
Comparative Analysis of RNAi-Based Methods to Down-Regulate Expression of Two Genes Expressed at Different Levels in Myzus persicae

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Aphid Rearing
2.2. Plant Material
2.3. Constitutive and Transient Expression of RNA Hairpins in Plants
2.4. Tobacco Rattle Virus (TRV)-Derived Constructs and Inoculation
2.5. Detection of Small Interfering RNA (siRNA) by Northern Blot and Small RNAs Purification for Aphid Acquisition
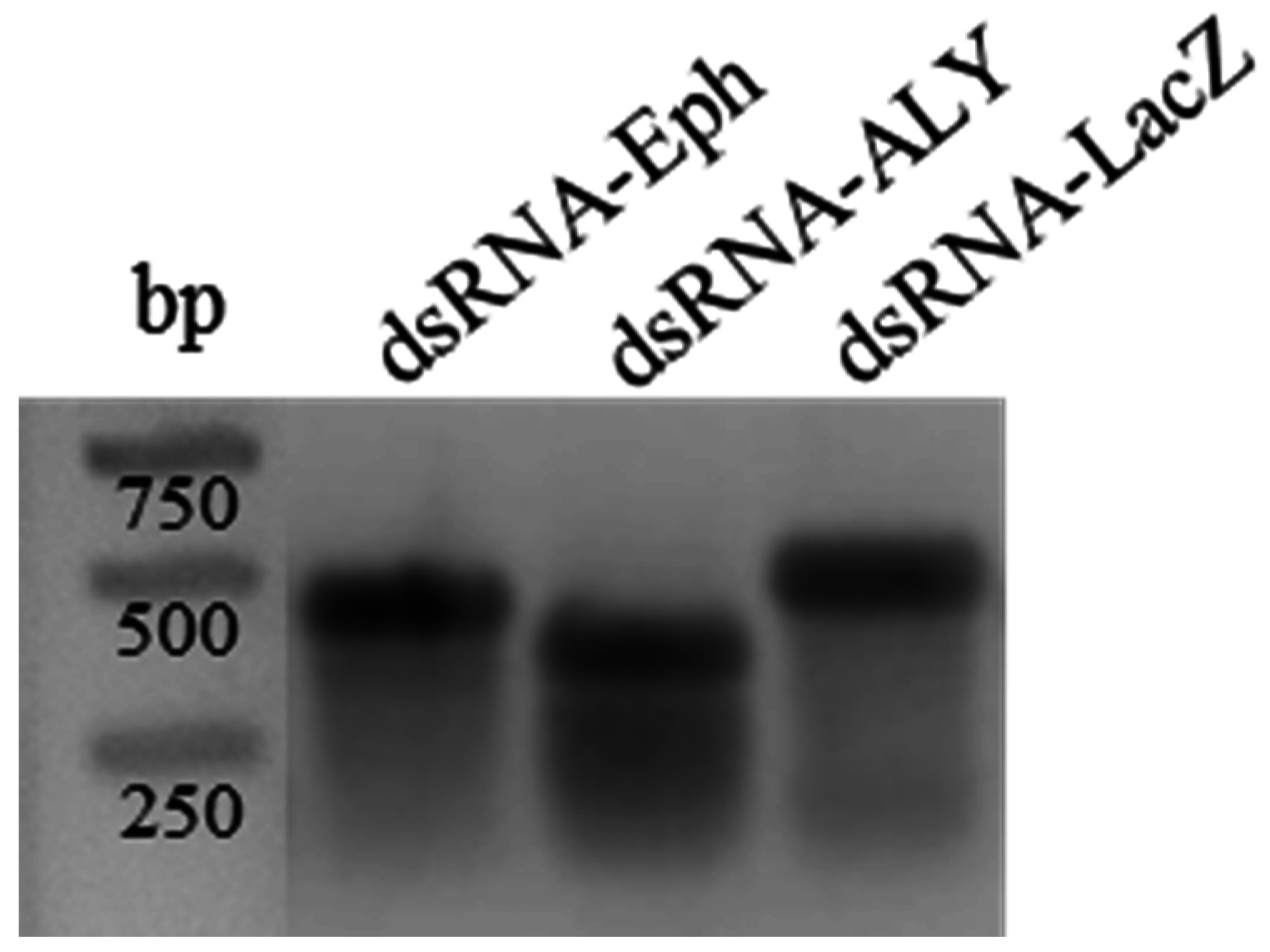
2.6. In Vitro-Synthesized double-stranded RNA (dsRNA)
2.7. dsRNA and/or siRNA Acquisition by M. persicae
2.8. RNA Extraction from Whole Aphids or Guts
2.9. Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR) and Biological Repeats
3. Results
3.1. Selection of the Candidate Genes to be Silenced in M. persicae
