Multiple Genome Wide Association Mapping Models Identify Quantitative Trait Nucleotides for Brown Planthopper (Nilaparvata lugens) Resistance in MAGIC Indica Population of Rice

 , ,
, ,  ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
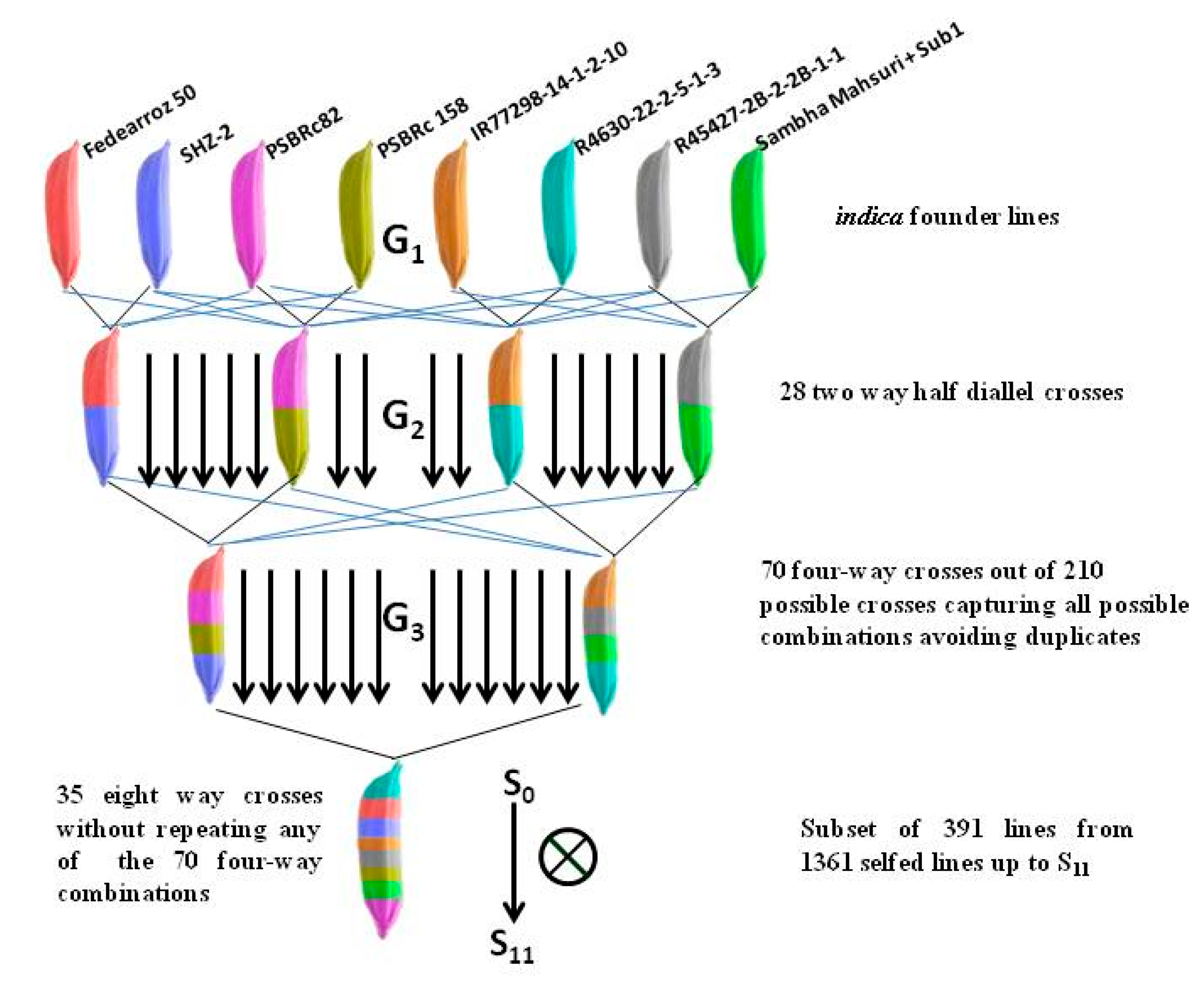
2.1. Plant Material
2.2. Phenotyping, Scoring and Data Analysis
2.3. Genotyping and SNP Calling in GBS Pipeline
2.4. Genome-Wide Association Analysis
3. Results and Discussion
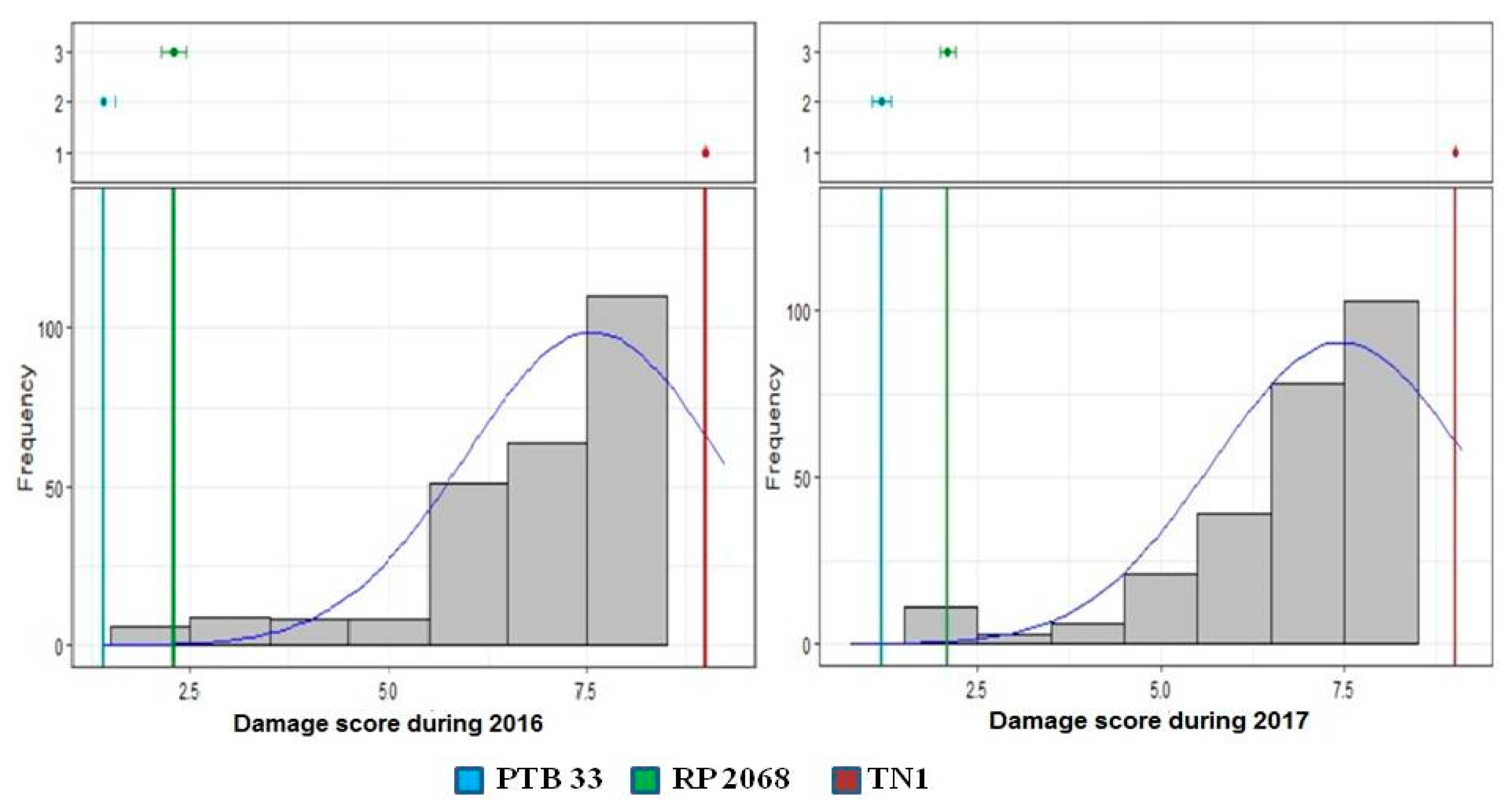
3.1. Response to Infestation
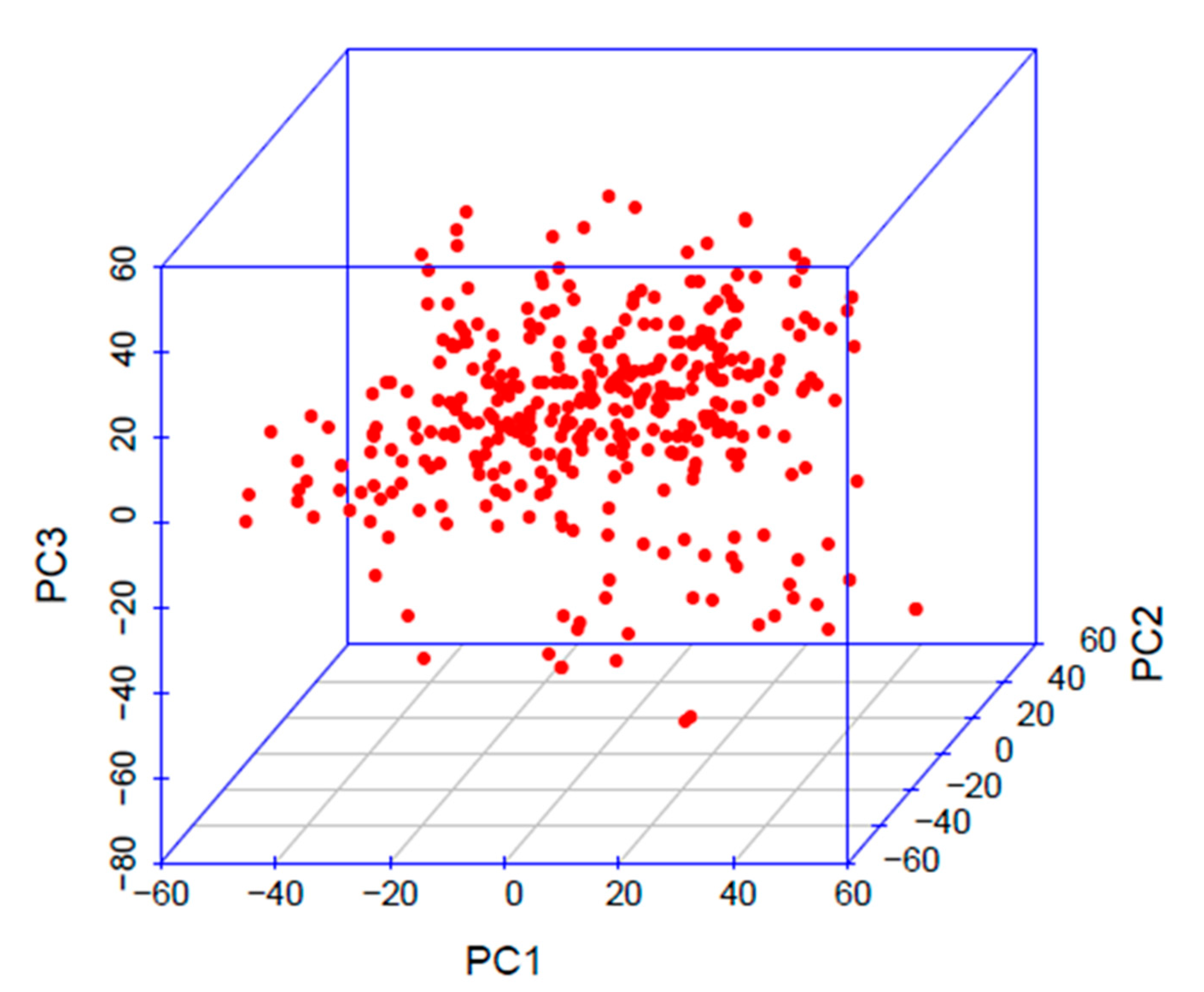
3.2. Marker and Covariates
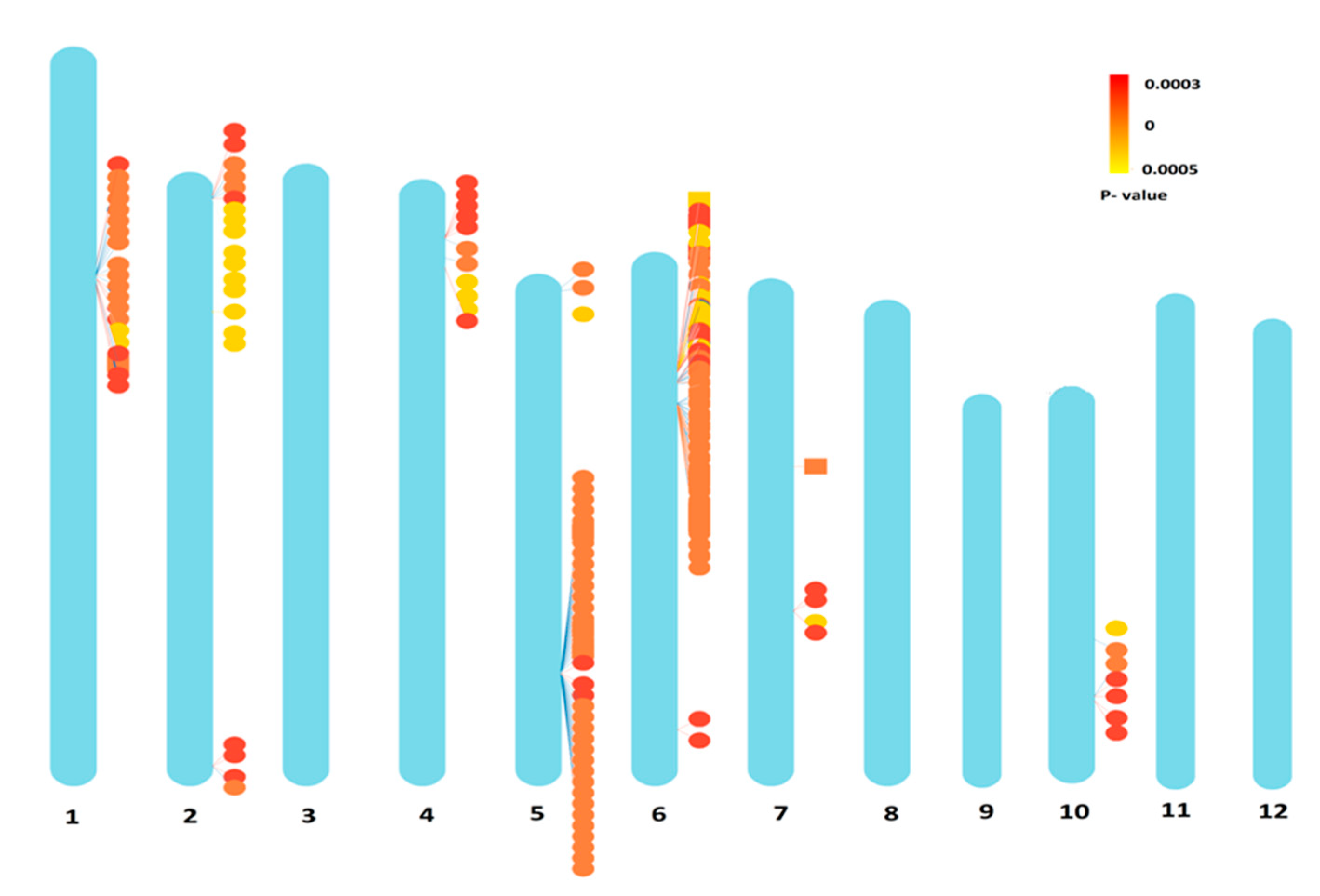
3.3. Identification of SNPs
3.4. Identification for Candidate Genes for Resistance Based on SNPs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Suenaga, H.; Nakatsuka, K. Studies on forecasting of the occurrence of leafhoppers in the paddy field. In The First Special Research Report on the Disease and Insect Forecasting; Promotion Office, Ministry of Agriculture and Forestry: Tokyo, Japan, 1958; p. 468. [Google Scholar]
- Koya, V.K.M. Brown hopper, the destructive pest. Kurukshetra 1974, 22, 13. [Google Scholar]
- Nalinakumari, T.; Mammen, K.V. Biology of the brown planthopper Nilaparvata lugens (Stål.) (Delphacidae, Hemiptera). Agric. Res. J. Kerala 1975, 13, 53–54. [Google Scholar]
- Jena, M.; Adak, T.; Sahu, R.K.; Somnath, S.; Pokhare; Berliner, J. Integrated management of Brown PlantHopper (BPH). Published by The Director, for the Central Rice Research Institute, Cuttak, India. CRRI Technol. Bull. 2015, 111. [Google Scholar]
- Catindig, J.L.A.; Arida, G.S.; Baehaki, S.E.; Bentur, J.S.; Cuong, L.Q.; Norowi, M. Situation of planthoppers in Asia. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Baños, CA, USA, 2009; pp. 191–220. [Google Scholar]
