Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities



1
Clinical Division of Fish Medicine, University of Veterinary Medicine Vienna, 1210 Vienna, Austria
2
Central Marine Fisheries Research Institute, Karwar 581301, India
*
Author to whom correspondence should be addressed.
Vaccines 2021, 9(2), 179; https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020179
Submission received: 27 January 2021
/
Revised: 16 February 2021
/
Accepted: 17 February 2021
/
Published: 20 February 2021
(This article belongs to the Section Veterinary Vaccines)
Abstract
:Globally, parasites are increasingly being recognized as catastrophic agents in both aquaculture sector and in the wild aquatic habitats leading to an estimated annual loss between 1.05 billion and 9.58 billion USD. The currently available therapeutic and control measures are accompanied by many limitations. Hence, vaccines are recommended as the “only green and effective solution” to address these concerns and protect fish from pathogens. However, vaccine development warrants a better understanding of host–parasite interaction and parasite biology. Currently, only one commercial parasite vaccine is available against the ectoparasite sea lice. Additionally, only a few trials have reported potential vaccine candidates against endoparasites. Transcriptome, genome, and proteomic data at present are available only for a limited number of aquatic parasites. Omics-based interventions can be significant in the identification of suitable vaccine candidates, finally leading to the development of multivalent vaccines for significant protection against parasitic infections in fish. The present review highlights the progress in the immunobiology of pathogenic parasites and the prospects of vaccine development. Finally, an approach for developing a multivalent vaccine for parasitic diseases is presented. Data sources to prepare this review included Pubmed, google scholar, official reports, and websites.
1. Introduction
Aquaculture continues to be one of the rapid food-producing sectors worldwide. According to an estimate, the latest global aquaculture production was 82 million tons and valued at 250 billion USD in 2018 [1]. However, the sector is frequently hit by several viral, bacterial, and parasitic diseases with devastating consequences [2]. Although viruses and bacteria have been recognized as the leading cause of huge economic losses to the sector, the role of parasites has been realized recently. Growing literature suggests a considerable increase in parasitic epidemics both in farmed and wild fish populations. Parasites belonging to different groups such as myxozoa, protozoa, crustaceans, monogeneans, and helminths result in heavy losses in aquaculture and consequently to the allied industries. As per a report, the annual global loss of juvenile fish on account of parasitic infections was estimated to vary from 107.31 to 134.14 million USD and loss of marketable size fish from 945.00 million to 9.45 billion USD, the total estimate being 1.05 billion to 9.58 billion USD [3].
The management of parasitic infections in culture facilities involves different strategies such as quarantine, disease-free sites, disinfection of water using UV radiation and chemicals, fallowing, and drying of pond bottoms [4]. Instead of ponds, concrete tanks or raceways have been used for fish culture to prevent infection by myxozoan parasites in order to restrict the oligochaetes, which serve as the alternate hosts of these parasites [5]. Nevertheless, early efforts of controlling a parasitic infection in fish relied heavily on the use of chemotherapeutics. Consequently, their relentless use is leading to the emergence of drug resistance [6,7] and deleterious environmental effects [8,9]. Although phytotherapy-based treatment options are favorable, they suffer from various disadvantages [10,11,12,13]. Lately, the use of attractants and traps has been suggested as a promising strategy for certain parasites such as sea lice, by exploiting their chemotactic and phototactic responses [14]. Furthermore, a study highlighted the use of urea and light-based traps for controlling the infection by Cryptocaryon irritans and Neobenedenia girellae in aquaculture [15]. However, at present, application of this approach in commercial aquaculture is limited due to the unavailability of efficient traps. Vaccination is considered the best method for safeguarding and promoting fish health and welfare against any parasite. Although several commercial vaccines are available for bacterial and viral diseases globally [16], only Chile has a commercial parasite vaccine against sea lice [17]. The development of parasite vaccines is limited by several inherent issues. One of the most important factors is the biological complexity of the parasite. The parasite life advances through different developmental stages, which may have a specific antigen profile. Moreover, the life cycle stages alternate between different host species in various parasites [18], thus, interfering with the culturing and maintenance of parasites under laboratory conditions owing to requirements for optimization of several parameters such as temperature and nutritional elements for the alternate host, giving rise to more labor requirement and increased economic costs.
Omics studies are powerful methods for developing vaccines by providing potential vaccine candidates. The suffix “omics” refers to the high-throughput analysis of cellular macromolecules. The most popular omics disciplines include genomics, transcriptomics, and proteomics. The omics era started with genomics, aiming to study the entire gene content (genome) of an organism [19]. The genomic analysis provides abundant information on individual genes, chromosomes, their organization, genetic variants of diseases as well as evolutionary relationships with other phyla and parasites. However, genomics does not provide information on aspects such as gene expression, function and regulation, and structure and characteristics of encoded proteins [20]. These limitations have resulted in the advent of the post-genomic era primarily dominated by transcriptomics and proteomics. A transcriptome comprises all RNA transcripts produced by the genome under a given environmental condition [21]. Transcriptomic profiling provides information on different categories of transcribed RNAs, the transcriptional structure of genes, and the expression of genes [22]. Nonetheless, transcriptomics does not reflect the actual protein complement due to the many events in translation of mRNA transcripts, e.g., post-transcriptional modifications and alternate splicing [23]. Proteomics followed transcriptomics, which is defined as the study of proteomes (the entire set of proteins, that are the key players in biological processes) [21].
Genomic and transcriptomic analyses of certain fish parasites have been performed. In addition, transcriptomics-based studies have been conducted to investigate the host–parasite interaction in some instances [24]. Similar to human and veterinary parasitology, omics data could provide potential therapeutic targets against aquatic parasites.
In this review, we highlight the need and the progress made in developing parasitic vaccines, with particular focus on the immunobiology of fish parasites. Furthermore, we discuss the present status, the prospects of developing successful parasite vaccines in the present era of omics, and an approach for developing multivalent vaccines.
2. Data Sources, Searches and Study Selection
Searches were performed on pubmed, google scholar and google with the keywords either alone or in combination “parasite vaccines”, “fish parasites”, “transcriptomics”, “genomics”, and “proteomics”. According to the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines, peer-reviewed articles were initially selected. The articles were then screened based on title and the abstract. Only full text articles were included in the study. Official reports of FAO (Food and Agriculture Organization of the United Nations), NCBI (National Center for Biotechnology Information) and OIE (Office International des Epizooties) websites were also referred to. Flow diagram is provided as Supplementary Data (Figure S1).
3. Economically Important Fish Parasites
The majority of the important fish endoparasites belong to the phylum Cnidaria, whereas the ectoparasites belong to phyla Ciliophora and Arthropoda (Table 1). The phylum Platyhelminthes encompasses both endoparasites and ectoparasites. Parasitic diseases in aquaculture cause both direct and indirect losses. Direct losses result from the mortality in farmed fish because of parasite outbreaks. Indirect losses are attributed to the investment made for treating infections, adopting management strategies, reduced growth resulting from infection, and the cost involved from carcass spoilage at harvest [3,18,25].
Some notable examples of commercially important myxozoan endoparasites include Tetracapsuloides bryosalmonae, Myxobolus cerebralis, Sphaerospora molnari, Ceratomyxa shasta, Kudoa thyrsites, and Enteromyxum sp. Proliferative kidney disease (PKD), caused by T. bryosalmonae, could result in up to 95% mortality in farmed salmonids [26]. C. shasta causes ceratomyxosis in chinook and coho salmon [38]. In addition, several other myxozoan parasites incur huge direct and indirect economic losses, e.g., Myxobolus cerebralis [5] and Enteromyxum sp. [39]. Some ectoparasites responsible for significant mortality and thus aquaculture losses include Ichthyophthirius multifiliis, C. irritans, Argulus, Lepeophtheirus salmonis, and Caligus rogercresseyi. Protozoan parasites I. multifiliis and C. irritans cause white spot disease in freshwater and marine fishes, respectively [18]. The crustacean parasites L. salmonis and C. rogercresseyi, commonly referred to as sea lice, cause severe infection in salmonids [40]. Argulus is the freshwater counterpart of sea lice reported from several fish hosts [41]. Furthermore, several parasites have been implicated in the decline of wild fish populations, e.g., M. cerebralis [42], T. bryosalmonae [43], C. shasta [44], I. multifiliis [45], L. salmonis and C. rogercresseyi [40].
Fish species of aquaculture importance differ with countries. Moreover, parasites causing mortality and incurring economic losses may differ. However, certain parasites are of global concern, e.g., M. cerebralis, I. multifiliis, and Argulus [18]. Despite reports of huge economic losses and fish mortality due to parasitic infections [18], their estimations are either lacking or are specific to locations. For instance, losses in salmonid farming accounted for 6 million Canadian dollars (CAD) in 2015 due to K. thyrsites infection in British Columbia, Canada [33]. Argulus is reported to cause losses amounting to 5.41 million United States dollars (USD) annually in carp aquaculture in India [31]. Globally, annual indirect and direct losses in salmonid aquaculture due to infestations with the sea louse L. salmonis have been estimated to be 500 million to 1 billion USD [46].
