, 74, 8775–8780. [Google Scholar] [PubMed]
- Kaye, J.F.; Lever, A.M.L. Nonreciprocal Packaging of Human Immunodeficiency Virus Type 1 and Type 2 RNA: a Possible Role for the p2 Domain of Gag in RNA Encapsidation. J. Virol. 1998, 72, 5877–5885. [Google Scholar] [PubMed]
- Damgaard, C.K.; Dyhr-Mikkelsen, H.; Kjems, J. Mapping the RNA binding sites for human immunodeficiency virus type-1 gag and NC proteins within the complete HIV-1 and -2 untranslated leader regions. Nucleic Acids Res. 1998, 26, 3667–3676. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.D.; Allen, J.F.; Lever, A.M. The major human immunodeficiency virus type 2 (HIV-2) packaging signal is present on all HIV-2 RNA species: cotranslational RNA encapsidation and limitation of Gag protein confer specificity. J. Virol. 2001, 75, 12058–12069. [Google Scholar] [CrossRef] [PubMed]
- Baig, T.T.; Lanchy, J.-M.; Lodmell, J.S. Randomization and in vivo selection reveal a GGRG motif essential for packaging human immunodeficiency virus type 2 RNA. J. Virol. 2009, 83, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Wang, S.-W.; Izmailova, E.; Aldovini, A. The simian immunodeficiency virus 5’ untranslated leader sequence plays a role in intracellular viral protein accumulation and in RNA packaging. J. Virol. 2003, 77, 6284–6292. [Google Scholar] [CrossRef] [PubMed]
- Whitney, J.B.; Wainberg, M.A. Impaired RNA incorporation and dimerization in live attenuated leader-variants of SIVmac239. Retrovirology 2006, 3, 96. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, F.; Ghazawi, A.; Jayanth, P.; Phillip, P.S.; Ali, J.; Rizvi, T.A. Sequences intervening between the core packaging determinants are dispensable for maintaining the packaging potential and propagation of feline immunodeficiency virus transfer vector RNAs. J. Virol. 2005, 79, 13817–13821. [Google Scholar] [CrossRef] [PubMed]
- Jaballah, S.A.; Aktar, S.J.; Ali, J.; Phillip, P.S.; Al Dhaheri, N.S.; Jabeen, A.; Rizvi, T.A. A G–C-Rich Palindromic Structural Motif and a Stretch of Single-Stranded Purines Are Required for Optimal Packaging of Mason–Pfizer Monkey Virus (MPMV) Genomic RNA. J. Mol. Biol. 2010, 401, 996–1014. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.D.; Mustafa, F.; Lew, K.A.; Browning, M.T.; Rizvi, T.A. Sequences within both the 5’ untranslated region and the gag gene are important for efficient encapsidation of Mason-Pfizer monkey virus RNA. Virology 2003, 309, 166–178. [Google Scholar] [CrossRef]
- Mustafa, F.; Al Amri, D.; Al Ali, F.; Al Sari, N.; Al Suwaidi, S.; Jayanth, P.; Philips, P.S.; Rizvi, T.A. Sequences within both the 5’ UTR and Gag are required for optimal in vivo packaging and propagation of mouse mammary tumor virus (MMTV) genomic RNA. PLoS ONE 2012, 7, e47088. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M.; Krueger, A.E.; Temin, H.M. The bovine leukemia virus encapsidation signal is discontinuous and extends into the 5’ end of the gag gene. J. Virol. 1995, 69, 3282–3289. [Google Scholar] [PubMed]
- Jewell, N.A.; Mansky, L.M. Packaging of heterologous RNAs by a minimal bovine leukemia virus RNA packaging signal into virus particles. Arch. Virol. 2005, 150, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M.; Wisniewski, R.M. The bovine leukemia virus encapsidation signal is composed of RNA secondary structures. J. Virol. 1998, 72, 3196–3204. [Google Scholar]
- Mansky, L.M.; Gajary, L.C. The primary nucleotide sequence of the bovine leukemia virus RNA packaging signal can influence efficient RNA packaging and virus replication. Virology 2002, 301, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Mann, R.; Mulligan, R.C.; Baltimore, D. Construction of a retrovirus packaging mutant and its use to produce helper-free defective retrovirus. Cell 1983, 33, 153–159. [Google Scholar] [CrossRef]
- Adam, M.A.; Miller, A.D. Identification of a signal in a murine retrovirus that is sufficient for packaging of nonretroviral RNA into virions. J. Virol. 1988, 62, 3802–3806. [Google Scholar] [PubMed]
- Miyazaki, Y.; Irobalieva, R.N.; Tolbert, B.S.; Smalls-Mantey, A.; Iyalla, K.; Loeliger, K.; D’Souza, V.; Khant, H.; Schmid, M.F.; Garcia, E.L.; et al. Structure of a conserved retroviral RNA packaging element by NMR spectroscopy and cryo-electron tomography. J. Mol. Biol. 2010, 404, 751–772. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Temin, H.M. Encapsidation sequences for spleen necrosis virus, an avian retrovirus, are between the 5’ long terminal repeat and the start of the gag gene. Proc. Natl. Acad. Sci. USA 1982, 79, 5986–5990. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Temin, H.M. A double hairpin structure is necessary for the efficient encapsidation of spleen necrosis virus retroviral RNA. EMBO J. 1994, 13, 713–726. [Google Scholar] [PubMed]
- Katz, R.A.; Cullen, B.R.; Malavarca, R.; Skalka, A.M. Role of the avian retrovirus mRNA leader in expression: evidence for novel translational control. Mol. Cell. Biol. 1986, 6, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.D.; Linial, M.L. Secondary Structure Analysis of a Minimal Avian Leukosis-Sarcoma Virus Packaging Signal. J. Virol. 2000, 74, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Bean, R.L.; Vogt, V.M.; Summers, M.F. Solution Structure of the Rous Sarcoma Virus Nucleocapsid Protein:μΨ RNA Packaging Signal Complex. J. Mol. Biol. 2007, 365, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Konings, D.A.; Nash, M.A.; Maizel, J.V.; Arlinghaus, R.B. Novel GACG-hairpin pair motif in the 5’ untranslated region of type C retroviruses related to murine leukemia virus. J. Virol. 1992, 66, 632–640. [Google Scholar] [PubMed]
- Lever, A.; Gottlinger, H.; Haseltine, W.; Sodroski, J. Identification of a sequence required for efficient packaging of human immunodeficiency virus type 1 RNA into virions. J. Virol. 1989, 63, 4085–4087. [Google Scholar] [PubMed]