- Cabauatan, P.Q.; Cabunagan, R.C.; Ryong Choi, I. Rice viruses transmitted by the brown planthopper Nilaparvata lugens Stål. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 357–368. [Google Scholar]
- Pan, Y.; Huang, L.; Song, S.; Hu, M.; Chang, S.; Lv, Q.; Li, Y.; Wang, T.; Ouyang, X.; Fu, X. Identification of brown planthopper resistance gene Bph32 in the progeny of a rice dominant genic male sterile recurrent population using genome-wide association study and RNA-seq analysis. Mol. Breed. 2019, 39, 72. [Google Scholar] [CrossRef]
- Khush, G.S.; Karim, A.N.; Angeles, E.R. Genetics of resistance of rice cultivar ARC 10550 to Bangladesh brown planthopper biotype. J. Genet. 1985, 64, 121–125. [Google Scholar] [CrossRef]
- Brar, D.S.; Virk, P.S.; Jena, K.K.; Khush, G.S. Breeding for resistance to planthoppers in rice. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 401–428. [Google Scholar]
- Pathak, M.D.; Khush, G.S. Studies of varietal resistance in Rice to the Brown planthopper at the International Rice Research Institute. In Brown Planthopper: Threat to Rice Production in Asia; International Rice Research Institute: Manila, Philippines, 1979; pp. 285–301. [Google Scholar]
- Prahalada, G.D.; Shivakumar, N.; Lohithaswa, H.C.; Sidde Gowda, D.K.; Ramkumar, G.; Kim, S.R.; Ramachandra, C.; Hittalmani, S.; Mohapatra, T.; Jena, K.K. Identification and fine mapping of a new gene, BPH31 conferring resistance to brown planthopper biotype 4 of India to improve rice, Oryza sativa L. Rice 2017, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Balachiranjeevi, C.H.; Prahalada, G.D.; Mahender, A.; Jamaloddin, M.D.; Sevilla, M.A.L.; Marfori-Nazarea, C.M. Identification of a novel locus, BPH38(t), conferring resistance to brown planthopper (Nilaparvata lugens Stal.) using early backcross population in rice (Oryza sativa L.). Euphytica 2019, 215. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Cheng, L.; Yan, L. Mapping and characterization of a quantitative trait locus resistance to the brown planthopper in the rice variety IR64. Hereditas 2019, 156, 22. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Sarao, P.S.; Bhatia, D.; Neelam, K.; Kaur, A.; Mangat, G.S. High-resolution genetic mapping of a novel brown planthopper resistance locus, Bph34 in Oryza sativa L X Oryza nivara (Sharma & Shastry) derived inter-specific F2 population. Theor. Appl. Genet. 2018, 131, 1163–1171. [Google Scholar] [CrossRef]
- Naik, S.B.; Divya, D.; Sahu, N.; Sundaram, R.M.; Sarao, P.S.; Singh, K. A new gene Bph33(t) conferring resistance to brown planthopper (BPH), Nilaparvata lugens (Stal) in rice line RP2068-18-3-5. Euphytica 2018, 214, 53. [Google Scholar] [CrossRef]
- Guo, J.P.; Xu, C.X.; Wu, D.; Zhao, Y.; Qiu, Y.; Wang, X. Bph6 encodes an exocyst localized protein and confers broad resistance to planthoppers in rice. Nat. Genet. 2018, 50, 297–306. [Google Scholar] [CrossRef]
- Jena, K.K.; Kim, S.M. Current status of brown planthopper (BPH) resistance and genetics. Rice 2010, 3, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Kover, P.X.; Valdar, W.; Trakalo, J.; Scarcelli, N.; Ehrenreich, I.M.; Purugganan, M.D. A multi parent advanced generation inter-cross to fine-map quantitative traits in Arabidopsis thaliana. PLoS Genet. 2009, 5, e1000551. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.E.; Verbyla, K.L.; Verbyla, A.P.; Raghavan, C.; Singh, V.K.; Gaur, P. MAGIC populations in crops: Current status and future prospects. Theor. Appl. Genet. 2015, 128, 999–1017. [Google Scholar] [CrossRef]
- Zaw, H.; Raghavan, C.; Pocsedio, A.; Swamy, B.P.M.; Jubay, M.L.; Singh, R.K. Exploring genetic architecture of grain yield and quality traits in a 16-way indica by japonica rice MAGIC global population. Sci. Rep. 2019, 9, 19605. [Google Scholar] [CrossRef]