4. Progress in Understanding of Host-Parasite Interactions
Teleosts are the lowest vertebrates that elicit both innate and adaptive immune responses against pathogen invasion [47]. However, compared to higher vertebrates, innate immunity plays a major role than the adaptive immune response in aquatic organisms, irrespective of the nature of the pathogen [48]. Remarkably, innate immune reactions are not only the first line of defense but also set the pace for ensuing adaptive immune responses. Thus, the development of an effective vaccine requires a good understanding of immune response during host–parasite interaction and pathogenesis mechanisms. Some progress has been made in understanding the immune response in fish against parasitic diseases. Studies using immunoassays, gene expression, Western blotting, and other techniques have elucidated the expression of different molecules in innate and adaptive immune responses during several parasitic infections. The generation of an elevated immune response upon re-infection in fish, which could survive after previous natural parasite infection, has also been reported. Few examples include endoparasites, e.g., T. bryosalmonae, C. shasta, E. scophthalmi, E. leei [49] and ectoparasites, e.g., I. multifiliis and C. irritans [45,50]. Similar to other vertebrates, the development of immune memory in fish infected with parasites forms the fundamental basis of vaccine development.
4.1. Innate Immune Response during Selected Parasitic Infections
Studies on the immune response generated by parasites during infection and post-immunization, report the involvement of toll-like receptors (TLRs), phagocytes, complement proteins, melanomacrophage centers, proteases, and cytokines. TLRs are type-I transmembrane receptors that recognize pathogen-associated molecular patterns to initiate an immune response [51]. Several TLRs play important roles in the defense of different fish hosts, against various parasites e.g., T. bryosalmonae [52], I. multifiliis [53], C. irritans [54], and C. rogercresseyi [55]. TLR1, TLR2, TLR9, TLR19, TLR21, and TLR25 reportedly play an important role in immunity in fish against I. multifiliis [53]. However, the concerned parasite epitopes that they recognize are unexplored currently for the majority of TLRs. For example, TLR1, TLR13, and TLR19 are significantly upregulated in the kidney of brown trout during PKD [52], but the moiety of T. bryosalmonae, which might be responsible for their activation is unknown. TLR2 on human NK cells are activated by lipophosphoglycan component in the hemoflagellate parasite Leishmania major, a phosphoglycan belonging to a family of unique Leishmania glycoconjugates [56]. Phagocytic cells such as neutrophils, acidophilic granulocytes, and monocyte-macrophages, are involved in pathogen clearance [57] by producing reactive oxygen intermediates and nitric oxide, resulting in a respiratory burst. Increased phagocytic oxidative burst due to the enhanced capability of phagocytes to produce a higher amount of reactive oxygen species was observed in natural T. bryosalmonae infection in rainbow trout [58]. A higher number of NBT-positive cells indicated similar results in fish immunized with either Sphaerospora dicentrarchi spores alone or in combination with an adjuvant on the seventh day post-injection [59]. Respiratory burst and abundant granulocytes were found in vaccinated fish with live Cryptobia salmositica [60]. Furthermore, the increased number of thrombocytes and monocytes indicated clearance of S. molnari by phagocytosis. Increased thrombocytes acted against inflammation and contributed to wound healing [61]. Moreover, these authors associated the immediately increased expression of cytokines such as interleukin-1β upon peritoneal injection of the parasite with parasite recognition by host TLRs.
Nonspecific cytotoxic cells (NCC) in fish comprise heterogenous leucocyte populations. These are considered as mammalian equivalents of natural killer cells (NK) sharing many characters in common including antiparasitic activity [62]. NCC occur in teleost thymus, kidney, spleen and blood. They are particularly considered important in young fish when specific responses are weak [63]. The antiparasitic activity of NCC is reported against I. multifiliis [62] and Tetrahymena pyriformis [64]. However, more information on the role of these cells during parasitic infections is required.
In vitro studies with inactivated serum have reported the function of the classical complement pathway in killing Philasterides dicentrarchi [65]. Melanomacrophage centers (MMCs) are aggregation of phagocytic cells in lymphoid organs and involved in pathogen sequestration and antigen presentation [66]. MMCs have been found to contain different myxozoan parasites such as Enterozoon scophthalami [67] and Myxobolus cyprini [68]. The non-specific cellular innate immune response plays a greater role in the recovery from infection with parasites such as T. bryosalmonae [58]. The antiparasitic activity of different fish antiproteases such as α-2 macroglobulin during E. scophthalmi infection [69], total serum antiprotease in E. leei [67], and serine protein inhibitors [70] have been reported.
Only a few studies have been conducted to understand the susceptibility and resistance to parasites by host fish. For example, the upregulation of IFN-γ was reported both in the resistant Hofer strain and the susceptible US strain of rainbow trout against M. cerebralis infection. However, the upregulation of this gene was found to be considerably higher in susceptible trout strains than in the resistant strains after 24 h of pathogen exposure. The higher expression of the gene in susceptible rainbow trout was suggested to be deleterious for the host [71]. Similar observations in susceptible and resistant strains of Chinook salmon to C. shashta infection have been reported [72]. IFN-γ has been identified as a key immune effector cytokine with multiple protective roles via enhancing antigen presentation by APCs and initiation of pro-inflammatory responses in coordination with other pathways. Its activity is highly regulated, and its overexpression can damage the host tissue [48]. Among the different genes analyzed, STAT-3 (signal transducer and activator of transcription 3) was the most differentially expressed gene during M. cerebralis infection in susceptible and resistant rainbow trouts. In the Hofer strain rainbow trout, the higher expression of STAT-3 gene likely confers resistance against whirling disease. The expression remains unchanged in susceptible rainbow trout during infection [71]. SOCS (suppressor of cytokine signaling) protein family negatively regulates cytokine and growth factor signaling [73]. T. bryosalmonae and M. cerebralis infection results in the overexpression of SOCS-1 and SOCS-3 genes in rainbow trout during parasite development [74]. The innate and adaptive immune systems are interconnected. Therefore, the knowledge of innate immune effectors in protection, susceptibility, and resistance against parasites in fish is paramount in designing successful vaccines.
Some parasites reportedly evade or suppress the innate immune responses for their continued survival in the fish host. For instance, the ectoparasite I. multifiliis invokes a host evasion mechanism by ingestion of neutrophils, thus suppressing further signaling pathways in the immune reactions [75]. L. salmonis induces limited innate immune response in one of its most susceptible salmonid host Atlantic salmon which might be an immune suppression approach. Mustafa et al. [76] reported depression in oxidative and phagocytic abilities of host macrophage following L. salmonis infection which could be a parasite strategy. Another possible mechanism might be the increased production of proteases by the parasite. L. salmonis reportedly produces trypsin-like proteases which help them to feed and evade host immune responses. Dalvin et al. [77] reported the suppression of complement and nitric oxide in rainbow trout infected with sea lice. Nitric oxide is a signaling molecule leading to an anti-inflammatory response. It modulates the release of different inflammatory mediators from the cells participating in inflammatory responses, e.g., leukocytes, macrophages, mast cells, endothelial cells, and platelets [78]. Thus, suppression of nitric oxide could be a possible parasite strategy to avoid excessive immune response and increase survival of fish host. Additionally, T. bryosalmonae causes a severe inflammatory response in the kidney of brown trout which gradually regresses over a period of time and the recovered fish continue to excrete parasite spores [43].
4.2. Adaptive Immune Response during Selected Parasite Infection
Three classes of immunoglobulins have been described in fish, namely IgM, IgD, and IgT [79]. Parasite-specific antibodies have been detected in several parasitic infections, e.g., T. bryosalmonae [80,81], M. cerebralis [82], S. dicentriarchi [83], C. shasta [84], E. scophthalmi [85], M. honghuensis [86], I. multifiliis [75] and L. salmonis [77]. IgT might be playing important roles against fish parasites. In fish surviving C. shasta infection, a higher number of IgT+ B cells were found than in healthy fish [87]. The authors confirmed the IgT protein abundance and the upregulation of IgT gene. IgT is reportedly the dominant immunoglobulin present in skin and gills of I. multifiliis-infected rainbow trout. Abundant IgT+ B cells were found to occur in the skin epidermis of infected fish. Additionally, IgT was found to cover the parasite and was present in the mucus at high quantities [88]. All three immunoglobulins are formed in response to PKD; however, these might act as an escape mechanism following infection with T. bryosalmonae. This study also suggested an important role of IgD in the humoral response to the parasite, based on the appearance of IgD+ IgM− B cells, somatic hypermutation, and clonal expansion of some IgD-expressing B cell subsets [89]. During I. multifiliis infection, antibodies cross-bind to Iag allowing the parasite to either escape from the host or result in destruction of the parasite [45]. During certain parasitic infections, antibodies were not detected. This response was observed in C. elongatus-infected Atlantic salmon because this parasite causes little damage to fish skin leading to decreased contact between louse antigen and the host immune system [90].