- Clavel, F.; Orenstein, J.M. A mutant of human immunodeficiency virus with reduced RNA packaging and abnormal particle morphology. J. Virol. 1990, 64, 5230–5234. [Google Scholar] [PubMed]
- Aldovini, A.; Young, R.A. Mutations of RNA and protein sequences involved in human immunodeficiency virus type 1 packaging result in production of noninfectious virus. J. Virol. 1990, 64, 1920–1926. [Google Scholar] [PubMed]
- Parolin, C.; Dorfman, T.; Palú, G.; Göttlinger, H.; Sodroski, J. Analysis in human immunodeficiency virus type 1 vectors of cis-acting sequences that affect gene transfer into human lymphocytes. J. Virol. 1994, 68, 3888–3895. [Google Scholar] [PubMed]
- Bender, M.A.; Palmer, T.D.; Gelinas, R.E.; Miller, A.D. Evidence that the packaging signal of Moloney murine leukemia virus extends into the gag region. J. Virol. 1987, 61, 1639–1646. [Google Scholar] [PubMed]
- Das, A.T.; Klaver, B.; Klasens, B.I.; van Wamel, J.L.; Berkhout, B. A conserved hairpin motif in the R-U5 region of the human immunodeficiency virus type 1 RNA genome is essential for replication. J. Virol. 1997, 71, 2346–2356. [Google Scholar]
- Aktar, S.J.; Jabeen, A.; Ali, L.M.; Vivet-Boudou, V.; Marquet, R.; Rizvi, T.A. SHAPE analysis of the 5’ end of the Mason-Pfizer monkey virus (MPMV) genomic RNA reveals structural elements required for genome dimerization. RNA 2013, 19, 1648–1658. [Google Scholar] [CrossRef] [PubMed]
- Aktar, S.J.; Vivet-Boudou, V.; Ali, L.M.; Jabeen, A.; Kalloush, R.M.; Richer, D.; Mustafa, F.; Marquet, R.; Rizvi, T.A. Structural basis of genomic RNA (gRNA) dimerization and packaging determinants of mouse mammary tumor virus (MMTV). Retrovirology 2014, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Paillart, J.-C.; Skripkin, E.; Ehresmann, B.; Ehresmann, C.; Marquet, R. In vitro evidence for a long range pseudoknot in the 5’-untranslated and matrix coding regions of HIV-1 genomic RNA. J. Biol. Chem. 2002, 277, 5995–6004. [Google Scholar] [CrossRef] [PubMed]
- Abbink, T.E.M.; Berkhout, B. A novel long distance base-pairing interaction in human immunodeficiency virus type 1 RNA occludes the Gag start codon. J. Biol. Chem. 2003, 278, 11601–11611. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, J.C.; Ghazawi, A.; Cheung, W.K.S.; Phillip, P.S.; Rizvi, T.A.; Lever, A.M.L. The secondary structure of the 5’ end of the FIV genome reveals a long-range interaction between R/U5 and gag sequences, and a large, stable stem-loop. RNA 2008, 14, 2597–2608. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, J.C.; Tanner, S.J.; Legiewicz, M.; Phillip, P.S.; Rizvi, T.A.; Le Grice, S.F.J.; Lever, A.M.L. SHAPE analysis of the FIV Leader RNA reveals a structural switch potentially controlling viral packaging and genome dimerization. Nucleic Acids Res. 2011, 39, 6692–6704. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, N.A.; Busan, S.; Rice, G.M.; Nelson, J.A.E.; Weeks, K.M. RNA motif discovery by SHAPE and mutational profiling (SHAPE-MaP). Nat. Methods 2014, 11, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Smyth, R.P.; Despons, L.; Huili, G.; Bernacchi, S.; Hijnen, M.; Mak, J.; Jossinet, F.; Weixi, L.; Paillart, J.-C.; von Kleist, M.; Marquet, R. Mutational interference mapping experiment (MIME) for studying RNA structure and function. Nat. Methods 2015, 12, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Keane, S.C.; Heng, X.; Lu, K.; Kharytonchyk, S.; Ramakrishnan, V.; Carter, G.; Barton, S.; Hosic, A.; Florwick, A.; Santos, J.; et al. RNA structure. Structure of the HIV-1 RNA packaging signal. Science 2015, 348, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Liu, Y.; Marchant, J.; Monti, S.; Seu, M.; Zaki, J.; Yang, A.L.; Bohn, J.; Ramakrishnan, V.; Singh, R.; et al. Conserved determinants of lentiviral genome dimerization. Retrovirology 2015, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Kalloush, R.M.; Vivet-Boudou, V.; Ali, L.M.; Mustafa, F.; Marquet, R.; Rizvi, T.A. Packaging of Mason-Pfizer monkey virus (MPMV) genomic RNA depends upon conserved long-range interactions (LRIs) between U5 and gag sequences. RNA 2016, 22, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Archer, E.J.; Simpson, M.A.; Watts, N.J.; O’Kane, R.; Wang, B.; Erie, D.A.; McPherson, A.; Weeks, K.M. Long-Range Architecture in a Viral RNA Genome. Biochemistry 2013, 52, 3182–3190. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Kafaie, J.; Laughrea, M. Role of the 5’ TAR stem--loop and the U5-AUG duplex in dimerization of HIV-1 genomic RNA. Biochemistry 2008, 47, 3283–3293. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, T.A.; Kenyon, J.C.; Ali, J.; Aktar, S.J.; Phillip, P.S.; Ghazawi, A.; Mustafa, F.; Lever, A.M.L. Optimal Packaging of FIV Genomic RNA Depends upon a Conserved Long-range Interaction and a Palindromic Sequence within gag. J. Mol. Biol. 2010, 403, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Wahab, E.W.; Smyth, R.P.; Mailler, E.; Bernacchi, S.; Vivet-Boudou, V.; Hijnen, M.; Jossinet, F.; Mak, J.; Paillart, J.-C.; Marquet, R. Specific recognition of the HIV-1 genomic RNA by the Gag precursor. Nat. Commun. 2014, 5, 4304. [Google Scholar] [CrossRef] [PubMed]
- Linial, M.L.; Miller, A.D. Retroviral RNA packaging: sequence requirements and implications. Curr. Top. Microbiol. Immunol. 1990, 157, 125–152. [Google Scholar] [PubMed]
- Guesdon, F.M.J.; Greatorex, J.; Rhee, S.R.; Fisher, R.; Hunter, E.; Lever, A.M.L. Sequences in the 5′ Leader of Mason-Pfizer Monkey Virus Which Affect Viral Particle Production and Genomic RNA Packaging: Development of MPMV Packaging Cell Lines. Virology 2001, 288, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Strappe, P.M.; Greatorex, J.; Thomas, J.; Biswas, P.; McCann, E.; Lever, A.M.L. The packaging signal of simian immunodeficiency virus is upstream of the major splice donor at a distance from the RNA cap site similar to that of human immunodeficiency virus types 1 and 2. J. Gen. Virol. 2003, 84, 2423–2430. [Google Scholar] [CrossRef] [PubMed]