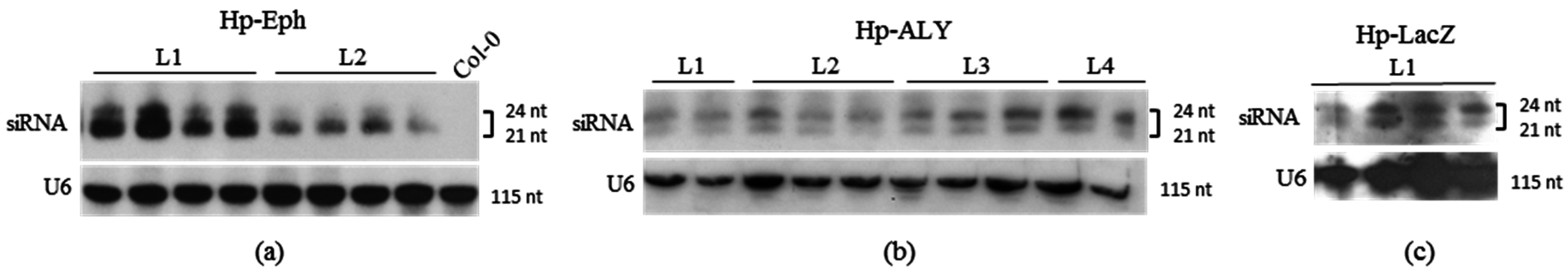
3.2. Acquisition of dsRNA or siRNA from Plants Constitutively Expressing RNA Hairpins
3.3. Acquisition of dsRNA/siRNA from Plants Expressing Transiently RNA Hairpins
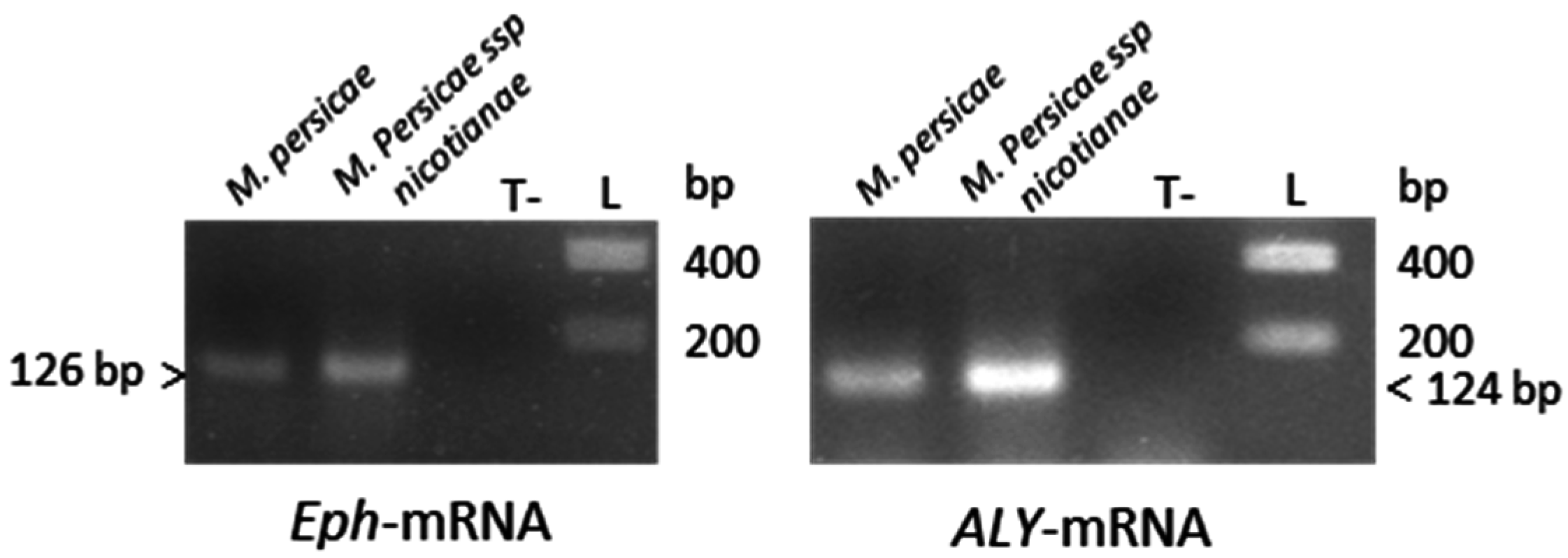
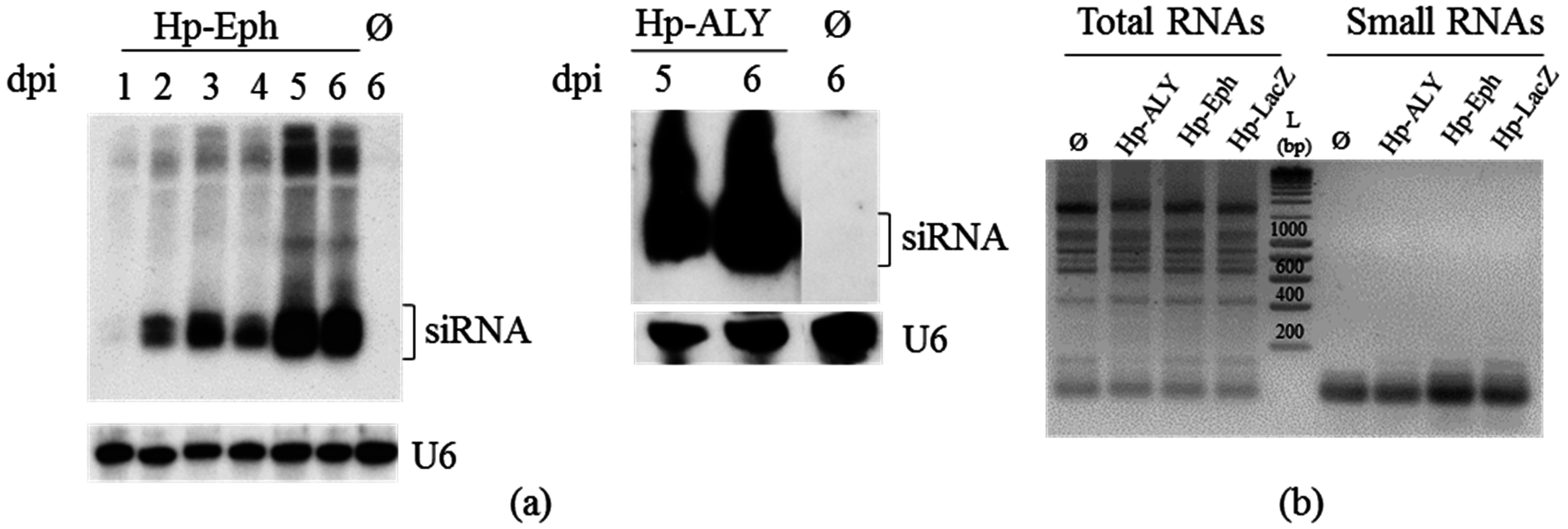
3.4. RNAi-Based Silencing by Feeding Aphids on Plants Infected with a TRV-Recombinant Virus
3.5. RNAi-Based Silencing by Artificial Feeding on In Vitro Synthesized dsRNA
4. Discussion
4.1. Oral Acquisition of dsRNA/siRNA from Transgenic Plants, from Plants Infected with Recombinant TRV, and from In Vitro-Synthesized Transcripts Are the Most Efficient Gene Silencing Methods in Aphids
4.2. Feeding Aphids on N. benthamiana Transiently Expressing Hairpin RNAs, or on Small RNAs from These Plants, Are Not Efficient Silencing Methods
4.3. Future Developments
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brault, V.; Uzest, M.; Monsion, B.; Jacquot, E.; Blanc, S. Aphids as transport devices for plant viruses. C. R. Biol. 2010, 333, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Dedryver, C.A.; Le Ralec, A.; Fabre, F. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. C. R. Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Tagu, D.; Klingler, J.P.; Moya, A.; Simon, J.C. Early progress in aphid genomics and consequences for plant-aphid interactions studies. Mol. Plant Microbe Interact. 2008, 21, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le plan ecophyto. Available online: http://agriculture.gouv.fr/ministere/le-plan-ecophyto-2018 (accessed on 12 April 2013).