T cells are involved in adaptive immunity in all vertebrates. These cells are characterized by the presence of T cell receptors [91]. At present, studies on T cell-mediated immune response in fish parasites are mostly limited to the associated surface markers. Experimental infection by anal intubation of E. leei-infected intestinal scraping in gilthead seabream provided evidence for specific T cell response in the head kidney and anterior intestine [35]. In this study, real-time PCR revealed the downregulation of T cell markers such as zap70, cd3, cd4–1, cd4–2, cd8β, and CTL receptors in the head kidney and their upregulation in the anterior intestine. Moreover, FACS analysis provided evidence on the involvement of CD8− T cells in resistance against M. cerebralis infection in the German Hofer strain of rainbow trout [92]. During infection with the intracellular parasite K. thyrsites, a Th 1 type response was reported as indicated by the upregulation of il-12 gene in the infected muscles of Atlantic salmon [33]. Currently, T cell responses are measured by ELISPOT assay, which is considered the most sensitive determination method of T cell cytokine production. Additionally, other assays are also used like intracellular staining for cytokines and others markers of T cell activation and function, in vivo CTL assay for measuring the lytic capacity of cytolytic T cells, which provide high sensitivity detection of specific pathogen peptides. These assays are useful in studying the specificity and potency of T cell responses of host and high throughput antigen screening [93]. Adaptive immune response mediated by high antibody titers with decreasing parasitemia was noticed following live vaccination against C. salmositica in Atlantic salmon [60]. Both the components of the adaptive immune system, i.e., the humoral response mediated by antibodies and the cellular response mediated by T cells are demonstrated in fish against parasitic infections. The humoral response is important against extracellular parasites, whereas T cell-mediated immune response is induced against intracellular parasites. Thus, an effective vaccination strategy would stimulate both these arms of the adaptive immune system [94].
5. Parasite Vaccines: Status and Prospects
Currently, there exist 24 commercially available vaccines for viral and bacterial diseases of aquaculture importance [16]. However, only one commercial parasite vaccine, Providean Aquatec Sea Lice produced by Tecnovax S.A. Argentina, is available. As discussed earlier, developing vaccines against aquatic parasites is an extremely challenging task.
Progress towards the development of parasite vaccines for aquaculture has been comparatively slower than those for humans and animals. In addition to the reasons mentioned, several other possibilities exist for this dismal scenario. A possible reason in several countries could be the smaller scale of aquaculture operations resulting in economic impracticality of vaccine production, the recent realization of their importance in causing huge disease losses, and fewer research groups working on this aspect. The available literature suggests that vaccines for parasites infecting fish are confined to a few trials against a limited number of parasites.
5.1. Ectoparasite Vaccine Trials
Vaccines have been attempted for parasites with significant detrimental effects on aquaculture. These include vaccination trials against parasites such as I. multifiliis, C. irritans, and L. salmonis (Table 2). Different vaccine trials comprising live, killed, parasite homogenate, subunit, and DNA have been tested for protection against I. multifiliis infection in various fish hosts. The earliest, as well as the most effective method of achieving protection to date remains immunization by live theronts or trophonts [30]. Immunization trials against this parasite with ciliary and whole cell preparation of I. multifiliis and Tetrahymena pyriformis were used to vaccinate channel catfish. The ciliary antigen of the latter was found to confer effective protection [95]. The lower protection rate obtained using I. multifiliis cilia could be attributed to less homogenous antigen preparation, resulting from the lower number of tomites used. The same fine structure of all protozoan cilia [96] and common antigenic determinant [97] provided cross protection. Burkart et al. [98] studied different antigen preparations and vaccination routes for immunizing channel catfish against I. multifiliis. The study concluded that intraperitoneal administration of live tomites effectively protected against infection as compared with a surface infection with the parasite. Fish vaccinated with formalin-killed trophonts resulted in a 51% mortality rate. Sonicated and formalin-killed trophont formulations protected rainbow trout ten weeks post-hatch fry. Moreover, this study reported a higher susceptibility to reinfection of bath-treated fish in comparison to IP-injected fish. However, the underlying reasons could not be ascertained [98]. Proteomic screening and in silico analysis were used to test three recombinant proteins (#5, #10, and #11), which were combined in a subunit vaccine and administered intraperitoneally in rainbow trout and resulted in partial protection. Parasite burden was found to be lower with a mean intensity of 1.3 parasites per g/cm fish in the vaccinated group as compared with the uninfected control group (2.8 parasites per g/cm fish).
In addition, specific antibody production was found to be significantly higher than in control fish. The highest antibody response was generated in fish against protein #10 both at four weeks and 13 weeks post-infection, suggesting it as a potential vaccine candidate along with cell and ciliary surface antigens commonly known as immobilization antigen or i-antigen [101]. For the DNA vaccine, the intramuscular administration of i-antigen alone or in combination with cysteine protease of I. multifiliis has been used in channel catfish and rainbow trout as a vaccine candidate [99,102]. However, the protection of DNA vaccine was comparatively lower than that of a live vaccine. Both live and killed C. irritans theronts conferred protective immunity in grouper, as evident from the high antibody titer in the mucus of immunized fish and better survival and reduced number of tomonts and trophonts [106].
5.2. Endoparasite Vaccine Trials
Currently, the literature on vaccination trials for endoparasites is available only for T. bryosalmonae, Myxobolus koi, and Uronema marinum (Table 3). DNA vaccine targeting a novel micro-exon genes (Tb MEG1) has been reported to elicit Tb-MEG1-specific immune response in rainbow trout [108]. Moreover, this study demonstrated the expression of proteins in and on the surface of parasites using anti-Tb-MEG1 monoclonal antibodies. A study using crude spore proteins reported better survival in M. koi-infected gold fish [109]. Intraperitoneal administration of poly D, L-lactic-co-glycolic acid (PLGA)-encapsulated vaccine against U. marinum infection significantly reduced the mortality in kelp grouper, Epinephelus bruneus [110]. This study reported early enhancement (1 to two weeks post-vaccination) and longer duration (4 weeks post-vaccination) of respiratory burst, complement activity, α2-macroglobulin activity, serum lysozyme, antiprotease activity, and antibody response. Studies on vaccination trials for ecto- and endoparasites report the use of different criteria for determining the efficacy of the tried vaccine, such as percentage mortality, percentage survival, parasite burden, and antibody titer in vaccinated fish as compared with those in the control fish.
6. Perspectives in Fish Parasite Vaccine Development
6.1. Vaccination Strategy
Live, killed, DNA and protein subunit vaccines are being applied in aquaculture to control various pathogenic organisms. The majority of the vaccine trials in fish against parasites have used the live or killed strategy that has proved effective. However, it has certain limitations. Live vaccination is similar to the natural process of infection. This approach employs a controlled infection with a virulent or less virulent or attenuated strain of the parasite. In the case of fish, the attenuated strains of few parasites has been used for immunizing fish. There is an increased risk of mortality with vaccination with live virulent parasites. Another important consideration for both live and killed vaccines is the requirement of numerous parasites of good quality, which currently is difficult to culture in vitro. In addition, live vaccines are not considered safe for use in aquaculture [16]. Although DNA and subunit vaccines are the most promising alternative approaches, both the approaches have conferred partial to moderate protection. This could be attributed to the fact that the parasites present several challenges such as antigenic variability, immune evasion, immunomodulation of effector molecules, and poor immunogenicity of individual antigens. To overcome these issues, the vaccine must target multiple antigens (multi-epitope) simultaneously to be effective. Although a successful example of a multivalent vaccine approach for parasite is not available from fish-based trials, a multivalent DNA vaccine encoding three antigens provided long-lasting protection after mice were challenged with Leishmania [112]. Certain factors must be considered while attempting a subunit vaccine, including the technical viability of antigen production, its formulation in suitable adjuvants, and the ease and frequency of delivery [113]. Similarly, the selection of expression vector must ensure the required post-translational processing to retain the immunogenicity of the desired protein.
6.2. Vaccination Routes
Broadly, vaccines can be administered in aquaculture via oral, immersion, and injection routes. In the oral process, antigens encapsulated in the feed are administered to fish. It is the most effective delivery method for aquaculture due to minimal stress, simple administration, and applicability to both large and small fish. However, difficulty in determining the precise dosage received by fish and lack of efficacy limit its application. In addition, antigen degradation is possible during its passage to the stomach before it reaches the hindgut where the antigen is adsorbed [114]. In the immersion process, fish are immersed in water with antigens for a specified period. Immersion can be performed in the form of dip and bath. In the dip method, the fish are kept in water containing antigens at a high dose for some minutes, whereas the bath method involves keeping the fish in water with a low antigen dose for a longer time [115]. The injection route is used to commonly deliver the antigen either intraperitoneally or intramuscularly. The advantages of injection vaccination include precise dosage delivery and longer protection [116]. In most studies, the intraperitoneal route of vaccine administration against parasites was used for live and killed antigen preparations, whereas the intramuscular route has been used for DNA antigens (Table 2 and Table 3). Several studies report significant systemic and mucosal antibody production against parasites following the intraperitoneal injection of antigens [98,101]. Certain disadvantages associated with the injection process include stress during handling and anesthetizing. In addition, the method is labor intensive, costly, and impractical for fish below 20 g and for mass vaccination [117].