- Balvay, L.; Lastra, M.L.; Sargueil, B.; Darlix, J.-L.; Ohlmann, T. Translational control of retroviruses. Nat. Rev. Microbiol. 2007, 5, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Linial, M.; Brown, S.; Neiman, P. A nonconditional mutant of Rous sarcoma virus containing defective polymerase. Virology 1978, 87, 130–141. [Google Scholar] [CrossRef]
- Donzé, O.; Spahr, P.F. Role of the open reading frames of Rous sarcoma virus leader RNA in translation and genome packaging. EMBO J. 1992, 11, 3747–3757. [Google Scholar] [PubMed]
- Donzé, O.; Damay, P.; Spahr, P.F. The first and third uORFs in RSV leader RNA are efficiently translated: implications for translational regulation and viral RNA packaging. Nucleic Acids Res. 1995, 23, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Kaye, J.F.; Lever, A.M. Human immunodeficiency virus types 1 and 2 differ in the predominant mechanism used for selection of genomic RNA for encapsidation. J. Virol. 1999, 73, 3023–3031. [Google Scholar] [PubMed]
- Luban, J.; Goff, S.P. Mutational analysis of cis-acting packaging signals in human immunodeficiency virus type 1 RNA. J. Virol. 1994, 68, 3784–3793. [Google Scholar] [PubMed]
- Ott, D.E.; Coren, L.V.; Sowder, R.C.; Adams, J.; Schubert, U. Retroviruses have differing requirements for proteasome function in the budding process. J. Virol. 2003, 77, 3384–3393. [Google Scholar] [CrossRef] [PubMed]
- Ott, D.E.; Coren, L.V.; Gagliardi, T.D.; Nagashima, K. Heterologous late-domain sequences have various abilities to promote budding of human immunodeficiency virus type 1. J. Virol. 2005, 79, 9038–9045. [Google Scholar] [CrossRef] [PubMed]
- Swanstrom, R.; Wills, J.W. Synthesis, Assembly, and Processing of Viral Proteins. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Vogt, V.M. Retroviral Virions and Genomes. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Green, L.M.; Berg, J.M. A retroviral Cys-Xaa2-Cys-Xaa4-His-Xaa4-Cys peptide binds metal ions: spectroscopic studies and a proposed three-dimensional structure. Proc. Natl. Acad. Sci. USA 1989, 86, 4047–4051. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.E.; Copeland, T.D.; Sowder, R.C.; Smythers, G.W.; Oroszlan, S. Primary structure of the low molecular weight nucleic acid-binding proteins of murine leukemia viruses. J. Biol. Chem. 1981, 256, 8400–8406. [Google Scholar] [PubMed]
- Bess, J.W.; Powell, P.J.; Issaq, H.J.; Schumack, L.J.; Grimes, M.K.; Henderson, L.E.; Arthur, L.O. Tightly bound zinc in human immunodeficiency virus type 1, human T-cell leukemia virus type I, and other retroviruses. J. Virol. 1992, 66, 840–847. [Google Scholar] [PubMed]
- Chance, M.R.; Sagi, I.; Wirt, M.D.; Frisbie, S.M.; Scheuring, E.; Chen, E.; Bess, J.W.; Henderson, L.E.; Arthur, L.O.; South, T.L. Extended x-ray absorption fine structure studies of a retrovirus: equine infectious anemia virus cysteine arrays are coordinated to zinc. Proc. Natl. Acad. Sci. USA 1992, 89, 10041–10045. [Google Scholar] [CrossRef] [PubMed]
- Summers, M.F.; Henderson, L.E.; Chance, M.R.; Bess, J.W.; South, T.L.; Blake, P.R.; Sagi, I.; Perez-Alvarado, G.; Sowder, R.C.; Hare, D.R. Nucleocapsid zinc fingers detected in retroviruses: EXAFS studies of intact viruses and the solution-state structure of the nucleocapsid protein from HIV-1. Protein Sci. 1992, 1, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Poon, D.T.; Wu, J.; Aldovini, A. Charged amino acid residues of human immunodeficiency virus type 1 nucleocapsid p7 protein involved in RNA packaging and infectivity. J. Virol. 1996, 70, 6607–6616. [Google Scholar] [PubMed]
- Gorelick, R.J.; Henderson, L.E.; Hanser, J.P.; Rein, A. Point mutants of Moloney murine leukemia virus that fail to package viral RNA: evidence for specific RNA recognition by a “zinc finger-like” protein sequence. Proc. Natl. Acad. Sci. USA 1988, 85, 8420–8424. [Google Scholar] [CrossRef] [PubMed]
- Jentoft, J.E.; Smith, L.M.; Fu, X.D.; Johnson, M.; Leis, J. Conserved cysteine and histidine residues of the avian myeloblastosis virus nucleocapsid protein are essential for viral replication but are not “zinc-binding fingers.”. Proc. Natl. Acad. Sci. USA 1988, 85, 7094–7098. [Google Scholar] [CrossRef] [PubMed]
- Méric, C.; Gouilloud, E.; Spahr, P.F. Mutations in Rous sarcoma virus nucleocapsid protein p12 (NC): deletions of Cys-His boxes. J. Virol. 1988, 62, 3328–3333. [Google Scholar] [PubMed]
- Gorelick, R.J.; Nigida, S.M.J.; Bess, J.W.; Arthur, L.O.; Henderson, L.E.; Rein, A. Noninfectious human immunodeficiency virus type 1 mutants deficient in genomic RNA. J. Virol. 1990, 64, 3207–11. [Google Scholar] [PubMed]
- Dorfman, T.; Luban, J.; Goff, S.P.; Haseltine, W.A.; Göttlinger, H.G. Mapping of functionally important residues of a cysteine-histidine box in the human immunodeficiency virus type 1 nucleocapsid protein. J. Virol. 1993, 67, 6159–6169. [Google Scholar] [PubMed]
- Gorelick, R.J.; Chabot, D.J.; Rein, A.; Henderson, L.E.; Arthur, L.O. The two zinc fingers in the human immunodeficiency virus type 1 nucleocapsid protein are not functionally equivalent. J. Virol. 1993, 67, 4027–4036. [Google Scholar] [PubMed]
- Wang, C.T.; Barklis, E. Assembly, processing, and infectivity of human immunodeficiency virus type 1 Gag mutants. J. Virol. 1993, 67, 4264–4273. [Google Scholar] [PubMed]
- Zhang, Y.; Barklis, E. Nucleocapsid protein effects on the specificity of retrovirus RNA encapsidation. J. Virol. 1995, 69, 5716–5722. [Google Scholar] [PubMed]
- Mizuno, A.; Ido, E.; Goto, T.; Kuwata, T.; Nakai, M.; Hayami, M. Mutational analysis of two zinc finger motifs in HIV type 1 nucleocapsid proteins: effects on proteolytic processing of Gag precursors and particle formation. AIDS Res. Hum. Retroviruses 1996, 12, 793–800. [Google Scholar] [CrossRef] [PubMed]