- IAGC. Genome sequence of the pea aphid acyrthosiphom pisum. PLoS Biol. 2010, 8, e1000313. [Google Scholar]
- Mathers, T.C.; Chen, Y.; Kaithakottil, G.; Legeai, F.; Mugford, S.T.; Baa-Puyoulet, P.; Bretaudeau, A.; Clavijo, B.; Collela, S.; Collin, O.; et al. A clonally reproducing generalist aphid pest colonises diverse host plants by rapid transcriptional plasticity of duplicated gene clusters. bioRxiv 2016. [Google Scholar]
- Nicholson, S.J.; Nickerson, M.L.; Dean, M.; Song, Y.; Hoyt, P.R.; Rhee, H.; Kim, C.; Puterka, G.J. The genome of diuraphis noxia, a global aphid pest of small grains. BMC Genomics 2015, 16, 429. [Google Scholar] [CrossRef] [PubMed]
- Brisson, J.A.; Jacquiery, J.; Legeai, F.; Le Trionnaire, G.; Tagu, D. Genomics of phenotypic plasticity in aphids. In Management of Insects Pests to Agriculture; Czosnek, H., Ghanim, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 65–96. [Google Scholar]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.D.; Liu, Z.C.; Huang, S.L.; Chen, Z.Q.; Sun, Y.W.; Duan, P.F.; Ma, Y.Z.; Xia, L.Q. RNAi-mediated plant protection against aphids. Pest. Manag. Sci. 2016, 72, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Doudna, J.A. A three-dimensional view of the molecular machinery of RNA interference. Nature 2009, 457, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Meister, G.; Tuschl, T. Mechanisms of gene silencing by double-stranded RNA. Nature 2004, 431, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Mello, C.C.; Conte, D., Jr. Revealing the world of RNA interference. Nature 2004, 431, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Sen, G.L.; Blau, H.M. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Siomi, H.; Siomi, M.C. On the road to reading the RNA-interference code. Nature 2009, 457, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X.P.; Wang, M.Q.; Ma, W.H.; Hua, H.X. Advances in the use of the RNA interference technique in hemiptera. Insect Sci. 2013, 20, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, O.; Swevers, L.; Smagghe, G. DsRNA degradation in the pea aphid (acyrthosiphon pisum) associated with lack of response in RNAi feeding and injection assay. Peptides 2014, 53, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.H.; Yu, X.R.; Shang, Q.L.; Shi, X.Y.; Gao, X.W. Oral delivery mediated RNA interference of a carboxylesterase gene results in reduced resistance to organophosphorus insecticides in the cotton aphid, aphis gossypii glover. PLoS One 2014, 9, e102823. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Zeng, F. Feeding-based RNA interference of a gap gene is lethal to the pea aphid, acyrthosiphon pisum. PLoS ONE 2012, 7, e48718. [Google Scholar] [CrossRef] [PubMed]
- Sapountzis, P.; Duport, G.; Balmand, S.; Gaget, K.; Jaubert-Possamai, S.; Febvay, G.; Charles, H.; Rahbe, Y.; Colella, S.; Calevro, F. New insight into the RNA interference response against cathepsin-l gene in the pea aphid, acyrthosiphon pisum: Molting or gut phenotypes specifically induced by injection or feeding treatments. Insect Biochem. Mol. Biol. 2014, 51, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakesby, A.J.; Wallace, I.S.; Isaacs, H.V.; Pritchard, J.; Roberts, D.M.; Douglas, A.E. A water-specific aquaporin involved in aphid osmoregulation. Insect Biochem. Mol. Biol. 2009, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Q.; Li, X.; Sun, Y.; Wang, H.; Xia, L. Double-stranded RNA in the biological control of grain aphid (Sitobion avenae F.). Funct. Integr. Genomics. 2015, 15, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, Y.; Wang, H.; Jones, H.; Gao, Q.; Wang, D.; Ma, Y.; Xia, L. Identifying potential RNAi targets in grain aphid (sitobion avenae f.) based on transcriptome profiling of its alimentary canal after feeding on wheat plants. BMC Genomics 2013, 14, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutti, N.S.; Louis, J.; Pappan, L.K.; Pappan, K.; Begum, K.; Chen, M.S.; Park, Y.; Dittmer, N.; Marshall, J.; Reese, J.C.; et al. A protein from the salivary glands of the pea aphid, acyrthosiphon pisum, is essential in feeding on a host plant. Proc. Natl. Acad. Sci. USA 2008, 105, 9965–9969. [Google Scholar] [CrossRef] [PubMed]
- Mutti, N.S.; Park, Y.; Reese, J.C.; Reeck, G.R. RNAi knockdown of a salivary transcript leading to lethality in the pea aphid, acyrthosiphon pisum. J. Insect Sci. 2006, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.; Bhattacharya, R.; Uniyal, P.L.; Singh, R.; N.iranjan, R.S. Host generated siRNAs attenuate expression of serine protease gene in myzus persicae. PLoS One 2012, 7, e46343. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Wang, W.; Luo, L.; Chen, J.; Guo, Y.; Cui, F. Characterization of an aphid-specific, cysteine-rich protein enriched in salivary glands. Biophys. Chem. 2014, 189, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Zeng, F. Plant-mediated RNAi of a gap gene-enhanced tobacco tolerance against the myzus persicae. Transgenic Res. 2014, 23, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Pitino, M.; Coleman, A.D.; Maffei, M.E.; Ridout, C.J.; Hogenhout, S.A. Silencing of aphid genes by dsRNA feeding from plants. PLoS One 2011, 6, e25709. [Google Scholar] [CrossRef] [PubMed]
- Pitino, M.; Hogenhout, S.A. Aphid protein effectors promote aphid colonization in a plant species-specific manner. Mol. Plant. Microbe Interact 2013, 26, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Duan, X.; Lv, Y.; Zhang, X.; Nie, Z.; Xie, C.; Ni, Z.; Liang, R. Silencing of an aphid carboxylesterase gene by use of plant-mediated RNAi impairs sitobion avenae tolerance of phoxim insecticides. Transgenic Res. 2014, 23, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Jaubert-Possamai, S.; Le Trionnaire, G.; Bonhomme, J.; Christophides, G.K.; Rispe, C.; Tagu, D. Gene knockdown by RNAi in the pea aphid acyrthosiphon pisum. BMC Biotechnol. 2007, 7, 63. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Lu, Y.H.; Shang, Q.L.; Song, D.L.; Gao, X.W. Gene silencing of two acetylcholinesterases reveals their cholinergic and non-cholinergic functions in rhopalosiphum padi and sitobion avenae. Pest. Manag. Sci. 2015, 71, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, Z. Peroxiredoxin 1 protects the pea aphid acyrthosiphon pisum from oxidative stress induced by micrococcus luteus infection. J. Invertebr. Pathol. 2015, 127, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.D.; Wouters, R.H.; Mugford, S.T.; Hogenhout, S.A. Persistence and transgenerational effect of plant-mediated RNAi in aphids. J. Exp. Bot. 2015, 66, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Monsion, B.; Mulot, M.; Rastegar, M.; Boissinot, S.; Erdinger, M.; Bochet, N.; Brault, V. Identification of an aphid protein involved in turnip yellows virus transmission by myzus persicae. In preparation.