6.3. Protective Immune Response
The success of any vaccine against a parasite depends on the development of protective immunity in the fish host. The developed protective immune response should mimic as during a naturally occurring parasitic infection. Furthermore, the magnitude of immune response depends on the pathogen, intensity, and the stage of infection [118]. The protective immune responses of fish against some parasites are dependent on the production of specific antibodies. For example, specific antibodies were found in the sera of recovered and experimentally infected fish, which were confirmed to involve in protective immunity in fish against Cryptobia salmositica and Scuticociliates [119,120]. Anti-T. bryosalmonae specific antibodies have been detected in sera of experimentally infected brown trout from 4 to 17 weeks post exposure; however, studies on the mechanisms involved in protection are limited (authors own unpublished data). Partial antibody-mediated protection against I. multifiliis and Trypanosoma carassii was obtained after immunization of fish with recombinant proteins [121,122]. However, in several cases, the presence of specific antibodies were not correlated with protection of fish against parasites.
In the context of fish parasites, information on specific parasite stages that elicit immune response is mostly available for ectoparasites. For example, immunity is directed against the theront and trophont stages of I. multifiliis [123] and C. irritans [124]. Additionally, from metanauplius larvae to adult Argulus stages induce limited host response [18]. With regard to myxozoan endoparasites, the knowledge of host response on intrapiscine development of myxospores exists; however, specific stages capable of eliciting immune response is unknown. In cases of parasites exploiting the mucosal surface to invade the host, such as the myxozoans, the ectoparasites such as I. multifiliis and C. irritans, the hosts counter the invasion by initiating a mucosal immune response. Therefore, the prophylactic vaccines for such pathogens should be designed to stimulate mucosal immunity. Humoral components such as complement impart protection against P. dicentrarchi infection in turbot [125]. Both B cell and T cell responses confer protection in several infections, which depends on both the host and the parasite. Therefore, a vaccine for controlling P. dicentrarchi should activate the complement, whereas in the latter case should result in B cell- and T cell-mediated responses. Vaccines for fish parasites, e.g., E. leei and K. thyrsites, should induce T cell-mediated immune response, using vectors such as plasmid DNA or viruses [126]. Vaccines targeting secretory enzymes have shown promising results against the hookworm Ancyclostoma caninum [127] and the helminth Schistosoma mansoni [128]. A similar approach can be followed for the crustacean parasites e.g., L. salmonis, C. elongatus and Argulus which use many secretions as their immunesuppression strategy. Moreover, the nature of adjuvants affects the desired immune response. For example, cell-mediated immune response can be achieved by the administration of cytokines along with antigen [129] or by heterologous prime-boost approach, wherein the antigen is delivered sequentially using different vaccine platforms [130]. Oil-based adjuvants generate a high antibody titre in blood [113]. Another important aspect of consideration for a successful vaccination is the vaccine delivery, including the mode of administration, the dosage, and the timing.
6.4. Long-Term Immunity
Immunological memory underpins the concept of long-term immunity and vaccination. Vaccination-induced immune memory provides antibodies continuously and maintains memory cells to allow rapid response on exposure to the pathogen. The existence of immunological memory has been demonstrated in fish [131]. However, studies on its duration in response to parasitic infections in fish are limited. An epidemiological investigation on PKD in rainbow trout reported the presence of immunity in surviving fish during the subsequent year after infection [132]. Following exposure to theronts, channel catfish were found to be immune to re-infection after 3 years due to activation of memory B cells and mobilization of Iag-specific antigen-secreting cells into both systemic and mucosal compartments [133]. T cell-mediated immune response has been demonstrated during certain parasitic infections, e.g., T. bryosalmonae [134], E. leei [35], and sea lice [135]. In vertebrates, during the natural course of infection, following its control, the pathogen load along with T cell declines. The T cell survivors are responsible for immunological memory. Therefore, the extent of expansion and contraction along with the resulting memory depends on several factors such as the type and amount of antigen, the duration of antigen exposure, the site of antigen introduction, and the ability of the antigen or its co-delivered components to activate innate immune responses [93].
7. Challenges in Vaccine Development
Lack of knowledge of biology and the life cycle of parasites are the biggest challenges in developing vaccines against them. Although the life cycle aspects of a few parasites are relatively well understood [93,135], it is unknown for the majority of the fish parasites. Consequently, the cultivation and maintenance of parasites under laboratory conditions becomes difficult; however, it is a necessity for the preparation of antigens and the production of antibodies for vaccine development [136].
Another limitation is the immune escape mechanism of parasites that allows them to evade the direct interaction with the host’s immune system. Fish parasites have devised different strategies to escape the host immune defense during infection. For example, M. cerebralis invades the nerves, which is an immunologically privileged site with low host immune response [137]. Antigenic variation is an important feature of protozoan parasites that enables them to evade the host immune response and leading to chronic infections [136]. Antigenic variations of I. multifiliis have been confirmed by Northern hybridization, suggesting the expression of genes coding immobilization antigens in different life stages of I. multifiliis [138]. In addition, antigenic variations, reflected by nine putative I-antigens, have been reported in C. irritans from different life stages (tomont, theront, and trophont) using transcriptomic analysis [124]. Further, vaccine development is challenged by poor information on host–parasite interaction and immune response. As an example, although TLRs are known to play an important role in host defense against pathogens, including parasites, the ligand specificity is not yet determined for the majority of the TLRs. In addition, no information is available on different populations of cells in fish expressing TLRs. Such knowledge holds immense importance in the understanding of the resistance mechanisms in fish against parasites, and the development of novel adjuvants and more effective vaccines [139].
Presently, the lack of knowledge of specific antigens of parasites, which trigger the protective immune response is a major constraint. Large-scale production of recombinant proteins that retain the immunological activity similar to the natural parasite protein is an additional roadblock for the development of recombinant vaccines.
8. Role of Omics Technologies in Vaccine Development
Significant research in the study of parasites of human and veterinary importance highlight the potential of omics in the development of vaccines. Analysis of data obtained from omics-studies are an important source of information on SNPs, resistance markers, changes in gene expression, splicing variants, protein modifications, and strain-specificities of parasites. Such information helps in understanding biological attributes of the parasite that may aid in developing disease control strategies [140]. For example, characterization of proteins encoded by the polymorphic loci detected from genomic data of Plasmodium falciparum [141], can help in identification of vaccine targets [140]. Hypervariability of parasite antigens is viewed as a major obstacle in developing vaccine against them. Alternative splicing of parasite surface proteins is an important phenomenon that may result in isoforms differing in cell localization, substrate affinities and functions. The structural differences of the isoforms can be large enough to enable the parasite to evade host-immune recognition [142]. RNA-seq based transcriptomic studies have elucidated the role of alternative splicing during cellular differentiation in Plasmodium berghei [143]. The majority of the proteins that play pivotal roles in invasion are either stored in the apical secretory organelles or located on the surface of the merozoite, the invasive stage of the Plasmodium [144]. Novel secretory organelle proteins and surface-exposed proteins were identified from proteomic analysis of P. falciparum [145]. Additionally, an integrated transcriptomic and proteomic approach was applied to describe the Fasciola hepatica secretory proteome, thus identifying proteins such as cathepsin, peroxiredoxin, glutathione S-transferase, and fatty acid-binding proteins essential for the design of the first-generation anti-fluke vaccines and flukicidal drugs.
In view of the commonalities existing between mammalian and aquatic parasites (e.g., life cycle with multiple stages, multiple hosts and stage specific antigen profile) and considering the fact that research in mammalian parasitology is far ahead of aquatic parasitology, important lessons need to be learnt from both their failures and successes. The human endoparasites such as Plasmodium, Leishmania and Trypanosoma have been the focus of extensive research for many decades. It has been established by now that early efforts to develop effective vaccines against these parasites have failed as a result of a poor selection of few antigens without the knowledge of antigen repertoire of the parasites [146]. Past efforts have relied on single gene, transcript or protein in vaccine formulations. Likewise, as discussed earlier, vaccination trials against fish parasites have focused on either single or dual target vaccines resulting in limited or suboptimal protection. However, evidences from laboratory experiments [147] and field studies [148] on Plasmodium uphold the requirement of a multivalent vaccine wherein a robust immune response towards multiple antigenic determinants can be elicited to provide optimal protection the in host. A multivalent vaccine against human visceral leishmaniasis reportedly elicited significant humoral and cellular responses in pre-clinical trials [149]. Another multivalent vaccine containing stage-specific antigens of Fasciola hepatica conferred 83% protection in vaccinated rats [150]. Drawing from these studies from mammalian parasites, it is likely that a multivalent approach might be a better strategy against both endo- and ectoparasites of fish.