- Bandillo, N.; Raghavan, C.; Muyco, P.A.; Sevilla, M.A.L.; Lobina, I.T.; Ermita, C.J.D. Multi-parent advanced generation inter-cross (MAGIC) populations in rice: Progress and potential for genetics research and breeding. Rice 2013, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, C.; Mauleon, R.; Lacorte, V.; Jubay, M.; Zaw, H.; Bonifacio, J. Approaches in Characterizing Genetic Structure and Mapping in a Rice Multi parental Population. G3 Bethesda 2017, 7, 1721–1730. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, E.A.; Medrano, F.G.; Rapusas, H.R. Genetic Evaluation for Insect Resistance in Rice; IRRI: Los Baños, Philippines, 1985. [Google Scholar]
- Velusamy, R.; Heinrichs, E.A.; Medrano, F.G. Greenhouse techniques to identify field resistance to the brown plant hopper, Nilaparvata lugens (Stål) (Homoptera: Delphacidae), in rice cultivars. Crop Prot. 1986, 5, 328–333. [Google Scholar] [CrossRef]
- Horgan, F.G. Mechanisms of resistance: A major gap in understanding planthopper-rice interactions. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 281–302. [Google Scholar]
- IRRI. Standard Evaluation System (SES) for Rice, 5th ed.; International Rice Research Institute: Manila, Philipines, 2013; pp. 38–39. [Google Scholar]
- Federer, W.T. Augmented designs with one-way elimination of heterogeneity. Biometrics 1961, 17, 447–473. [Google Scholar] [CrossRef]
- Aravind, J.; MukeshSankar, S.; Wankhede, D.P.; Kaur, V. Augmented RCBD: Analysis of Augmented Randomised Complete Block Designs. R Package Version 0.1.2.9000. 2020. Available online: https://www.aravind-j.github.io/augmentedRCBD/https://cran.r-project.org/package=augmentedRCBD (accessed on 3 December 2019).