- De Guzman, R.N.; Turner, R.B.; Summers, M.F. Protein-RNA recognition. Biopolymers 1998, 48, 181–195. [Google Scholar] [CrossRef]
- Tanchou, V.; Decimo, D.; Péchoux, C.; Lener, D.; Rogemond, V.; Berthoux, L.; Ottmann, M.; Darlix, J.L. Role of the N-terminal zinc finger of human immunodeficiency virus type 1 nucleocapsid protein in virus structure and replication. J. Virol. 1998, 72, 4442–4447. [Google Scholar] [PubMed]
- Gorelick, R.J.; Gagliardi, T.D.; Bosche, W.J.; Wiltrout, T.A.; Coren, L.V.; Chabot, D.J.; Lifson, J.D.; Henderson, L.E.; Arthur, L.O. Strict conservation of the retroviral nucleocapsid protein zinc finger is strongly influenced by its role in viral infection processes: characterization of HIV-1 particles containing mutant nucleocapsid zinc-coordinating sequences. Virology 1999, 256, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wu, T.; Anderson, J.; Kane, B.F.; Johnson, D.G.; Gorelick, R.J.; Henderson, L.E.; Levin, J.G. Zinc finger structures in the human immunodeficiency virus type 1 nucleocapsid protein facilitate efficient minus- and plus-strand transfer. J. Virol. 2000, 74, 8980–8988. [Google Scholar] [CrossRef] [PubMed]
- Housset, V.; De Rocquigny, H.; Roques, B.P.; Darlix, J.L. Basic amino acids flanking the zinc finger of Moloney murine leukemia virus nucleocapsid protein NCp10 are critical for virus infectivity. J. Virol. 1993, 67, 2537–2545. [Google Scholar] [PubMed]
- Cimarelli, A.; Sandin, S.; Höglund, S.; Luban, J. Basic residues in human immunodeficiency virus type 1 nucleocapsid promote virion assembly via interaction with RNA. J. Virol. 2000, 74, 3046–3057. [Google Scholar] [CrossRef] [PubMed]
- Gonsky, J.; Bacharach, E.; Goff, S.P. Identification of residues of the Moloney murine leukemia virus nucleocapsid critical for viral DNA synthesis in vivo. J. Virol. 2001, 75, 2616–2626. [Google Scholar] [CrossRef] [PubMed]
- Dupraz, P.; Spahr, P.F. Specificity of Rous sarcoma virus nucleocapsid protein in genomic RNA packaging. J. Virol. 1992, 66, 4662–4670. [Google Scholar] [PubMed]
- Berkowitz, R.D.; Ohagen, A.; Höglund, S.; Goff, S.P. Retroviral nucleocapsid domains mediate the specific recognition of genomic viral RNAs by chimeric Gag polyproteins during RNA packaging in vivo. J. Virol. 1995, 69, 6445–6456. [Google Scholar] [PubMed]
- Fu, W.; Hu, W.-S. Functional Replacement of Nucleocapsid Flanking Regions by Heterologous Counterparts with Divergent Primary Sequences: Effects of Chimeric Nucleocapsid on the Retroviral Replication Cycle. J. Virol. 2003, 77, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Freed, E.O. HIV-1 gag proteins: diverse functions in the virus life cycle. Virology 1998, 251, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Garcia, E.L.; King, S.R.; Iyalla, K.; Loeliger, K.; Starck, P.; Syed, S.; Telesnitsky, A.; Summers, M.F. An RNA structural switch regulates diploid genome packaging by Moloney murine leukemia virus. J. Mol. Biol. 2010, 396, 141–152. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, V.; Summers, M.F. Structural basis for packaging the dimeric genome of Moloney murine leukaemia virus. Nature 2004, 431, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Poon, D.T.; Li, G.; Aldovini, A. Nucleocapsid and matrix protein contributions to selective human immunodeficiency virus type 1 genomic RNA packaging. J. Virol. 1998, 72, 1983–1993. [Google Scholar] [PubMed]
- Guo, X.; Roy, B.B.; Hu, J.; Roldan, A.; Wainberg, M.A.; Liang, C. The R362A mutation at the C-terminus of CA inhibits packaging of human immunodeficiency virus type 1 RNA. Virology 2005, 343, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.S.; Roldan, A.; Detorio, M.; Hu, J.; Wainberg, M.A.; Liang, C. Effects of a single amino acid substitution within the p2 region of human immunodeficiency virus type 1 on packaging of spliced viral RNA. J. Virol. 2003, 77, 12986–12995. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.B.; Russell, R.S.; Turner, D.; Liang, C. The T12I mutation within the SP1 region of Gag restricts packaging of spliced viral RNA into human immunodeficiency virus type 1 with mutated RNA packaging signals and mutated nucleocapsid sequence. Virology 2006, 344, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Butterfield-Gerson, K.L.; Scheifele, L.Z.; Ryan, E.P.; Hopper, A.K.; Parent, L.J. Importin-beta family members mediate alpharetrovirus gag nuclear entry via interactions with matrix and nucleocapsid. J. Virol. 2006, 80, 1798–1806. [Google Scholar] [CrossRef] [PubMed]
- Kenney, S.P.; Lochmann, T.L.; Schmid, C.L.; Parent, L.J. Intermolecular interactions between retroviral Gag proteins in the nucleus. J. Virol. 2008, 82, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Gudleski, N.; Flanagan, J.M.; Ryan, E.P.; Bewley, M.C.; Parent, L.J. Directionality of nucleocytoplasmic transport of the retroviral gag protein depends on sequential binding of karyopherins and viral RNA. Proc. Natl. Acad. Sci. USA 2010, 107, 9358–9363. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R. Nuclear mRNA export: insights from virology. Trends Biochem. Sci. 2003, 28, 419–424. [Google Scholar] [CrossRef]
- Jouvenet, N.; Simon, S.M.; Bieniasz, P.D. Imaging the interaction of HIV-1 genomes and Gag during assembly of individual viral particles. Proc. Natl. Acad. Sci. USA 2009, 106, 19114–19119. [Google Scholar] [CrossRef] [PubMed]
- Kemler, I.; Meehan, A.; Poeschla, E.M. Live-cell coimaging of the genomic RNAs and Gag proteins of two lentiviruses. J. Virol. 2010, 84, 6352–6366. [Google Scholar] [CrossRef] [PubMed]
- Saad, J.S.; Miller, J.; Tai, J.; Kim, A.; Ghanam, R.H.; Summers, M.F. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc. Natl. Acad. Sci. USA 2006, 103, 11364–11369. [Google Scholar] [CrossRef] [PubMed]