- Zhou, Z.; Luo, M.J.; Straesser, K.; Katahira, J.; Hurt, E.; Reed, R. The protein aly links pre-messenger-RNA splicing to nuclear export in metazoans. Nature 2000, 407, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Perez White, B.E.; Getsios, S. Eph receptor and ephrin function in breast, gut, and skin epithelia. Cell. Adh. Migr. 2014, 8, 327–338. [Google Scholar] [CrossRef] [PubMed]
- E-RNAi. Available online: http://www.e-rnai.org (accessed on 5 May 2010).
- Martinez-Trujillo, M.; Limones-Briones, V.; Cabrera-Ponce, J.L.; Herrera-Estrella, L. Improving transformation efficiency of arabidopsis thaliana by modifying the floral dip method. Plant. Mol. Biol. Report. 2004, 22, 63–70. [Google Scholar] [CrossRef]
- English, J.J.; Davenport, G.F.; Elmayan, T.; Vaucheret, H.; Baulcombe, D.C. Requirement of sense transcription for homology-dependent virus resistance and trans-inactivation. Plant J. 1997, 12, 597–603. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Bruyère, A.; Brault, V.; Ziegler-Graff, V.; Simonis, M.T.; Van den Heuvel, J.F.; Richards, K.; Guilley, H.; Jonard, G.; Herrbach, E. Effects of mutations in the beet western yellows virus readthrough protein on its expression and packaging and on virus accumulation, symptoms, and aphid transmission. Virology 1997, 230, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Pitulescu, M.E.; Adams, R.H. Eph/ephrin molecules--a hub for signaling and endocytosis. Genes Dev. 2010, 24, 2480–2492. [Google Scholar] [CrossRef] [PubMed]
- Stutz, F.; Bachi, A.; Doerks, T.; Braun, I.C.; Seraphin, B.; Wilm, M.; Bork, P.; Izaurralde, E. Ref, an evolutionary conserved family of hnrnp-like proteins, interacts with tap/mex67p and participates in mRNA nuclear export. RNA 2000, 6, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Aphidbase. Available online: http://www.aphidbase.com/ (accessed on 6 May 2010).