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef]
- Lipka, A.E.; Tian, F.; Wang, Q.S.; Peiffer, J.; Li, M.; Bradbury, P.J. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [Green Version]
- Bossa-Castro, A.M.; Tekete, C.; Raghavan, C.; Delorean, E.E.; Dereeper, A.; Dagno, K. Allelic variation for broad-spectrum resistance and susceptibility to bacterial pathogens identified in a rice MAGIC population. Plant Biotechnol. J. 2016, 16, 1559–1568. [Google Scholar] [CrossRef] [Green Version]
- Price, A.L.; Patterson, N.J.; Plenge, R.M.; Weinblatt, M.E.; Shadick, N.A.; Reich, D. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 2006, 38, 904–909. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Mao, Y.; Xie, C.; Smith, H.; Luo, L.; Xu, S. Mapping quantitative trait loci using naturally occurring genetic variance among commercial inbred lines of maize (Zea mays L.). Genetics 2005, 169, 2267–2275. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.M.; Pressoir, G.; Briggs, W.H.; Bi, I.V.; Yamasaki, M.; Doebley, J.F. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Ersoz, E.; Lai, C.Q.; Todhunter, R.J.; Tiwari, H.K.; Gore, M.A. Mixed linear model approach adapted for genome-wide association studies. Nat. Genet. 2010, 42, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Tian, F.; Pan, Y.; Buckler, E.S.; Zhang, Z. A super powerful method for genome wide association study. PLoS ONE 2014, 9, e107684. [Google Scholar] [CrossRef]
- Segura, V.; Vilhjalmsson, B.J.; Platt, A.; Korte, A.; Seren, U.; Long, Q. An efficient multi-locus mixed-model approach for genome-wide association studies in structured populations. Nat. Genet. 2012, 44, 825–830. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.X.; Weng, Q.M.; Ren, X.; Huang, Z.; Zhu, L.L.; He, G.C. Two white backed planthopper resistance genes in rice share the same loci with those for brown planthopper resistance. Heredity 2004, 92, 212–217. [Google Scholar] [CrossRef]
- Zhang, X.; Mao, K.; Liao, X.; He, B.; Jin, R.; Tang, T.; Wan, H.; Li, J. Fitness cost of nitenpyram resistance in the brown planthopper Nilaparvata Lugens. J. Pest Sci. 2018, 91, 1145–1151. [Google Scholar] [CrossRef]
- Zhu, P.; Zheng, X.; Xu, H.; Johnson, A.C.; Heong, K.L.; Gurr, G.M.; Lu, Z. Nitrogen fertilization of rice plants improves ecological fitness of an entomophagous predator but dampens its impact on prey, the rice brown planthopper, Nilaparvata lugens. J. Pest Sci. 2020, 93, 747–755. [Google Scholar] [CrossRef]
- Wu, S.F.; Zeng, B.; Zheng, C.; Mu, X.C.; Zhang, Y.; Hu, J. The evolution of insecticide resistance in the brown planthopper (Nilaparvata lugens Stål) of China in the period 2012–2016. Sci. Rep. 2018, 8, 4586. [Google Scholar] [CrossRef]
- Du, B.; Zhang, W.L.; Liu, B.F.; Hu, J.; Wei, Z.; Shi, Z.Y. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc Natl Acad. Sci. USA 2009, 106, 22163–22168. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Chen, R.; Guo, J.; He, G. Current understanding of the genomic, genetic, and molecular control of insect resistance in rice. Mol. Breed. 2020, 40, 24. [Google Scholar] [CrossRef] [Green Version]
- Fujita, D.; Kohli, A.; Horgan, F.G. Rice resistance to planthoppers and leaf hoppers. Crit. Rev. Plant Sci. 2013, 32, 162–191. [Google Scholar] [CrossRef]
- Hu, J.; Xiao, C.; He, Y.Q. Recent progress on the genetics and molecular breeding of brown planthopper resistance in rice. Rice 2016, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Jing, S.; Zhao, Y.; Du, B.; Chen, R.; Zhu, L.; He, G. Genomics of interaction between the brown planthopper and rice. Cur. Opin. Insect Sci. 2017, 19, 82–87. [Google Scholar] [CrossRef]