- Ono, A.; Ablan, S.D.; Lockett, S.J.; Nagashima, K.; Freed, E.O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc. Natl. Acad. Sci. USA 2004, 101, 14889–14894. [Google Scholar] [CrossRef] [PubMed]
- Strappe, P.M.; Hampton, D.W.; Brown, D.; Cachon-Gonzalez, B.; Caldwell, M.; Fawcett, J.W.; Lever, A.M. Identification of unique reciprocal and non-reciprocal cross packaging relationships between HIV-1, HIV-2 and SIV reveals an efficient SIV/HIV-2 lentiviral vector system with highly favourable features for in vivo testing and clinical usage. Retrovirology 2005, 2, 55. [Google Scholar] [CrossRef] [PubMed]
- Certo, J.L.; Shook, B.F.; Yin, P.D.; Snider, J.T.; Hu, W.-S. Nonreciprocal Pseudotyping: Murine Leukemia Virus Proteins Cannot Efficiently Package Spleen Necrosis Virus-Based Vector RNA. J. Virol. 1998, 72, 5408–5413. [Google Scholar] [PubMed]
- Kewalramani, V.N.; Panganiban, A.T.; Emerman, M. Spleen necrosis virus, an avian immunosuppressive retrovirus, shares a receptor with the type D simian retroviruses. J. Virol. 1992, 66, 3026–3031. [Google Scholar] [PubMed]
- Dougherty, J.P.; Wisniewski, R.; Yang, S.L.; Rhode, B.W.; Temin, H.M. New retrovirus helper cells with almost no nucleotide sequence homology to retrovirus vectors. J. Virol. 1989, 63, 3209–3212. [Google Scholar] [PubMed]
- Embretson, J.E.; Temin, H.M. Lack of competition results in efficient packaging of heterologous murine retroviral RNAs and reticuloendotheliosis virus encapsidation-minus RNAs by the reticuloendotheliosis virus helper cell line. J. Virol. 1987, 61, 2675–2683. [Google Scholar] [PubMed]
- Chakrabarti, R.; Hofman, F.M.; Pandey, R.; Mathes, L.E.; Roy-Burman, P. Recombination between feline exogenous and endogenous retroviral sequences generates tropism for cerebral endothelial cells. Am. J. Pathol. 1994, 144, 348–358. [Google Scholar] [PubMed]
- Chong, H.; Starkey, W.; Vile, R.G. A replication-competent retrovirus arising from a split-function packaging cell line was generated by recombination events between the vector, one of the packaging constructs, and endogenous retroviral sequences. J. Virol. 1998, 72, 2663–2670. [Google Scholar] [PubMed]
- Bukovsky, A.A.; Song, J.P.; Naldini, L. Interaction of human immunodeficiency virus-derived vectors with wild-type virus in transduced cells. J. Virol. 1999, 73, 7087–7092. [Google Scholar] [PubMed]
- Bartosch, B.; Stefanidis, D.; Myers, R.; Weiss, R.; Patience, C.; Takeuchi, Y. Evidence and consequence of porcine endogenous retrovirus recombination. J. Virol. 2004, 78, 13880–13890. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.T.; Garcia, J.V. Lentivirus vector mobilization and spread by human immunodeficiency virus. Hum. Gene Ther. 2000, 11, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.H.; Alamgir, A.S.M.; Owens, N.; Weber, N.; Virtaneva, K.; Barbian, K.; Babar, A.; Malik, F.; Rosenke, K. Mobilization of endogenous retroviruses in mice after infection with an exogenous retrovirus. J. Virol. 2009, 83, 2429–2435. [Google Scholar] [CrossRef] [PubMed]
- Dull, T.; Zufferey, R.; Kelly, M.; Mandel, R.J.; Nguyen, M.; Trono, D.; Naldini, L. A third-generation lentivirus vector with a conditional packaging system. J. Virol. 1998, 72, 8463–8471. [Google Scholar] [PubMed]
- Ismail, S.I.; Kingsman, S.M.; Kingsman, A.J.; Uden, M. Split-intron retroviral vectors: enhanced expression with improved safety. J. Virol. 2000, 74, 2365–2371. [Google Scholar] [CrossRef] [PubMed]
- Zufferey, R.; Dull, T.; Mandel, R.J.; Bukovsky, A.; Quiroz, D.; Naldini, L.; Trono, D. Self-inactivating lentivirus vector for safe and efficient in vivo gene delivery. J. Virol. 1998, 72, 9873–9880. [Google Scholar] [PubMed]
- Westerman, K.A.; Ao, Z.; Cohen, E.A.; Leboulch, P. Design of a trans protease lentiviral packaging system that produces high titer virus. Retrovirology 2007, 4, 96. [Google Scholar] [CrossRef] [PubMed]
- Logan, A.C.; Haas, D.L.; Kafri, T.; Kohn, D.B. Integrated self-inactivating lentiviral vectors produce full-length genomic transcripts competent for encapsidation and integration. J. Virol. 2004, 78, 8421–8436. [Google Scholar] [CrossRef] [PubMed]
- Hanawa, H.; Persons, D.A.; Nienhuis, A.W. Mobilization and mechanism of transcription of integrated self-inactivating lentiviral vectors. J. Virol. 2005, 79, 8410–8421. [Google Scholar] [CrossRef] [PubMed]
- Schambach, A.; Galla, M.; Maetzig, T.; Loew, R.; Baum, C. Improving transcriptional termination of self-inactivating gamma-retroviral and lentiviral vectors. Mol. Ther. J. Am. Soc. Gene Ther. 2007, 15, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Goujon, C.; Jarrosson-Wuilleme, L.; Bernaud, J.; Rigal, D.; Darlix, J.-L.; Cimarelli, A. Heterologous human immunodeficiency virus type 1 lentiviral vectors packaging a simian immunodeficiency virus-derived genome display a specific postentry transduction defect in dendritic cells. J. Virol. 2003, 77, 9295–9304. [Google Scholar] [CrossRef] [PubMed]
- Poeschla, E.M.; Wong-Staal, F.; Looney, D.J. Efficient transduction of nondividing human cells by feline immunodeficiency virus lentiviral vectors. Nat. Med. 1998, 4, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.V.; Gilbert, J.; Wong-Staal, F.; Gasmi, M.; Looney, D.J. Transduction of cell lines and primary cells by FIV-packaged HIV vectors. Mol. Ther. J. Am. Soc. Gene Ther. 2004, 10, 181–190. [Google Scholar] [CrossRef]
- Delviks, K.A.; Hu, W.S.; Pathak, V.K. Psi- vectors: murine leukemia virus-based self-inactivating and self-activating retroviral vectors. J. Virol. 1997, 71, 6218–6224. [Google Scholar] [PubMed]
- Saller, R.M.; Oztürk, F.; Salmons, B.; Günzburg, W.H. Construction and characterization of a hybrid mouse mammary tumor virus/murine leukemia virus-based retroviral vector. J. Virol. 1998, 72, 1699–1703. [Google Scholar] [PubMed]