In fisheries and aquaculture, the application of omics has just begun, triggered by the advancement as well as cost reduction in NGS technologies [151]. Of the many omics techniques, the application of transcriptomics, genomics and proteomics are emerging in fish parasitology.
8.1. Transcriptomics
Transcriptomics-based studies have widely been used to explore the viral and bacterial pathogen–fish interactions [24]. Similar studies focusing on parasitic diseases in fish are limited. Yet, these restricted works on parasite-induced pathologies have elucidated important phenomena related to the host, such as immune response and concerning the parasite, possible escape strategies and functional biology. Recently, an RNA sequencing-based transcriptome study provided valuable insights into the immune response mounted by brown trout host, as well as modulated host machineries in response to T. bryosalmonae infection [152]. Additionally, transcriptome of bryozoan Fredericella sultana has been demonstrated, which provides valuable resources for the understanding of the unique biological characteristics and functional transcripts of this important bryozoan species that is the primary host of T. bryosalmonae [153]. Transcriptome analysis of T. bryosalmonae-infected F. sultana revealed 1166 differentially expressed genes with Eukaryotic Initiation Factor 2 signaling as a top canonical pathway and MYCN as a top upstream regulator [154]. Furthermore, the transcriptome of T. bryosalmonae from F. sultana is found to contain several members of the protease family, e.g., cathepsin L, cysteine protease, zinc metalloprotease, and serine protease [155]. The transcriptome analysis of E. scophthalmi-infected turbot suggested the role of IFN-mediated signaling pathways during incipient enteromyxosis as well as the downregulation of complement and acute phase proteins as possible immune evasion mechanisms [156]. De novo assembled transcriptome of Sphaerospora molnari blood stages are reported to contain 9436 proteins. This work has provided valuable information on a proteolytic depot of the parasite consisting 235 putative proteases, mainly of cysteine proteases [157]. Furthermore, genome and transcriptome-based data analysis of Kudoa iwatai and M. cerebralis was used to study the evolution of endoparasitism in myxozoans [158].
The comparative transcriptomic profile analysis of trophont, tomont and theront stages of C. irritans haselucidated the differentially expressed genes with functions in cell division, nutrition analysis and cell growth. Moreover, nine putative immobilization antigen (I-antigen) and protease transcripts, which can be considered as potential vaccine and drug targets were also found [159]. Argulus siamensis transcriptome examination revealed the presence of serine and metalloprotease that are known antigens in many ectoparasites. The study also led to the characterization of the downward signaling molecules of toll pathway [31].
8.2. Proteomics and Genomics
Proteomics and genomics are the other modern high-throughput techniques that have become instrumental in exploring different aspects of both the host and pathogen separately and their interactions at the molecular level. Kumar et al. [160] identified host–parasite protein interaction during proliferative kidney disease using antibody-based purification followed by electrospray ionization mass spectrometry. These identified proteins can be used for understanding the pathogenesis and defense mechanisms of T. bryosalmonae. Piriatinskiy et al. [161] identified polar capsule proteins of C. shasta using tandem mass spectrometry and suggested that polar capsules and nematocysts are homologous organelles. This study unraveled 112 proteins present in polar capsules of C. shasta with diverse functions such as the structural (nematogalectin and minicollagen), protein folding (HSP 70 and isomerases) enzymes involved in poly-γ-glutamate biosynthesis in addition to some novel proteins containing cysteine-rich and proline rich stretches.
Nano-LC ESI MS/MS based proteome analysis of I. multifiliis infected fish skin mucus revealed the involvement of innate immune components, e.g., lectins and serpins, in providing protection against the parasite [162]. An iTRAQ (Isobaric tags for relative and absolute quantitation) based quantitative proteomic study identified 2300 proteins in theront and trophont stages of I. multifiliis, of which 1520 proteins were differentially expressed in trophonts. These proteins played important roles of binding, catalytic activity, structural molecule activity and transporter activity in the parasite life-cycle [163]. The comparative proteomic analysis of theront, trophont, and tomont of C. irritans using 2D-gel electrophoresis and mass spectrophotometry identified different proteins, which could be used as vaccine candidates. Among these, α-tubulin and actin were found to be expressed in all the three developmental stages, whereas enolase was present in theront and trophont and vacuolar ATP synthase (V-ATPases) catalytic subunit α was detected only in theronts [163].
Genome sequencing and analysis of the transcriptionally active macronucleus of I. multifiliis has revealed several gene classes functioning in behavior, cellular functions, and host immunogenicity, including protein kinases, membrane transporters, proteases, and surface antigens, providing avenues for selecting vaccine and drug targets [164]. These gene families are identified as lead vaccine targets in many parasites of medical and veterinary importance. For example, members of the protein kinase family, e.g., Toxoplasma gondii calcium-dependent protein kinase 2 [165] and proteases [166] are considered promising vaccine candidates against the apicomplexan parasite Toxoplasma gondii that infects warm blooded animals [165]. Proteases are excellent shistostome vaccine targets as well. Membrane transport proteins are amongst the most attractive molecular targets of FDA-approved drugs [167].
Overall, the unprecedented wealth of information generated by the analysis of genomic, transcriptomic and proteomic data can better our knowledge of the host as well as the parasite. As discussed, omics can be significant in the identification of suitable antigen candidates of parasites; as such, it is important to note that the efficacy of previously reported vaccines can also be increased. This can be achieved by combining the already reported antigens with the ones selected based on omics data. For example, Jorgensen et al. [75] reported the immunogenicity of the recombinant protein #10; however, it is also noted that in combination with other suitable candidates, it can lead to better protection in fish on vaccination.
Additionally, advanced genome-based techniques can be significant in overcoming many issues concerning complex parasitic organisms, one of the important hurdles being the requirement of parasite culture. This is because maintaining a parasite life-cycle necessitates the maintenance of intermediate and final hosts, which are costly and exhaustive procedures requiring a large amount of time and effort. This is followed by antigen identification, isolation, and purification, which further involves several challenging steps. Besides, omics approaches will possibly be advantageous in cases where many genes and proteins are expressed only during the course of infection and where biomolecules are present in insufficient quantities to be recognized by assays. To date, sequence data are available for a limited number of ectoparasites and endoparasites (Table 4).
9. Approach of Multivalent Vaccines
The insights gained from omics data can be invaluable in the development of multivalent vaccines. A multivalent vaccine is a combination of several antigens to elicit a broad protective immune response by the host [173]. The different antigens can be selected from the same parasite, different parasites, parasite strains, and developmental stages. Considering the multiple critical problems associated with the aquatic parasites, particularly the diverse antigenic profile of the developmental stages and parasite strains, a multivalent vaccine, in general, would be an effective strategy against them. For certain parasites like T. bryosalmonae, although cell-mediated immune response is speculated to protect fish against infection [58], the exact protective response is not yet defined. Under these circumstances, multiple antigenic candidates can be selected and targeted based on studies on the parasite transcriptome, genome, and proteome along with host–parasite interaction data. Furthermore, a study with inactivated vaccine containing two different isolates of P. dicentrarchi did not confer cross-protection in turbot [65]. This could be explained by the fact that the parasite exhibits intraspecific variation both at the morphological and genetic levels [174]. The development of vaccines for such parasites would entail a pan-genomic approach wherein antigens conserved across genomes can be the target for a broad spectrum of protection. A conceptual representation of the multivalent vaccine using an omics dataset is presented in Figure 1.
In designing a multivalent vaccine, genomics, transcriptomics, structural genomics, proteomics, and immunoproteomics can be used to identify a suitable antigen. An approach for the selection of antigenic candidates and the development of vaccines utilizing omics approaches is presented in Figure 2. In this approach, whole-genome sequencing, RNA sequencing, and proteomics data can be used as a starting material of the targeted parasite or the infected host tissue. Further annotation of the data can be carried out using different bioinformatics tools such as Blast, Blast2GO, and UniProt/Swiss-Prot. After annotation, the gene, RNA transcripts, and proteins can be analyzed for their function as well as the different biological pathways. Vaccine candidates can be administered in fish to check the efficacy and potency of the vaccine.
10. Conclusions
There has been a recent increase in the outbreak of parasitic diseases in farmed and wild fish populations. The general practice for controlling parasitic infestations is to use chemotherapy. Over the past few decades, several concerns have been raised regarding their use, including environmental safety. The experts have recommended vaccines as effective solution to address these issues. Different types of vaccines have shown varying degrees of protection in fish against parasites. However, deeper insights into the host–parasite interaction and parasite’s life cycle with different stages are needed to be overcome for the development of successful vaccines. Research on some ectoparasites have been focused with the aim of developing vaccines. However, little attention has been given to the endoparasites. As an example, very little is known about the genes of myxozoans that are induced and expressed in both hosts (invertebrates and vertebrates) and involved in pathogenicity in the fish host during the course of infection. Hence, the identification of the in vivo induced genes of parasites related to disease development is required to improve our understanding of pathogenesis and promote the discovery of novel therapeutic targets. At present, transcriptome, genome, and proteomic data are limited for fish parasites. However, there is a need of large amount of omics data pertaining to more fish parasites for the identification of potential vaccine candidates and designing multivalent vaccines.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2076-393X/9/2/179/s1, Figure S1: PRISMA flow diagram of study search and selection.