- Abdellatef, E.; Will, T.; Koch, A.; Imani, J.; Vilcinskas, A.; Kogel, K.H. Silencing the expression of the salivary sheath protein causes transgenerational feeding suppression in the aphid sitobion avenae. Plant Biotechnol. J. 2015, 13, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Song, X.; Wang, G.; Yang, K.; Wang, Y.; Niu, L.; Chen, X.; Fang, R. Plant-generated artificial small RNAs mediated aphid resistance. PLoS ONE 2014, 9, e97410. [Google Scholar] [CrossRef] [PubMed]
- Tzin, V.; Yang, X.; Jing, X.; Zhang, K.; Jander, G.; Douglas, A.E. Rna interference against gut osmoregulatory genes in phloem-feeding insects. J. Insect Physiol. 2015, 79, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Purkayastha, A.; Dasgupta, I. Virus-induced gene silencing: A versatile tool for discovery of gene functions in plants. Plant. Physiol. Biochem. 2009, 47, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, F.; Martin-Hernandez, A.M.; Baulcombe, D.C. Technical advance. Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001, 25, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Jia, L.; Goggin, F. The reliability of virus-induced gene silencing experiments using tobacco rattle virus in tomato is influenced by the size of the vector control. Mol. Plant Pathol. 2011, 12, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Cilia, M.; Tamborindeguy, C.; Fish, T.; Howe, K.; Thannhauser, T.W.; Gray, S. Genetics coupled to quantitative intact proteomics links heritable aphid and endosymbiont protein expression to circulative polerovirus transmission. J. Virol. 2011, 85, 2148–2166. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cox-Foster, D.; Gray, S.M.; Gildow, F. Vector specificity of barley yellow dwarf virus (bydv) transmission: Identification of potential cellular receptors binding bydv-mav in the aphid, sitobion avenae. Virology 2001, 286, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Seddas, P.; Boissinot, S.; Strub, J.M.; Van Dorsselaer, A.; Van Regenmortel, M.H.; Pattus, F. Rack-1, gapdh3, and actin: Proteins of myzus persicae potentially involved in the transcytosis of beet western yellows virus particles in the aphid. Virology 2004, 325, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Thannhauser, T.W.; Burrows, M.; Cox-Foster, D.; Gildow, F.E.; Gray, S.M. Coupling genetics and proteomics to identify aphid proteins associated with vector-specific transmission of polerovirus (luteoviridae). J. Virol. 2008, 82, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Linz, L.B.; Liu, S.; Chougule, N.P.; Bonning, B.C. In vitro evidence supports membrane alanyl aminopeptidase n as a receptor for a plant virus in the pea aphid vector. J. Virol. 2015, 89, 11203–11212. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sivakumar, S.; Sparks, W.O.; Miller, W.A.; Bonning, B.C. A peptide that binds the pea aphid gut impedes entry of pea enation mosaic virus into the aphid hemocoel. Virology 2010, 401, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Ramaseshadri, P.; Anderson, J.; Bachman, P.; Clinton, W.; Flannagan, R.; Ilagan, O.; Lawrence, C.; Levine, S.; Moar, W.; et al. Characterizing the mechanism of action of double-stranded RNA activity against western corn rootworm (diabrotica virgifera virgifera leconte). PLoS ONE 2012, 7, e47534. [Google Scholar] [CrossRef] [PubMed]
- Jaubert-Possamai, S.; Rispe, C.; Tanguy, S.; Gordon, K.; Walsh, T.; Edwards, O.; Tagu, D. Expansion of the miRNA pathway in the hemipteran insect acyrthosiphon pisum. Mol. Biol. Evol. 2010, 27, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.H.; Waterhouse, P.M. Rnai for insect-proof plants. Nat. Biotechnol. 2007, 25, 1231–1232. [Google Scholar] [CrossRef] [PubMed]