- Beasley, B.E.; Hu, W.-S. cis-Acting elements important for retroviral RNA packaging specificity. J. Virol. 2002, 76, 4950–4960. [Google Scholar] [CrossRef] [PubMed]
- Browning, M.T.; Schmidt, R.D.; Lew, K.A.; Rizvi, T.A. Primate and feline lentivirus vector RNA packaging and propagation by heterologous lentivirus virions. J. Virol. 2001, 75, 5129–5140. [Google Scholar] [CrossRef] [PubMed]
- Kol, N.; Gladnikoff, M.; Barlam, D.; Shneck, R.Z.; Rein, A.; Rousso, I. Mechanical properties of murine leukemia virus particles: effect of maturation. Biophys. J. 2006, 91, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Günzburg, W.H.; Salmons, B. Mouse mammary tumor virus mediated transfer and expression of neomycin resistance to infected cultured cells. Virology 1986, 155, 236–248. [Google Scholar] [CrossRef]
- Masuda, T.; Kuroda, M.J.; Harada, S. Specific and independent recognition of U3 and U5 att sites by human immunodeficiency virus type 1 integrase in vivo. J. Virol. 1998, 72, 8396–8402. [Google Scholar] [PubMed]
- Hlavaty, J.; Stracke, A.; Klein, D.; Salmons, B.; Günzburg, W.H.; Renner, M. Multiple modifications allow high-titer production of retroviral vectors carrying heterologous regulatory elements. J. Virol. 2004, 78, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- Ghazawi, A.; Mustafa, F.; Phillip, P.S.; Jayanth, P.; Ali, J.; Rizvi, T.A. Both the 5’ and 3’ LTRs of FIV contain minor RNA encapsidation determinants compared to the two core packaging determinants within the 5’ untranslated region and gag. Microbes Infect. 2006, 8, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Adkins, B.; Hunter, T. Identification of a packaged cellular mRNA in virions of rous sarcoma virus. J. Virol. 1981, 39, 471–480. [Google Scholar] [PubMed]
- Aronoff, R.; Linial, M. Specificity of retroviral RNA packaging. J. Virol. 1991, 65, 71–80. [Google Scholar] [PubMed]
- Bishop, J.M.; Levinson, W.E.; Sullivan, D.; Fanshier, L.; Quintrell, N.; Jackson, J. The low molecular weight RNAs of Rous sarcoma virus. II. The 7 S RNA. Virology 1970, 42, 927–937. [Google Scholar] [CrossRef]
- Gallis, B.; Linial, M.; Eisenman, R. An avian oncovirus mutant deficient in genomic RNA: characterization of the packaged RNA as cellular messenger RNA. Virology 1979, 94, 146–161. [Google Scholar] [CrossRef]
- Ikawa, Y.; Ross, J.; Leder, P. An association between globin messenger RNA and 60S RNA derived from Friend leukemia virus. Proc. Natl. Acad. Sci. USA 1974, 71, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Muriaux, D.; Mirro, J.; Harvin, D.; Rein, A. RNA is a structural element in retrovirus particles. Proc. Natl. Acad. Sci. USA 2001, 98, 5246–5251. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.C.; Mullin, B.C. Transfer RNA into RNA tumor viruses. Prog. Nucleic Acid Res. Mol. Biol. 1977, 20, 131–160. [Google Scholar] [PubMed]
- Martin, P.; Henry, C.; Ferre, F.; Bechade, C.; Begue, A.; Calothy, C.; Debuire, B.; Stehelin, D.; Saule, S. Characterization of a myc-containing retrovirus generated by propagation of an MH2 viral subgenomic RNA. J. Virol. 1986, 57, 1191–1194. [Google Scholar] [PubMed]
- Stacey, D.W. Messenger activity of virion RNA for avian leukosis viral envelope glycoprotein. J. Virol. 1979, 29, 949–956. [Google Scholar] [PubMed]
- Svoboda, J.; Dvorák, M.; Guntaka, R.; Geryk, J. Transmission of (LTR, v-src, LTR) without recombination with a helper virus. Virology 1986, 153, 314–317. [Google Scholar] [CrossRef]
- Onafuwa-Nuga, A.A.; King, S.R.; Telesnitsky, A. Nonrandom packaging of host RNAs in moloney murine leukemia virus. J. Virol. 2005, 79, 13528–13537. [Google Scholar] [CrossRef] [PubMed]
- Eckwahl, M.J.; Telesnitsky, A.; Wolin, S.L. Host RNA Packaging by Retroviruses: A Newly Synthesized Story. MBio 2016, 7, e02025-15. [Google Scholar] [CrossRef] [PubMed]
- Onafuwa-Nuga, A.; Telesnitsky, A. The remarkable frequency of human immunodeficiency virus type 1 genetic recombination. Microbiol. Mol. Biol. Rev. 2009, 73, 451–480, Table of Contents. [Google Scholar] [CrossRef] [PubMed]
- Marquet, R.; Baudin, F.; Gabus, C.; Darlix, J.L.; Mougel, M.; Ehresmann, C.; Ehresmann, B. Dimerization of human immunodeficiency virus (type 1) RNA: stimulation by cations and possible mechanism. Nucleic Acids Res. 1991, 19, 2349–2357. [Google Scholar] [CrossRef] [PubMed]
- Delviks-Frankenberry, K.; Galli, A.; Nikolaitchik, O.; Mens, H.; Pathak, V.K.; Hu, W.-S. Mechanisms and Factors that Influence High Frequency Retroviral Recombination. Viruses 2011, 3, 1650–1680. [Google Scholar] [CrossRef] [PubMed]
- Negroni, M.; Buc, H. Mechanisms of retroviral recombination. Annu. Rev. Genet. 2001, 35, 275–302. [Google Scholar] [CrossRef] [PubMed]
- Ni, N.; Nikolaitchik, O.A.; Dilley, K.A.; Chen, J.; Galli, A.; Fu, W.; Prasad, V.V.S.P.; Ptak, R.G.; Pathak, V.K.; Hu, W.-S. Mechanisms of human immunodeficiency virus type 2 RNA packaging: efficient trans packaging and selection of RNA copackaging partners. J. Virol. 2011, 85, 7603–7612. [Google Scholar] [CrossRef] [PubMed]
- Dilley, K.A.; Ni, N.; Nikolaitchik, O.A.; Chen, J.; Galli, A.; Hu, W.-S. Determining the frequency and mechanisms of HIV-1 and HIV-2 RNA copackaging by single-virion analysis. J. Virol. 2011, 85, 10499–10508. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, J.G.; Rasmussen, S.V.; Pedersen, F.S. Complementarity-directed RNA dimer-linkage promotes retroviral recombination in vivo. Nucleic Acids Res. 2004, 32, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.V.; Mikkelsen, J.G.; Pedersen, F.S. Modulation of homo- and heterodimerization of Harvey sarcoma virus RNA by GACG tetraloops and point mutations in palindromic sequences. J. Mol. Biol. 2002, 323, 613–628. [Google Scholar] [CrossRef]
- Vogt, P.K. Genetically stable reassortment of markers during mixed infection with avian tumor viruses. Virology 1971, 46, 947–952. [Google Scholar] [CrossRef]