Author Contributions
G.K. and M.E.-M. conceived the idea and supervised the manuscript. S.S. compiled the information and drafted the paper. G.K. and M.E.-M. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Austrian Science Fund (FWF) Project Number P 30981-B32 to G.K.
Acknowledgments
We are thankful to the Austrian Science Fund (FWF) Project Number P 30981-B32 for providing Open Access Funding. S.S. is thankful to the Indian Council of Agricultural Research, New Delhi, India for providing her with the scholarship to pursue PhD under Netaji Subhas ICAR International Fellowship 2018–2019.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Murray, A.G.; Peeler, E.J. A framework for understanding the potential for emerging diseases in aquaculture. Prev. Vet. Med. 2005, 67, 223–235. [Google Scholar] [CrossRef]
- Shinn, A.P.; Pratoomyot, J.; Bron, J.E.; Paladini, G.; Brooker, E.E.; Brooker, A.J. Economic costs of protistan and metazoan parasites to global mariculture. Parasitology 2015, 142, 196–270. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H. A review: Gaps in our knowledge on myxozoan parasites of fishes. Fish Pathol. 2003, 38, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, G.L. Myxobolus cerebralis, a worldwide cause of salmonid whirling disease. J. Aquat. Anim. Health 1990, 2, 30–37. [Google Scholar] [CrossRef]
- Bravo, S.; Sevatdal, S.; Horsberg, T.E. Sensitivity assessment of Caligus rogercresseyi to emamectin benzoate in Chile. Aquaculture 2008, 282, 7–12. [Google Scholar] [CrossRef]
- Sevatdal, S.; Copley, L.; Wallace, C.; Jackson, D.; Horsberg, T.E. Monitoring of the sensitivity of sea lice (Lepeophtheirus salmonis) to pyrethroids in Norway, Ireland and Scotland using bioassays and probit modelling. Aquaculture 2005, 244, 19–27. [Google Scholar] [CrossRef]
- Boxall, A.B.A. The environmental side effects of medication: How are human and veterinary medicines in soils and water bodies affecting human and environmental health? EMBO Rep. 2004, 5, 1110–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.C.; Jürgens, M.D.; Williams, R.J.; Kümmerer, K.; Kortenkamp, A.; Sumpter, J.P. Do cytotoxic chemotherapy drugs discharged into rivers pose a risk to the environment and human health? An overview and UK case study. J. Hydrol. 2008, 348, 167–175. [Google Scholar] [CrossRef]
- Valladão, G.M.R.; Gallani, S.U.; Pilarski, F. Phytotherapy as an alternative for treating fish disease. J. Vet. Pharmacol. Ther. 2015, 38, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Neurol. 2014, 4, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, H.; Ogawa, K.; Wakabayashi, H. Chemotherapy with fumagillin and toltrazuril against kidney enlargement disease of goldfish caused by the myxosporean Hoferellus carassii. Fish. Pathol. 1990, 25, 157–163. [Google Scholar] [CrossRef]
- Morris, D.J.; Adams, A.; Smith, P.; Richards, R.H. Effects of oral treatment with TNP-470 on rainbow trout (Oncorhynchus mykiss) infected with Tetracapsuloides bryosalmonae (Malacosporea), the causative agent of proliferative kidney disease. Aquaculture 2003, 221, 51–64. [Google Scholar] [CrossRef]
- Flamarique, I.N.; Gulbransen, C.; Galbraith, M.; Stucchi, D. Monitoring and potential control of sea lice using an LED-based light trap. Can. J. Fish. Aquat. Sci. 2009, 66, 1371–1382. [Google Scholar] [CrossRef]
- Skilton, D.C.; Saunders, R.J.; Hutson, K.S. Parasite attractants: Identifying trap baits for parasite management in aquaculture. Aquaculture 2020, 516, 734557. [Google Scholar] [CrossRef]
- Adams, A. Progress, challenges and opportunities in fish vaccine development. Fish Shellfish Immunol. 2019, 90, 210–214. [Google Scholar] [CrossRef]
- Villegas, A. First Ever Sea Lice Vaccine Launched in Chile. Available online: https://www.undercurrentnews.com/2015/11/24/first-ever-sea-lice-vaccine-launched-in-chile/ (accessed on 26 February 2020).
- Woo, P.T.K.; Buchmann, K. Fish Parasites:Pathobiology and Protection; CAB International: Wallingford, UK, 2012; ISBN 9781845938062. [Google Scholar]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kavallaris, M.; Marshall, G.M. Proteomics and disease: Opportunities and challenges. Med. J. Aust. 2005, 182, 575–579. [Google Scholar] [CrossRef]
- Hegde, P.S.; White, I.R.; Debouck, C. Interplay of transcriptomics and proteomics. Curr. Opin. Biotechnol. 2003, 14, 647–651. [Google Scholar] [CrossRef]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef] [Green Version]
- Feder, M.E.; Walser, J.C. The biological limitations of transcriptomics in elucidating stress and stress responses. J. Evol. Biol. 2005, 18, 901–910. [Google Scholar] [CrossRef]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. Transcriptome analysis based on RNA-Seq in understanding pathogenic mechanisms of diseases and the immune system of fish: A comprehensive review. Int. J. Mol. Sci. 2018, 19, 245. [Google Scholar] [CrossRef] [Green Version]
- Feist, S.W.; Longsaw, M. Phylum myxozoa. In Fish Diseases and Disorders; Woo, P.T.K., Ed.; CAB International: Wallingford, UK, 2006; pp. 230–296. [Google Scholar]
- Clifton-Hadley, R.S.; Richards, R.H.; Bucke, D. Proliferative kidney disease (PKD) in rainbow trout Salmo gairdneri: Further observations on the effects of water temperature. Aquaculture 1986, 55, 165–171. [Google Scholar] [CrossRef]
- Rognlie, M.C.; Knapp, S.E. Myxobolus cerebralis in Tubifex tubifex from a Whirling Disease Epizootic in Montana. J. Parasitol. 1998, 84, 711–713. [Google Scholar]
- Lom, J.; Dyková, I.; Pavlásková, M.; Grupcheva, G. Sphaerospora molnari sp.nov. (Myxozoa:Myxosporea) an Agent of Gill, Skin and Blood Sphaerosporosis of Common Carp in Europe. Parasitology 1983, 86, 529–535. [Google Scholar] [CrossRef]
- Körting, W. Larval development of Bothriocephalus sp. (Cestoda: Pseudophyllidea) from carp (Cyprinm carpio L.) in Germany. J. Fish Biol. 1975, 7, 727–733. [Google Scholar] [CrossRef]
- Matthews, R.A. Ichthyophthirius multifiliis fouquet and ichthyophthiriosis in freshwater teleosts. Adv. Parasitol. 2005, 59, 159–241. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, P.K.; Kar, B.; Mohapatra, A.; Mohanty, J. De novo whole transcriptome analysis of the fish louse, Argulus siamensis: First molecular insights into characterization of Toll downstream signalling molecules of crustaceans. Exp. Parasitol. 2013, 135, 629–641. [Google Scholar] [CrossRef]
- Bakke, T.A.; Cable, J.; Harris, P.D. The Biology of Gyrodactylid Monogeneans: The “Russian-Doll Killers”. Adv. Parasitol. 2007, 64. [Google Scholar] [CrossRef]
- Braden, L.M.; Rasmussen, K.J.; Purcell, S.L.; Ellis, L.; Mahony, A.; Cho, S.; Whyte, S.K.; Jones, S.R.M.; Fast, M.D. Acquired protective immunity in Atlantic salmon Salmo salar against the myxozoan Kudoa thyrsites involves induction of MHIIβ+ CD83+ antigen-presenting cells. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Kent, M.L.; Margolis, L.; Whitaker, D.J.; Hoskins, G.E.; McDonald, T.E. Review of Myxosporea of importance in salmonid fisheries and aquaculture in British Columbia. Folia Parasitol. 1994, 41, 27–37. [Google Scholar]
- Piazzon, M.C.; Estensoro, I.; Calduch-Giner, J.A.; Del Pozo, R.; Picard-Sánchez, A.; Pérez-Sánchez, J.; Sitjà-Bobadilla, A. Hints on T cell responses in a fish-parasite model: Enteromyxum leei induces differential expression of T cell signature molecules depending on the organ and the infection status. Parasites Vectors 2018, 11, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzon, M.C.; Leiro, J.; Lamas, J. Reprint of “Fish immunity to scuticociliate parasites”. Dev. Comp. Immunol. 2014, 43, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.C.; Treasurer, J.W.; Bravo, S.; Nagasawa, K.; Kabata, Z. A review of the impact of parasitic copepods on marine aquaculture. Zool. Stud. 2004, 43, 229–243. [Google Scholar]
- Jones, S.R.M. The occurrence and mechanisms of innate immunity against parasites in fish. Dev. Comp. Immunol. 2001, 25, 841–852. [Google Scholar] [CrossRef]