- Tomoyasu, Y.; Miller, S.C.; Tomita, S.; Schoppmeier, M.; Grossmann, D.; Bucher, G. Exploring systemic RNA interference in insects: A genome-wide survey for RNAi genes in tribolium. Genome Biol. 2008, 9, R10. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.M.; Ashfaq, M.; Khan, A.A.; Rasool, A.; Iqbal, J.; Mansoor, S. Inoculation of nicotiana tabacum with recombinant potato virus x induces RNA interference in the solenopsis mealybug, phenacoccus solenopsis tinsley (hemiptera: Pseudococcidae). Biotechnol. Lett. 2015, 37, 2083–2090. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.M.; Ashfaq, M.; Kiss, Z.; Khan, A.A.; Mansoor, S.; Falk, B.W. Use of recombinant tobacco mosaic virus to achieve RNA interference in plants against the citrus mealybug, planococcus citri (hemiptera: Pseudococcidae). PLoS ONE 2013, 8, e73657. [Google Scholar] [CrossRef] [PubMed]
- Wuriyanghan, H.; Falk, B.W. Rna interference towards the potato psyllid, bactericera cockerelli, is induced in plants infected with recombinant tobacco mosaic virus (TMV). PLoS ONE 2013, 8, e66050. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.; Wang, H.; Pallett, D.; Dalmay, T. Evidence for targeting common siRNA hotspots and gc preference by plant dicer-like proteins. FEBS Lett. 2007, 581, 3267–3272. [Google Scholar] [CrossRef] [PubMed]
- Buhtz, A.; Springer, F.; Chappell, L.; Baulcombe, D.C.; Kehr, J. Identification and characterization of small RNAs from the phloem of brassica napus. Plant J. 2008, 53, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Kehr, J.; Buhtz, A. Long distance transport and movement of RNA through the phloem. J. Exp. Bot. 2008, 59, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.A.; Bitra, K.; Zhang, S.; Wang, L.; Lynn, D.E.; Strand, M.R. The uga-cie1 cell line from chrysodeixis includens exhibits characteristics of granulocytes and is permissive to infection by two viruses. Insect Biochem. Mol. Biol. 2010, 40, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Christiaens, O.; Liu, J.; Niu, J.; Cappelle, K.; Caccia, S.; Huvenne, H.; Smagghe, G. Delivery of dsRNA for RNAi in insects: An overview and future directions. Insect Sci. 2013, 20, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the african malaria mosquito (anopheles gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Tagu, D.; Le Trionnaire, G.; Tanguy, S.; Gauthier, J.P.; Huynh, J.R. Ems mutagenesis in the pea aphid acyrthosiphon pisum. G3 2014, 4, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Van Munster, M.; Dullemans, A.M.; Verbeek, M.; van den Heuvel, J.F.; Reinbold, C.; Brault, V.; Clerivet, A.; van der Wilk, F. Characterization of a new densovirus infecting the green peach aphid myzus persicae. J. Invertebr. Pathol. 2003, 84, 6–14. [Google Scholar] [CrossRef]
- Van Munster, M.; Dullemans, A.M.; Verbeek, M.; van den Heuvel, J.F.; Reinbold, C.; Brault, V.; Clerivet, A.; van der Wilk, F. A new virus infecting myzus persicae has a genome organization similar to the species of the genus densovirus. J. Gen. Virol. 2003, 84, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Liu, M.; Deng, Y.; Peng, H.; Chen, X. Development of an efficient recombinant mosquito densovirus-mediated RNA interference system and its preliminary application in mosquito control. PLoS ONE 2011, 6, e21329. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aphid Feeding Source | Exp. | Plant or RNA Concentration | AAP b | Relative Fold Change Eph-mRNA in Whole M. persicae c | |
|---|---|---|---|---|---|
| A | Transgenic A. thaliana expressing Hp-Eph a | 1 | Ara:Hp-Eph (T1) line 1 | 10 d | 0.248 ± 0.037 (−75.2%) d,* |
| Ara:Hp-Eph (T1) line 2 | 0.873 ± 0.103 (−12.7%) d | ||||
| 2 | Ara:Hp-Eph (T1) line 1 | 10 d | 0.140 ± 0.065 (−86.0%) d,* | ||
| Ara:Hp-Eph (T1) line 2 | 0.533 ± 0.089 (−46.7%) d,* | ||||
| Ara:Hp-Eph (T1) line 2 | 0.428 ± 0.145 (−57.2%) d,* | ||||
| 3 | Ara:Hp-Eph (T3) | 10 d | 0.818 ± 0.032 (−18.2%) * | ||
| 1.032 ± 0.024 (+3.2%) | |||||
| 4 | Ara:Hp-Eph (T3) | 10 d | 1.048 ± 0.028 (+4.8%) | ||
| 1.027 ± 0.060 (−2.8%) | |||||
| 5 | Ara:Hp-Eph (T3) | 10 d | 0.843 ± 0.049 (−15.7%) * | ||
| 1.065 ± 0.055 (+6.5%) | |||||
| 0.914 ± 0.43 (−8.6%) * | |||||
| 6 | Ara:Hp-Eph (T3) | 13 d | 0.882 ± 0.078 (−11.8%) | ||
| 1.120 ± 0.068 (+12.0%) | |||||
| 0.726 ± 0.034 (−27.3%) * | |||||
| B | N. benthamiana transiently expressing Hp-Eph | 1 | Bentha:Hp-Eph | 10 d | unstable reference genes f |
| C | N. benthamiana infected with TRV-Eph | 1 | Bentha:TRV-Eph | 7 d | 0.581 ± 0.018 (−41.9%) *,e |