- Jiang, H.C.; Hu, J.; Li, Z.; Liu, J.; Gao, G.J.; Zhang, Q.L. Evaluation and breeding application of six brown planthopper resistance genes in rice maintainer line Jin 23B. Rice 2018, 11, 22. [Google Scholar] [CrossRef]
- Horgan, F.G.; Ramal, A.F.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S. Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia against resistant rice varieties. Crop. Prot. 2015, 78, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Sogawa, K. Planthopper outbreaks in different paddy ecosystems in Asia: Man-made hopper plagues that threatened the green revolution in rice. In Rice Planthoppers: Ecology, Management, Socioeconomics and Policy; Heong, K.L., Ed.; Zhejiang University Press: Hangzhou, China; Hangzhou and Springer Science: Hangzhou, China; Business Media Dordrecht: Berlin, Germany, 2015. [Google Scholar] [CrossRef]
- Deen, R.; Ramesh, K.; Gautam, S.K.; Rao, Y.K.; Lakshmi, V.J.; Viraktamath, B.C. Identification of new gene for BPH resistance introgressed from O. rufipogon. Rice Genet. Newsl. 2010, 25, 70–72. [Google Scholar]
- Leung, H.; Raghavan, C.; Zhou, B.; Oliva, R.; Choi, I.R.; Lacorte, V. Allele mining and enhanced genetic recombination for rice breeding. Rice 2015, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.F.; Ladejobi, O.; Amer, S.; Bentley, A.R.; Biernaskie, J. Multi-parent populations in crops: A toolbox integrating genomics and genetic mapping with breeding. Heredity 2020. [Google Scholar] [CrossRef]
- Descalsota, G.I.L.; Swamy, M.; Zaw, H.; Asilo, M.A.I.; Amparado, A. Genome-wide association mapping in a rice magic plus population detects QTLS and genes useful for biofortification. Front. Plant Sci. 2018, 9, 1347. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Liu, G.Q.; Yan, H.H.; Fu, Q.; Qian, Q.; Zhang, Z.T.; Zhai, W.X. Mapping of a new gene for brown planthopper resistance in cultivated rice introgressed from Oryza eichingeri. Chin. Sci. Bull. 2001, 46, 738–742. [Google Scholar]
- Sun, L.; Su, C.; Wang, C.; Zai, H.; Wan, J. Mapping of a major resistance gene to brown plant hopper in the rice cultivar RathuHeenati. Breed. Sci. 2005, 55, 391–396. [Google Scholar]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. Development and characterization of japonica rice lines carrying the brown plant hopper-resistance genes BPH12 and BPH6. Theor Appl. Genet. 2012, 124, 485–494. [Google Scholar] [CrossRef]
- Khush, G.S.; Virk, P.S. IR Varieties and Their Impact; International Rice Research Institute: Manila, Philipines, 2005. [Google Scholar]
- Myint, K.K.M.; Fujita, D.; Matsumura, M.; Sonoda, T.; Yoshimura, A.; Yasui, H. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stål]) in the rice cultivar ADR52. Theor. Appl. Genet. 2012, 124, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Gao, F.; Wu, X.; Lu, X.; Zeng, L.; Lv, J. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Huang, D.R.; Chen, J.; Lai, F.X.; Liu, G.J.; Zhuang, J.Y. Analysis of quantitative trait loci for resistance to brown planthopper in Dongxiang wild rice (Oryza rufipogon Griff.). Acta Agron. Sin. 2012, 38, 210–214. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, J.; Wang, Z.Z.; Jing, S.L.; Wang, Y.; Ouyang, Y.D. Allelic diversity in an NLR gene BPH9 enables rice to combat plant hopper variation. Proc. Natl. Acad. Sci. USA 2016, 113, 12850–12855. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.; Kim, S.R.; Kim, Y.H.; Suh, J.P.; Park, H.M.; Sreenivasulu, N. Map-based cloning and characterization of the BPH18 gene from wild rice conferring resistance to brown planthopper (BPH) insect pest. Sci. Rep. 2016, 6, 34376. [Google Scholar] [CrossRef]