- Sitja-Bobadilla, A.; Palenzuela, O. Enteromyxum species. In Fish Parasites: Pathobiology and Protection; Woo, P.T.K., Buchmann, K., Eds.; CAB International: Oxfordshire, UK, 2012; p. 163. [Google Scholar]
- Frazer, L.N. Sea-cage aquaculture, sea lice, and declines of wild fish. Conserv. Biol. 2009, 23, 599–607. [Google Scholar] [CrossRef]
- Moller, O.S. Argulus foliaceus. In Fish Parasites: Pathobiology and Protection; Woo, P.T.K., Buchmann, K., Eds.; CAB International: Oxfordshire, UK, 2012; pp. 327–330. [Google Scholar]
- Bartholomew, J.L.; Reno, P.W. The history and dissemination of whirling disease. Am. Fish. Soc. Symp. 2002, 2002, 3–24. [Google Scholar]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. The malacosporean myxozoan parasite Tetracapsuloides bryosalmonae: A threat to wild salmonids. Pathogens 2020, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Hallett, S.L.; Ray, R.A.; Hurst, C.N.; Holt, R.A.; Buckles, G.R.; Atkinson, S.D.; Bartholomew, J.L. Density of the waterborne parasite Ceratomyxa shasta and its biological effects on salmon. Appl. Environ. Microbiol. 2012, 78, 3724–3731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickerson, H.W.; Findly, R.C. Immunity to Ichthyophthirius infections in fish: A synopsis. Dev. Comp. Immunol. 2014, 43, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Barker, S.E.; Bricknell, I.R.; Covello, J.; Purcell, S.; Fast, M.D.; Wolters, W.; Bouchard, D.A. Sea lice, Lepeophtheirus salmonis (Krøyer 1837), infected Atlantic salmon (Salmo salar L.) are more susceptible to infectious salmon anemia virus. PLoS ONE 2019, 14, e0209178. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Rauta, P.R.; Nayak, B.; Das, S. Immune system and immune responses in fish and their role in comparative immunity study: A model for higher organisms. Immunol. Lett. 2012, 148, 23–33. [Google Scholar] [CrossRef]
- Picard-Sánchez, A.; Estensoro, I.; del Pozo, R.; Piazzon, M.C.; Palenzuela, O.; Sitjà-Bobadilla, A. Acquired protective immune response in a fish-myxozoan model encompasses specific antibodies and inflammation resolution. Fish Shellfish Immunol. 2019, 90, 349–362. [Google Scholar] [CrossRef]
- Rigos, G.; Pavlidis, M.; Divanach, P. Host susceptibility to Cryptocaryon sp. infection of Mediterranean marine broodfish held under intensive culture conditions: A case report. Bull. Eur. Assoc. Fish. Pathol. 2001, 21, 33–36. [Google Scholar]
- Takano, T.; Kondo, H.; Hirono, I.; Endo, M.; Saito-Taki, T.; Aoki, T. Toll-like receptors in teleosts. In Diseases in Asian Aquaculture VII; Fish Health Section, Asian Fisheries Society: Selangor, Malaysia, 2011; Volume 385, pp. 197–208. [Google Scholar]
- Sudhagar, A.; El-Matbouli, M.; Kumar, G. Identification and expression profiling of toll-like receptors of brown trout (Salmo trutta) during proliferative kidney disease. Int. J. Mol. Sci. 2020, 21, 3755. [Google Scholar] [CrossRef]
- Zhao, F.; Li, Y.W.; Pan, H.J.; Shi, C.B.; Luo, X.C.; Li, A.X.; Wu, S.Q. Expression profiles of toll-like receptors in channel catfish (Ictalurus punctatus) after infection with Ichthyophthirius multifiliis. Fish Shellfish Immunol. 2013, 35, 993–997. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.W.; Xu, D.D.; Li, X.; Mo, Z.Q.; Luo, X.C.; Li, A.X.; Dan, X.M. Identification and characterization of three TLR1 subfamily members from the orange-spotted grouper, Epinephelus coioides. Dev. Comp. Immunol. 2016, 61, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Muñoz, V.; Boltaña, S.; Gallardo-Escárate, C. Comparative immunity of Salmo salar and Oncorhynchus kisutch during infestation with the sea louse Caligus rogercresseyi: An enrichment transcriptome analysis. Fish Shellfish Immunol. 2016, 59, 276–287. [Google Scholar] [CrossRef]
- Becker, I.; Salaiza, N.; Aguirre, M.; Delgado, J.; Carrillo-Carrasco, N.; Kobeh, L.G.; Ruiz, A.; Cervantes, R.; Torres, A.P.; Cabrera, N.; et al. Leishmania lipophosphoglycan (LPG) activates NK cells through toll-like receptor. Mol. Biochem. Parasitol. 2003, 130, 65–74. [Google Scholar] [CrossRef]
- Sepulcre, M.; Pelegrín, P.; Mulero, V.; Meseguer, J. Characterisation of gilthead seabream acidophilic granulocytes by a monoclonal antibody unequivocally points to their involvement in fish phagocytic response. Cell Tissue Res. 2002, 308, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Palikova, M.; Papezikova, I.; Markova, Z.; Navratil, S.; Mares, J.; Mares, L.; Vojtek, L.; Hyrsl, P.; Jelinkova, E.; Schmidt-Posthaus, H. Proliferative kidney disease in rainbow trout (Oncorhynchus mykiss) under intensive breeding conditions: Pathogenesis and haematological and immune parameters. Vet. Parasitol. 2017, 238, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Álvarez-Pellitero, P.; Sitja-Bobadilla, A. Modulation of the in vitro activity of European sea bass (Dicentrarchus labrax L.) phagocytes by the myxosporean parasite Sphaerospora dicentrarchi (Myxosporea: Bivalvulida). Fish Shellfish Immunol. 2000, 10, 567–581. [Google Scholar] [CrossRef]
- Chin, A.; Woo, P.T.K. Innate cell-mediated immune response and peripheral leukocyte populations in Atlantic salmon, Salmo salar L.; to a live Cryptobia salmositica vaccine. Parasitol. Res. 2005, 95, 299–304. [Google Scholar] [CrossRef]
- Korytář, T.; Wiegertjes, G.F.; Zusková, E.; Tomanová, A.; Lisnerová, M.; Patra, S.; Sieranski, V.; Šíma, R.; Born-Torrijos, A.; Wentzel, A.S.; et al. The kinetics of cellular and humoral immune responses of common carp to presporogonic development of the myxozoan Sphaerospora molnari. Parasites Vectors 2019, 12, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, S.S.; Evans, D.L.; Dawe, D.L. Mobilization and activation of nonspecific cytotoxic cells (ncc) in the channel catfish (Ictalurus punctatus) infected with Ichthyophthirius multifiliis. Comp. Immunol. Microbiol. Infect. Dis. 1985, 8, 43–51. [Google Scholar] [CrossRef]
- Mali, P.; Sanyal, K.B.; Mukherjee, D.; Guchhait, A.; Dash, G. Nonspecific cytotoxic cells (NCC) in fish: A review. J. Interacad. 2017, 21, 372–378. [Google Scholar]
- Jaso-Friedmann, L.; Leary, J.H.; Weisman, Z.; Evans, D.L. Activation of nonspecific cytotoxic cells with a multiple antigenic peptide: Specificity and requirements for receptor crosslinkage. Cell. Immunol. 1996, 170, 195–201. [Google Scholar] [CrossRef]
- Leiro, J.; Piazzón, M.C.; Budiño, B.; Sanmartín, M.L.; Lamas, J. Complement-mediated killing of Philasterides dicentrarchi (Ciliophora) by turbot serum: Relative importance of alternative and classical pathways. Parasite Immunol. 2008, 30, 535–543. [Google Scholar] [CrossRef]
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef]
- Cuesta, A.; Muñoz, P.; Rodríguez, A.; Salinas, I.; Sitjà-Bobadilla, A.; Álvarez-Pellitero, P.; Esteban, M.A.; Meseguer, J. Gilthead seabream (Sparus aurata L.) innate defence against the parasite Enteromyxum leei (Myxozoa). Parasitology 2006, 132, 95–104. [Google Scholar] [CrossRef]
- Holzer, A.S.; Schachner, O. Myxosporidia and macrophage centres in chub (Leuciscus cephalus)-Quantitative interactions focus on Myxobolus cyprini. Parasitology 2001, 122, 55–62. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Redondo, M.J.; Bermúdez, R.; Palenzuela, O.; Ferreiro, I.; Riaza, A.; Quiroga, I.; Nieto, J.M.; Alvarez-Pellitero, P. Innate and adaptive immune responses of turbot, Scophthalmus maximus (L.), following experimental infection with Enteromyxum scophthalmi (Myxosporea: Myxozoa). Fish Shellfish Immunol. 2006, 21, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Davey, G.C.; Calduch-Giner, J.A.; Houeix, B.; Talbot, A.; Sitjà-Bobadilla, A.; Prunet, P.; Pérez-Sánchez, J.; Cairns, M.T. Molecular profiling of the gilthead sea bream (Sparus aurata L.) response to chronic exposure to the myxosporean parasite Enteromyxum leei. Mol. Immunol. 2011, 48, 2102–2112. [Google Scholar] [CrossRef] [Green Version]