- Clavel, F.; Hoggan, M.D.; Willey, R.L.; Strebel, K.; Martin, M.A.; Repaske, R. Genetic recombination of human immunodeficiency virus. J. Virol. 1989, 63, 1455–1459. [Google Scholar] [PubMed]
- Wong, P.K.; Russ, L.J.; McCarter, J.A. Rapid, selective procedure for isolation of spontaneous temperature-sensitive mutants of Moloney leukemia virus. Virology 1973, 51, 424–431. [Google Scholar] [CrossRef]
- Wyke, J.A. Temperature sensitive mutants of avian sarcoma viruses. Biochim. Biophys. Acta 1975, 417, 91–121. [Google Scholar] [CrossRef]
- Dang, Q.; Chen, J.; Unutmaz, D.; Coffin, J.M.; Pathak, V.K.; Powell, D.; KewalRamani, V.N.; Maldarelli, F.; Hu, W.-S. Nonrandom HIV-1 infection and double infection via direct and cell-mediated pathways. Proc. Natl. Acad. Sci. USA 2004, 101, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Dang, Q.; Unutmaz, D.; Pathak, V.K.; Maldarelli, F.; Powell, D.; Hu, W.-S. Mechanisms of nonrandom human immunodeficiency virus type 1 infection and double infection: preference in virus entry is important but is not the sole factor. J. Virol. 2005, 79, 4140–4149. [Google Scholar] [CrossRef] [PubMed]
- Del Portillo, A.; Tripodi, J.; Najfeld, V.; Wodarz, D.; Levy, D.N.; Chen, B.K. Multiploid inheritance of HIV-1 during cell-to-cell infection. J. Virol. 2011, 85, 7169–7176. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.A.; Mason, W.S.; Vogt, P.K. Genetic recombinants and heterozygotes derived from endogenous and exogenous avian RNA tumor viruses. Virology 1973, 52, 535–552. [Google Scholar] [CrossRef]
- Hu, W.S.; Temin, H.M. Genetic consequences of packaging two RNA genomes in one retroviral particle: pseudodiploidy and high rate of genetic recombination. Proc. Natl. Acad. Sci. USA 1990, 87, 1556–1560. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.K.; Hu, W.-S. “Might as Well Jump!” Template Switching by Retroviral Reverse Transcriptase, Defective Genome Formation, and Recombination. Semin. Virol. 1997, 8, 141–150. [Google Scholar] [CrossRef]
- Gilboa, E.; Mitra, S.W.; Goff, S.; Baltimore, D. A detailed model of reverse transcription and tests of crucial aspects. Cell 1979, 18, 93–100. [Google Scholar] [CrossRef]
- Goff, S.P. Retroviral reverse transcriptase: synthesis, structure, and function. J. Acquir. Immune Defic. Syndr. 1990, 3, 817–831. [Google Scholar] [PubMed]
- Yu, H.; Jetzt, A.E.; Ron, Y.; Preston, B.D.; Dougherty, J.P. The nature of human immunodeficiency virus type 1 strand transfers. J. Biol. Chem. 1998, 273, 28384–28391. [Google Scholar] [CrossRef] [PubMed]
- van Wamel, J.L.; Berkhout, B. The first strand transfer during HIV-1 reverse transcription can occur either intramolecularly or intermolecularly. Virology 1998, 244, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Stuhlmann, H.; Berg, P. Homologous recombination of copackaged retrovirus RNAs during reverse transcription. J. Virol. 1992, 66, 2378–2388. [Google Scholar] [PubMed]
- Hu, W.S.; Bowman, E.H.; Delviks, K.A.; Pathak, V.K. Homologous recombination occurs in a distinct retroviral subpopulation and exhibits high negative interference. J. Virol. 1997, 71, 6028–6036. [Google Scholar] [PubMed]
- Katz, R.A.; Skalka, A.M. Generation of diversity in retroviruses. Annu. Rev. Genet. 1990, 24, 409–445. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.A.; Teufel, R.J.; Yin, P.D.; Hu, W.S. Correlated template-switching events during minus-strand DNA synthesis: a mechanism for high negative interference during retroviral recombination. J. Virol. 1998, 72, 1186–1194. [Google Scholar] [PubMed]
- Zhang, J.; Tang, L.-Y.; Li, T.; Ma, Y.; Sapp, C.M. Most Retroviral Recombinations Occur during Minus-Strand DNA Synthesis. J. Virol. 2000, 74, 2313–2322. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.K.; Temin, H.M. Broad spectrum of in vivo forward mutations, hypermutations, and mutational hotspots in a retroviral shuttle vector after a single replication cycle: deletions and deletions with insertions. Proc. Natl. Acad. Sci. USA 1990, 87, 6024–6028. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathi, S.; Varela-Echavarría, A.; Ron, Y.; Preston, B.D.; Dougherty, J.P. Genetic rearrangements occurring during a single cycle of murine leukemia virus vector replication: characterization and implications. J. Virol. 1995, 69, 7991–8000. [Google Scholar] [PubMed]
- Panganiban, A.T.; Fiore, D. Ordered interstrand and intrastrand DNA transfer during reverse transcription. Science 1988, 241, 1064–1069. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.M.; Brown, A.M.; Ron, Y.; Dougherty, J.P. Spleen necrosis virus, an avian retrovirus, can infect primate cells. J. Virol. 1991, 65, 4769–4776. [Google Scholar] [PubMed]
- Chiu, C.N.; Mitra, R.; Chiu, I.-M. Exchange of genetic sequences of long terminal repeat and the env gene by a promiscuous primate type D retrovirus. Virus Res. 2003, 96, 107–111. [Google Scholar] [CrossRef]
- Wayengera, M. On the general theory of the origins of retroviruses. Theor. Biol. Med. Model. 2010, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Nikolaitchik, O.A.; Galli, A.; Moore, M.D.; Pathak, V.K.; Hu, W.-S. Multiple barriers to recombination between divergent HIV-1 variants revealed by a dual-marker recombination assay. J. Mol. Biol. 2011, 407, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Galli, A.; Kearney, M.; Nikolaitchik, O.A.; Yu, S.; Chin, M.P.S.; Maldarelli, F.; Coffin, J.M.; Pathak, V.K.; Hu, W.-S. Patterns of Human Immunodeficiency Virus type 1 recombination ex vivo provide evidence for coadaptation of distant sites, resulting in purifying selection for intersubtype recombinants during replication. J. Virol. 2010, 84, 7651–7661. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, T.; Wargo, H.; Hu, W.-S. High rates of human immunodeficiency virus type 1 recombination: near-random segregation of markers one kilobase apart in one round of viral replication. J. Virol. 2003, 77, 11193–11200. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.A.; An, W.; King, S.R.; Telesnitsky, A. Nonrandom dimerization of murine leukemia virus genomic RNAs. J. Virol. 2004, 78, 12129–12139. [Google Scholar] [CrossRef] [PubMed]