- Tamura, Y.; Hattori, M.; Yoshioka, H.; Yoshioka, M.; Takahashi, A.; Wu, J.Z. Map-based cloning and characterization of a brown plant hopper resistance gene BPH 26 from Oryza sativa L. ssp. indica cultivar ADR52. Sci. Rep. 2014, 4, 5872. [Google Scholar] [CrossRef] [Green Version]
- Khong, G.N.; Richaud, F.; Coudert, Y.; Pati, P.K.; Santi, C.; Périn, C. Modulating rice stress tolerance by transcription factors. Biotech. Gen. Eng. Rev. 2008, 25, 381–404. [Google Scholar] [CrossRef] [Green Version]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Qiu, D.; Xiao, J.; Ding, X.; Xiong, M.; Cai, M.; Cao, Y. OsWRKY13 mediates rice disease resistance by regulating defense-related genes in salicylate- and jasmonate-dependent signaling. Mol. Plant. Microbe Interact. 2007, 20, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wu, Y.; Wu, D.; Rao, W.; Guo, J.; Ma, Y. The coiled-coil and nucleotide binding domains of brown planthopper resistance 14 function in signaling and resistance against planthopper in rice. Plant Cell 2017, 29, 3157–3185. [Google Scholar] [CrossRef] [Green Version]
- Fromont-Racine, M.; Senger, B.; Saveanu, C.; Fasiolo, F. Ribosome assembly in eukaryotes. Gene 2003, 313, 17–42. [Google Scholar] [CrossRef]
- Ferreira-Cerca, S.; Hurt, E. Cell biology: Arrest by ribosome. Nature 2009, 459, 46–47. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
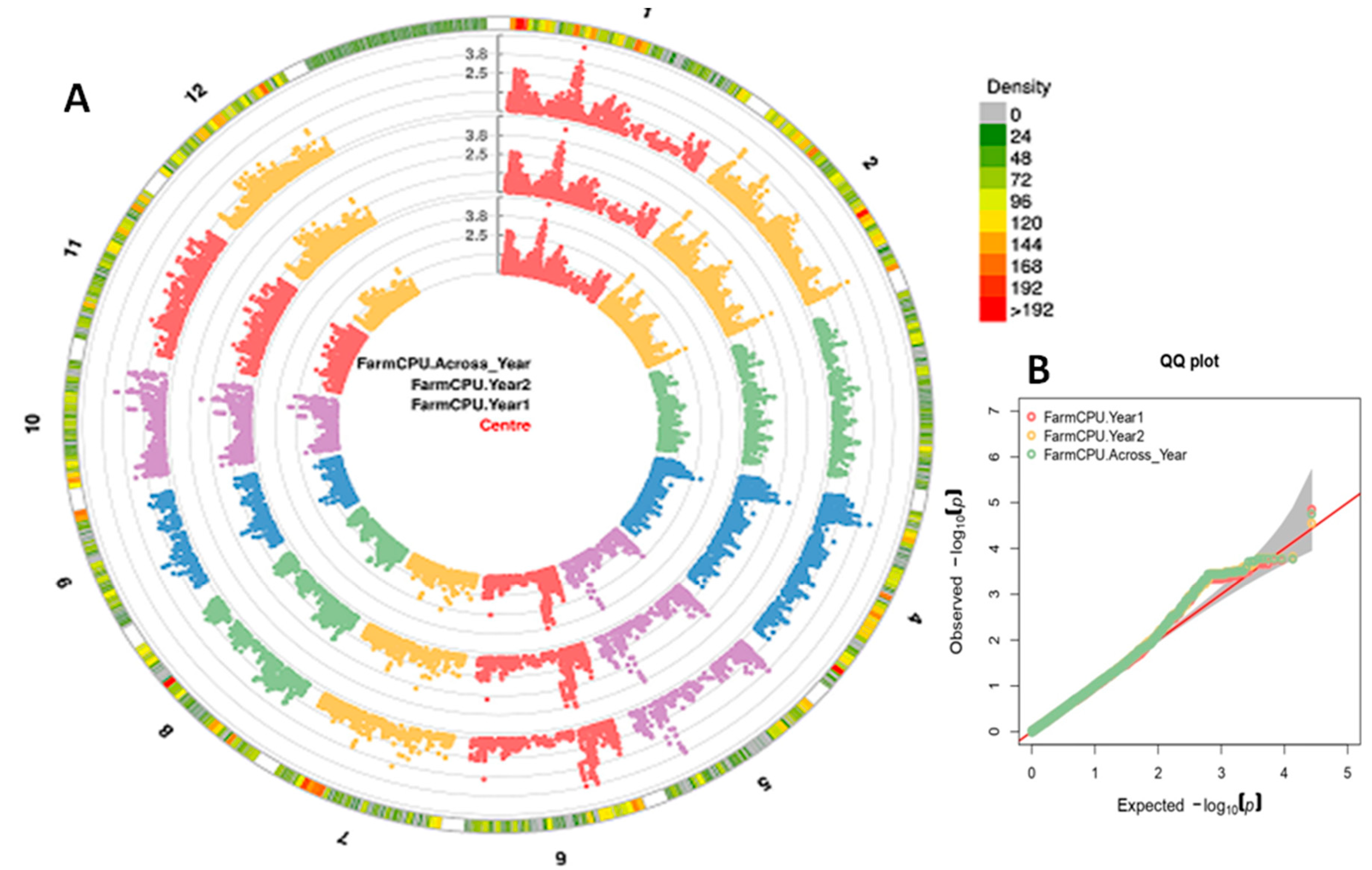{kind=link}
| Source | df | 2016 | 2017 | Mean Over Years |
|---|---|---|---|---|
| Treatment (ignoring blocks) | 393 | 4.07 ** | 4.47 ** | 4.30 ** |
| Treatment: Check | 2 | 172.43 ** | 182.1 ** | 178.53 ** |
| Treatment: Test vs. Check | 1 | 313.6 0 ** | 318.46 ** | 318.45 ** |
| Treatment: Test | 390 | 2.42 ** | 2.76 ** | 2.60 ** |
| Block (eliminating treatments) | 9 | 0.08 ns | 0.08 ns | 0.09 ns |
| Residuals | 18 | 0.21 | 0.10 | 0.09 |
| Chromosome | GLM | MLM | CMLM | SUPER | Farm CPU | MLMM | Common SNPs |
|---|---|---|---|---|---|---|---|
| 1 | 28 | 12 | 8 | 30 | 20 | 26 | S1_12742211, S1_13273091, S1_13274423, S1_13365703, S1_13374131, S1_13374151, S1_13424589 |
| 2 | 21 | 1 | 5 | 13 | 20 | 13 | S2_34796411 |
| 4 | 13 | 2 | 1 | 11 | 11 | 4 | S4_4571354 |
| 5 | 40 | 14 | 14 | 39 | 39 | 40 | S5_23256042, S5_23257107, S5_23257108, S5_23279024, S5_23279042, S5_23287785, S5_23303726, S5_2330752, S5_23310970, S5_23312204, S5_23314218 |
| 6 | 90 | 49 | 40 | 60 | 88 | 61 | S6_6769492, S6_6769526, S6_6769533, S6_7646336, S6_8850552, S6_8851568, S6_8870236, S6_8876540, S6_8876586, S6_8876606, S6_8876610, S6_8886408, S6_8886434, S6_8914643, S6_8914650, S6_8914651, S6_8914671, S6_8914769, S6_8917185, S6_8917933, S6_8917936, S6_8917962, S6_8921200, S6_8921201, S6_8932488, S6_8956202, S6_8962742, S6_8962796, S6_8975500, S6_8977107, S6_8977116, S6_8977156, S6_8977190, S6_9003844, S6_9003866, S6_9004225 |
| 7 | 6 | - | - | 1 | 5 | - | - |
| 10 | 8 | 9 | 8 | 2 | 7 | 19 | S10_18127881, S10_18145180 |
| 11 | - | - | - | 2 | - | - | - |
| 12 | 7 | 4 | 4 | 4 | - | 4 | - |
| Total | 213 | 91 | 80 | 162 | 190 | 167 |
| S. No. | Gene locus ID | Chromosome | QTNs | Putative Function |
|---|---|---|---|---|
| 1 | LOC_Os01g24690 | 1 | S1_13898444 | 60S ribosomal protein L23A, putative, expressed, response to abiotic stimulus, response to stress |