| 2 | 0.581 ± 0.012 (−41.9%) * | ||||
| D | siRNA purified from N. benthamiana transiently expressing Hp-Eph | 1 | siRNA 60 ng/µL | 24 h | 1.143 ± 0.054 (+14.3%) |
| 2 | siRNA 120 ng/µL | 24 h | 0.856 ± 0.042 (−14.4%) NA | ||
| 0.832 ± 0.048 (−16.8%) NA | |||||
| 3 | siRNA 70 ng/µL | 36 h | 0.936 ± 0.077 (−6.3%) | ||
| 4 | siRNA 100 ng/µL | 60 h | 1.056 ± 0.060 (+5.6%) | ||
| 5 | siRNA 100 ng/µL | 72 h | unstable reference genes | ||
| E | In vitro-synthesized dsRNA-Eph | 1 | dsRNA 100 ng/µL | 72 h | 0.902 ± 0.013 (−9.8%) * |
| 2 | 0.917 ± 0.092 (−8.3%) |
| Aphid Feeding Source | Exp. | Plant or RNA Concentration | AAP b | Relative Fold Change Eph-mRNA in M. persicae Guts c | |
|---|---|---|---|---|---|
| A | Transgenic A. thaliana expressing Hp-Eph a | 1 | Ara:Hp-Eph (T3) Ara:Hp-Eph (T4) | 7 d | 0.126 ± 0.021 (−87.4%) * |
| 2 | 0.928 ± 0.062 (−7.8%) | ||||
| B | N. benthamiana transiently expressing Hp-Eph | 1 | Bentha:Hp-Eph | 10 d | 1.038 ± 0.037 (+3.8%) |
| C | N. benthamiana infected with TRV-Eph | 1 | Bentha:TRV-Eph | 7 d | 0.812 ± 0.013 (−18.8%) * |
| 2 | 0.441 ± 0.037 (−55.9%) * | ||||
| D | siRNA purified from N. benthamiana transiently expressing Hp-Eph | 1 | siRNA 100 ng/µL | 72 h | unstable reference gene d |
| E | In vitro-synthesized dsRNA-Eph | 1 | dsRNA 200 ng/µL | 72 h | 0.470 ± 0.021 (−53.0%) * |
| 2 | dsRNA 400 ng/µL | 0.153 ± 0.015 (−84.7%) * |
| Aphid Feeding Source | Exp. | Plant or RNA Concentration | AAP b | Relative Fold Change ALY-mRNA in Whole M. persicae c | |
|---|---|---|---|---|---|
| A | Transgenic A. thaliana expressing Hp-ALY a | 1 | Ara:Hp-ALY (T1) | 10 d | 0.862 ± 0.105 (−13.8%) d * |
| 2 | 1.177 ± 0.023 (+17.7%) d * | ||||
| 3 | 1.040 ± 0.026 (+4.0%) d | ||||
| 4 | 1.079 ± 0.161 (+7.9%) d | ||||
| B | N. benthamiana transiently expressing Hp-ALY | 1 | Bentha:Hp-ALY | 10 d | unstable reference genes e |
| D | N. benthamiana infected with TRV-ALY | 1 | Bentha:TRV-ALY | 7 d | 0.881 ± 0.033 (−11.9%) |
| 2 | 0.847 ± 0.033 (−15.3%) * | ||||
| C | siRNA purified from N. benthamiana transiently expressing Hp-ALY | 1 | siRNA 100 ng/µL | 72 h | unstable reference genes |
| E | In vitro synthesized dsRNA-ALY | 1 | dsRNA 400 ng/µL | 72 h | 1.041 ± 0.037(+4.0%) |
| Aphid Feeding Source | Exp. | Plant or RNA Concentration | AAP b | Relative Fold Change ALY-mRNA in M. persicae Guts c | |
|---|---|---|---|---|---|
| A | Transgenic A. thaliana expressing Hp-ALY a | 1 | Ara:Hp-ALY (T3) | 13 d | 1.315 ± 0.055 (+31.5%) * |
| 2 | 13 d | 1.062 ± 0.007 (+6,2%) * | |||
| B | N. benthamiana transiently expressing Hp-ALY | 1 | Bentha:Hp-ALY | 10 d | 1.073 ± 0.051 (+7.3%) |
| C | N. benthamiana infected with TRV-ALY | 1 | Bentha:Hp-ALY | 7 d | 1.136 ± 0.036 (+13.6%) * |
| 2 | 0.915 ± 0.023 (−8.5%) * | ||||
| D | siRNA purified from N. benthamiana transiently expressing Hp-ALY | 1 | siRNA 100 ng/µL | 72 h | unstable reference genes d |
| E | In vitro-synthesized dsRNA-ALY | 1 | dsRNA 400 ng/µL | 72 h | 0.263 ± 0.023 (−73.7%) * |
| 2 | 0.942 ± 0.039 (−5.8%) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulot, M.; Boissinot, S.; Monsion, B.; Rastegar, M.; Clavijo, G.; Halter, D.; Bochet, N.; Erdinger, M.; Brault, V. Comparative Analysis of RNAi-Based Methods to Down-Regulate Expression of Two Genes Expressed at Different Levels in Myzus persicae. Viruses 2016, 8, 316. https://0-doi-org.brum.beds.ac.uk/10.3390/v8110316
Mulot M, Boissinot S, Monsion B, Rastegar M, Clavijo G, Halter D, Bochet N, Erdinger M, Brault V. Comparative Analysis of RNAi-Based Methods to Down-Regulate Expression of Two Genes Expressed at Different Levels in Myzus persicae. Viruses. 2016; 8(11):316. https://0-doi-org.brum.beds.ac.uk/10.3390/v8110316
Chicago/Turabian StyleMulot, Michaël, Sylvaine Boissinot, Baptiste Monsion, Maryam Rastegar, Gabriel Clavijo, David Halter, Nicole Bochet, Monique Erdinger, and Véronique Brault. 2016. "Comparative Analysis of RNAi-Based Methods to Down-Regulate Expression of Two Genes Expressed at Different Levels in Myzus persicae" Viruses 8, no. 11: 316. https://0-doi-org.brum.beds.ac.uk/10.3390/v8110316