- Roda, R.H.; Balakrishnan, M.; Hanson, M.N.; Wöhrl, B.M.; Le Grice, S.F.J.; Roques, B.P.; Gorelick, R.J.; Bambara, R.A. Role of the Reverse Transcriptase, Nucleocapsid Protein, and Template Structure in the Two-step Transfer Mechanism in Retroviral Recombination. J. Biol. Chem. 2003, 278, 31536–31546. [Google Scholar] [CrossRef] [PubMed]
- Moumen, A.; Polomack, L.; Roques, B.; Buc, H.; Negroni, M. The HIV-1 repeated sequence R as a robust hot-spot for copy-choice recombination. Nucleic Acids Res. 2001, 29, 3814–3821. [Google Scholar] [CrossRef] [PubMed]
- Dykes, C.; Balakrishnan, M.; Planelles, V.; Zhu, Y.; Bambara, R.A.; Demeter, L.M. Identification of a preferred region for recombination and mutation in HIV-1 gag. Virology 2004, 326, 262–279. [Google Scholar] [CrossRef] [PubMed]
- Galli, A.; Lai, A.; Corvasce, S.; Saladini, F.; Riva, C.; Dehò, L.; Caramma, I.; Franzetti, M.; Romano, L.; Galli, M.; et al. Recombination analysis and structure prediction show correlation between breakpoint clusters and RNA hairpins in the pol gene of human immunodeficiency virus type 1 unique recombinant forms. J. Gen. Virol. 2008, 89, 3119–3125. [Google Scholar] [CrossRef] [PubMed]
- Galetto, R.; Moumen, A.; Giacomoni, V.; Véron, M.; Charneau, P.; Negroni, M. The structure of HIV-1 genomic RNA in the gp120 gene determines a recombination hot spot in vivo. J. Biol. Chem. 2004, 279, 36625–36632. [Google Scholar] [CrossRef] [PubMed]
- Galetto, R.; Giacomoni, V.; Véron, M.; Negroni, M. Dissection of a circumscribed recombination hot spot in HIV-1 after a single infectious cycle. J. Biol. Chem. 2006, 281, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Certo, J.L.; Kabdulov, T.O.; Paulson, M.L.; Anderson, J.A.; Hu, W.S. The nucleocapsid domain is responsible for the ability of spleen necrosis virus (SNV) Gag polyprotein to package both SNV and murine leukemia virus RNA. J. Virol. 1999, 73, 9170–9177. [Google Scholar] [PubMed]
- Méric, C.; Goff, S.P. Characterization of Moloney murine leukemia virus mutants with single-amino-acid substitutions in the Cys-His box of the nucleocapsid protein. J. Virol. 1989, 63, 1558–1568. [Google Scholar] [PubMed]
- Rein, A.; Henderson, L.E.; Levin, J.G. Nucleic-acid-chaperone activity of retroviral nucleocapsid proteins: significance for viral replication. Trends Biochem. Sci. 1998, 23, 297–301. [Google Scholar] [CrossRef]
- Zhang, Y.; Qian, H.; Love, Z.; Barklis, E. Analysis of the assembly function of the human immunodeficiency virus type 1 gag protein nucleocapsid domain. J. Virol. 1998, 72, 1782–1789. [Google Scholar] [PubMed]
- Darlix, J.L.; Lapadat-Tapolsky, M.; de Rocquigny, H.; Roques, B.P. First glimpses at structure-function relationships of the nucleocapsid protein of retroviruses. J. Mol. Biol. 1995, 254, 523–537. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
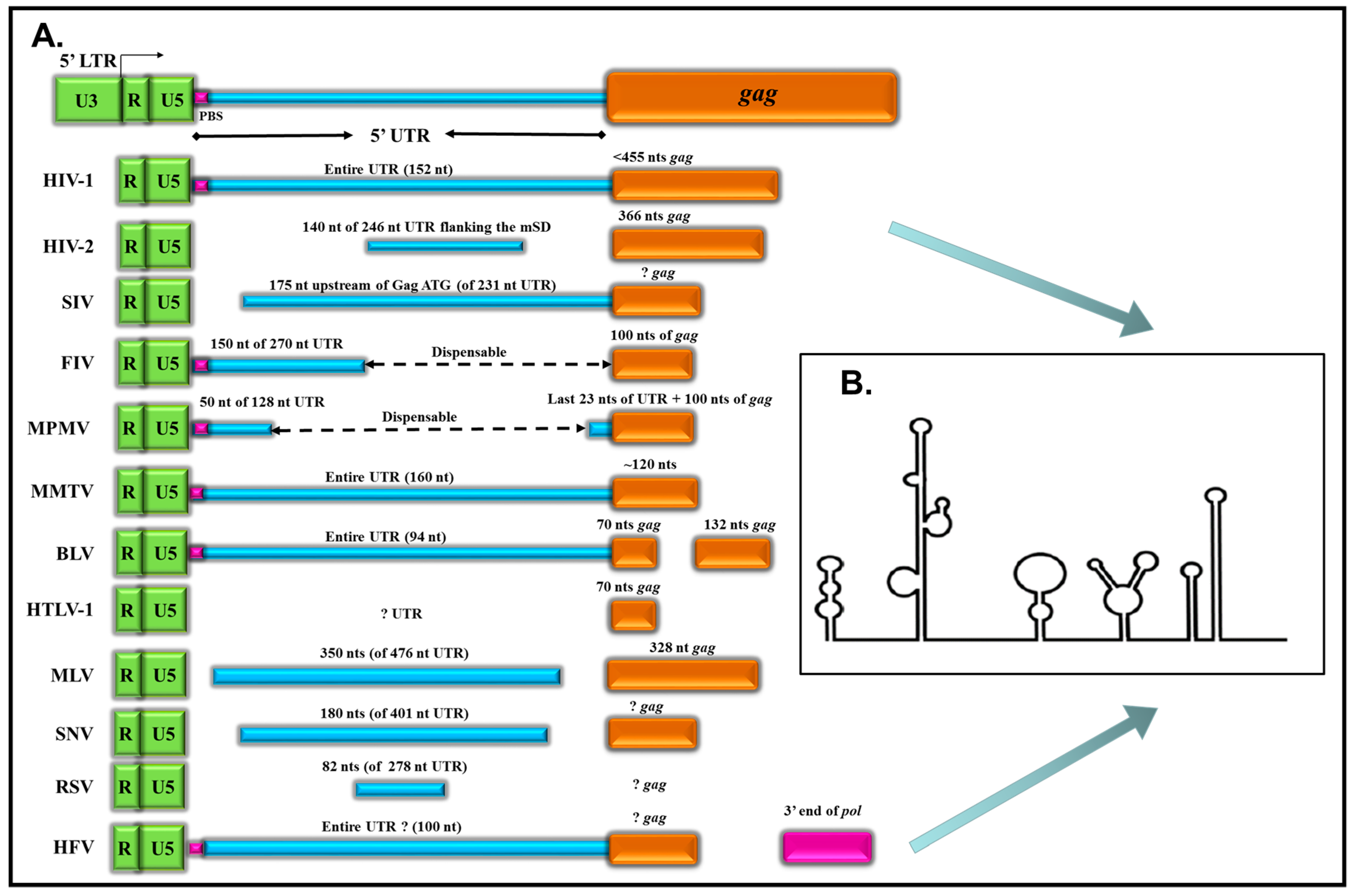
| Virus | Length of 5’ UTR* | 5’ UTR* and R/U5 Sequences Required | gag Sequences Required | Reference |
|---|---|---|---|---|
| HIV-1 | ~152 nts | Entire UTR, SL1 and SL3 | <455 nts | [18,29] |
| HIV-2 | ~246 nts | 140 nts | 366 nts | [40,41,42,43] |
| SIVmac239 | ~231 nts | 117–175 nts upstream of gag ATG (SL1, SL3 and SL4) | ? | [29,44,45] |
| FIV | ~270 nts | First 150 nts | 100 nts | [46] |
| MPMV | ~128 nts | First 50 nts + last 23 nts | 100 nts | [47,48] |
| MMTV | ~160 nts | Entire UTR | 120 nts | [49] |
| BLV | ~94 nts | Entire UTR | First 70 nts of gag (SL1 and SL2) + 132 nts (nt 1015–1147) in CA | [50,51,52] |
| HTLV-1 | ~94 nts | ? | ~70 nts in gag (SL1 and SL2) | [53] |
| MLV | ~476 nts | 350 nts (nt 215–565) of 5’ UTR | 328 nts | [54,55,56] |
| SNV (REV) | ~401 nts | 180 nts (SL1 and SL2) | ? | [57,58] |
| RSV | ~278 nts | 82 nts (O3 stem) | ? | [59,60,61] |
| HFV | ~100 nts | Entire UTR | ? | [34,35,36] |
| Protein | HIV-1 | HIV-2 | SIV | FIV | MPMV | MMTV | SNV/REV | MLV | RSV | BLV | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RNA | PACK | PROP | PACK | PROP | PACK | PROP | PACK | PROP | PACK | PROP | PACK | PROP | PACK | PROP | PACK | PROP | PACK | PROP | PACK | PROP |
| HIV-1 | [40,137] | [40,137] | ND | [30] | [161] | [161] | [24] | [24] | [125]* | ND | [28] | [28] | [110] | [30] | ||||||
| HIV-2 | [40,137] | [40,137] | [137] | [137] | ||||||||||||||||
| SIV | [29,137] | [29,137] | [137] | [137] | [161] | [161] | [24] | [24] | ||||||||||||
| FIV | [161] | [161] | [161] | [161] | [24] | [24] | ||||||||||||||
| MPMV | [24] | [24] | [24] | [24] | [161] | [161] | [25] | [25] | ||||||||||||
| MMTV | [125] | ND | [25] | [25] | [163] | [163] | ||||||||||||||
| SNV | [28] | [28] | [138] | [138] | ||||||||||||||||
| MLV | [110]* | ND | ND | [30] | [31,58], [121]*, [138,141] | [31,58], [121]*, [138,141] | [119]* | [119]* | [52] | ND | ||||||||||
| RSV | ||||||||||||||||||||
| BLV | ||||||||||||||||||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, L.M.; Rizvi, T.A.; Mustafa, F. Cross- and Co-Packaging of Retroviral RNAs and Their Consequences. Viruses 2016, 8, 276. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100276
Ali LM, Rizvi TA, Mustafa F. Cross- and Co-Packaging of Retroviral RNAs and Their Consequences. Viruses. 2016; 8(10):276. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100276
Chicago/Turabian StyleAli, Lizna M., Tahir A. Rizvi, and Farah Mustafa. 2016. "Cross- and Co-Packaging of Retroviral RNAs and Their Consequences" Viruses 8, no. 10: 276. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100276