| 2 | LOC_Os05g39590 | 5 | S5_23249078, S5_23249119, S5_23249125, S5_23249237, S5_23249605 | AP2 domain containing protein, expressed |
| 3 | LOC_Os05g39720 | 5 | S5_23310970, S5_23312204, S5_23314218 | WRKY70, expressed |
| 4 | LOC_Os06g11010 | 6 | S6_5759360 | Eukaryotic aspartyl protease domain containing protein, expressed |
| 5 | LOC_Os06g12160.1 | 6 | S6_6513819, S6_6514012 | AAA-type ATPase family protein, putative, expressed |
| 6 | LOC_Os06g12360.1 | 6 | S6_6697070 | Pentatricopeptide, putative, expressed |
| 7 | LOC_Os06g12610.1 | 6 | S6_6869009 | Auxin efflux carrier component, putative, expressed |
| 8 | LOC_Os06g12870.1 | 6 | S6_7045328, S6_7045355 | Leaf senescence related protein, putative, expressed |
| 9 | LOC_Os06g13600.1 | 6 | S6_7531094, S6_7531433, S6_7531437, S6_7538495 | HEAT repeat family protein, putative, expressed |
| 10 | LOC_Os06g14510.3 | 6 | S6_8153078 | Glucose-6-phosphate isomerase, putative, expressed |
| 11 | LOC_Os06g15750 | 6 | S6_8932488 | NB-ARC domain containing protein, expressed |
| 12 | LOC_Os06g15820.1 | 6 | S6_8977107, S6_8977116, S6_8977156, S6_8977190, S6_8977712, S6_8977949, S6_8977972, S6_8982135 | NHL repeat-containing protein, putative, expressed |
| 13 | LOC_Os07g18600 | 7 | S7_10995384 | OsFBL37-F-box domain and LRR containing protein, expressed |
| S. No | Chromosome | SNP Position | Allelic Status | Putative Function |
|---|---|---|---|---|
| 1 | 5 | 23249125 | C | AP2 domain containing protein, expressed |
| 2 | 6 | 5759360 | T | Eukaryotic aspartyl protease domain containing protein, expressed |
| 3 | 6 | 6513819 | T | AAA-type ATPase family protein, putative, expressed |
| 4 | 6 | 6514012 | G | |
| 5 | 6 | 6697070 | T | Pentatricopeptide, putative, expressed |
| 6 | 6 | 6869009 | G | auxin efflux carrier component, putative, expressed |
| 7 | 6 | 7045328 | C | Leaf senescence related protein, putative, expressed |
| 8 | 6 | 7045355 | G | |
| 9 | 6 | 7531094 | C | HEAT repeat family protein, putative, expressed |
| 10 | 6 | 7538495 | G | |
| 11 | 6 | 8153078 | G | Glucose-6-phosphate isomerase, putative, expressed |
| 12 | 7 | 10995384 | C | OsFBL37 - F-box domain and LRR containing protein, expressed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satturu, V.; Vattikuti, J.L.; J, D.S.; Kumar, A.; Singh, R.K.; M, S.P.; Zaw, H.; Jubay, M.L.; Satish, L.; Rathore, A.; et al. Multiple Genome Wide Association Mapping Models Identify Quantitative Trait Nucleotides for Brown Planthopper (Nilaparvata lugens) Resistance in MAGIC Indica Population of Rice. Vaccines 2020, 8, 608. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8040608
Satturu V, Vattikuti JL, J DS, Kumar A, Singh RK, M SP, Zaw H, Jubay ML, Satish L, Rathore A, et al. Multiple Genome Wide Association Mapping Models Identify Quantitative Trait Nucleotides for Brown Planthopper (Nilaparvata lugens) Resistance in MAGIC Indica Population of Rice. Vaccines. 2020; 8(4):608. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8040608
Chicago/Turabian StyleSatturu, Vanisri, Jhansi Lakshmi Vattikuti, Durga Sai J, Arvind Kumar, Rakesh Kumar Singh, Srinivas Prasad M, Hein Zaw, Mona Liza Jubay, Lakkakula Satish, Abhishek Rathore, and et al. 2020. "Multiple Genome Wide Association Mapping Models Identify Quantitative Trait Nucleotides for Brown Planthopper (Nilaparvata lugens) Resistance in MAGIC Indica Population of Rice" Vaccines 8, no. 4: 608. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8040608