- Baerwald, M.R. Temporal expression patterns of rainbow trout immune-related genes in response to Myxobolus cerebralis exposure. Fish Shellfish Immunol. 2013, 35, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Bjork, S.J.; Zhang, Y.A.; Hurst, C.N.; Alonso-Naveiro, M.E.; Alexander, J.D.; Sunyer, J.O.; Bartholomew, J.L. Defenses of susceptible and resistant Chinook salmon (Onchorhynchus tshawytscha) against the myxozoan parasite Ceratomyxa shasta. Fish Shellfish Immunol. 2014, 37, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wangkahart, E.; Secombes, C.J.; Wang, T. Insights into the evolution of the suppressors of cytokine signaling (SOCS) gene family in vertebrates. Mol. Biol. Evol. 2019, 36, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Kotob, M.H.; Kumar, G.; Saleh, M.; Gorgoglione, B.; Abdelzaher, M.; El-Matbouli, M. Differential modulation of host immune genes in the kidney and cranium of the rainbow trout (Oncorhynchus mykiss) in response to Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections. Parasites Vectors 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, L.v.G. The fish parasite Ichthyophthirius multifiliis—Host immunology, vaccines and novel treatments. Fish Shellfish Immunol. 2017, 67, 586–595. [Google Scholar] [CrossRef]
- Mustafa, A.; MacWilliams, C.; Fernandez, N.; Matchett, K.; Conboy, G.A.; Burka, J.F. Effects of sea lice (Lepeophtheirus salmonis Kröyer, 1837) infestation on macrophage functions in Atlantic salmon (Salmo salar L.). Fish Shellfish Immunol. 2000, 10, 47–59. [Google Scholar] [CrossRef]
- Dalvin, S.; Jørgensen, L.v.G.; Kania, P.W.; Grotmol, S.; Buchmann, K.; Øvergård, A.C. Rainbow trout Oncorhynchus mykiss skin responses to salmon louse Lepeophtheirus salmonis: From copepodid to adult stage. Fish Shellfish Immunol. 2020, 103, 200–210. [Google Scholar] [CrossRef]
- Wallace, J.L. Nitric oxide as a regulator of inflammatory processes. Mem. Inst. Oswaldo Cruz 2005, 100, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Salinas, I.; Zhang, Y.A.; Sunyer, J.O. Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol. 2011, 35, 1346–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saulnier, D.; De Kinkelin, P. Antigenic and biochemical study of PKX, the myxosporean causative agent of proliferative kidney disease of salmonid fish. Dis. Aquat. Organ. 1996, 27, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.; Abd-Elfattah, A.; El-Matbouli, M. Differential modulation of host genes in the kidney of brown trout Salmo trutta during sporogenesis of Tetracapsuloides bryosalmonae (Myxozoa). Vet. Res. 2014, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, R.P.; El-Matbouli, M.; Adkison, M.A.; MacConnell, E. Whirling disease: Re-emergence among wild trout. Immunol. Rev. 1998, 166, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Palenzuela, O.; Sitjà-Bobadilla, A.; Álvarez-Pellitero, P. Immunohistochemical reactivity of polyclonal antibodies against Sphaerospora testicularis and Ceratomyxa labracis (Myxosporea: Bivalvulida), with other myxosporean parasites. Int. J. Parasitol. 1999, 29, 521–525. [Google Scholar] [CrossRef]
- Bartholomew, J.L. Host resistance to infection by the myxosporean parasite Ceratomyxa shasta: A review. J. Aquat. Anim. Health 1998, 10, 112–120. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Redondo, M.J.; Macias, M.A.; Ferreiro, I.; Riaza, A.; Alvarez-Pellitero, P. Development of immunohistochemistry and enzyme-linked immunosorbent assays for the detection of circulating antibodies against Enteromyxum scophthalmi (Myxozoa) in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2004, 17, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, X.; Sato, H.; Zhang, Q.; Li, A.; Zhang, J. RNA-seq analysis of local tissue of Carassius auratus gibelio with pharyngeal myxobolosis: Insights into the pharyngeal mucosal immune response in a fish-parasite dialogue. Fish Shellfish Immunol. 2019, 94, 99–112. [Google Scholar] [CrossRef]
- Zhang, Y.A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; Lapatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Parra, D.; Gómez, D.; Salinas, I.; Zhang, Y.A.; Von Gersdorff Jørgensen, L.; Heinecke, R.D.; Buchmann, K.; LaPatra, S.; Oriol Sunyer, J. Teleost skin, an ancient mucosal surface that elicits gut-like immune responses. Proc. Natl. Acad. Sci. USA 2013, 110, 13097–13102. [Google Scholar] [CrossRef] [Green Version]
- Abos, B.; Estensoro, I.; Perdiguero, P.; Faber, M.; Hu, Y.; Rosales, P.D.; Granja, A.G.; Secombes, C.J.; Holland, J.W.; Tafalla, C. Dysregulation of B cell activity during proliferative kidney disease in rainbow trout. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKinnon, B.M. Host Response of Atlantic Salmon (Salmo salar) to Infection by Sea Lice (Caligus elongatus). Can. J. Fish. Aquat. Sci. 1993, 50, 789–792. [Google Scholar] [CrossRef]
- Nakanishi, T.; Shibasaki, Y.; Matsuura, Y. T cells in fish. Biology 2015, 4, 640–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, M.; Montero, R.; Kumar, G.; Sudhagar, A.; Friedl, A.; Köllner, B.; El-Matbouli, M. Kinetics of local and systemic immune cell responses in whirling disease infection and resistance in rainbow trout. Parasites Vectors 2019, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarleton, R.L. New approaches in vaccine development for parasitic infections. Cell. Microbiol. 2005, 7, 1379–1386. [Google Scholar] [CrossRef]
- Clem, A.S. Fundamentals of vaccine immunology. J. Glob. Infect. Dis. 2011, 3, 73–78. [Google Scholar] [CrossRef]
- Goven, B.A.; Dawe, D.L.; Gratzek, J.B. Protection of channel catfish, Ictalurus punctatus Rafmesque, against Ichthyophthirius multifiliis Fouquet by immunization. J. Fish. Biol. 1980, 17, 311–316. [Google Scholar] [CrossRef]
- Gibbons, I.R. Studies on the Protein Components of Cilia From Tetrahymena pyriformis. Proc. Natl. Acad. Sci. USA 1963, 50, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Sleigh, M.A. Cilia and Flagella; Academic Press: London, UK, 1974. [Google Scholar]
- Burkart, M.A.; Clark, T.G.; Dickerson, H.W. Immunization of channel catfish, Ictalurus punctatus Rafinesque, against Ichthyophthirius multifiliis (Fouquet): Killed versus live vaccines. J. Fish Dis. 1990, 13, 401–410. [Google Scholar] [CrossRef]
- Xu, D.H.; Zhang, D.; Shoemaker, C.; Beck, B. Immune response of channel catfish (Ictalurus punctatus) against Ichthyophthirius multifiliis post vaccination using DNA vaccines encoding immobilization antigens. Fish Shellfish Immunol. 2019, 94, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.H.; Klesius, P.H. Comparison of serum antibody responses and host protection against parasite Ichthyophthirius multifiliis between channel catfish and channel × blue hybrid catfish. Fish Shellfish Immunol. 2013, 34, 1356–1359. [Google Scholar] [CrossRef]
- Von Gersdorff Jørgensen, L.; Kania, P.W.; Rasmussen, K.J.; Mattsson, A.H.; Schmidt, J.; Al-Jubury, A.; Sander, A.; Salanti, A.; Buchmann, K. Rainbow trout (Oncorhynchus mykiss) immune response towards a recombinant vaccine targeting the parasitic ciliate Ichthyophthirius multifiliis. J. Fish. Dis. 2017, 40, 1815–1821. [Google Scholar] [CrossRef] [Green Version]
- von Gersdorff Jørgensen, L.; Sigh, J.; Kania, P.W.; Holten-Andersen, L.; Buchmann, K.; Clark, T.; Rasmussen, J.S.; Einer-Jensen, K.; Lorenzen, N. Approaches towards DNA Vaccination against a Skin Ciliate Parasite in Fish. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Jørgensen, L.V.G.; Nemli, E.; Heinecke, R.D.; Raida, M.K.; Buchmann, K. Immune-relevant genes expressed in rainbow trout following immunisation with a live vaccine against Ichthyophthirius multifiliis. Dis. Aquat. Organ. 2008, 80, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Dalgaard, M.; Buchmann, K.; Li, A. Immunization of rainbow trout fry with Ichthyophthirius multifiliis sonicate: Protection of host and immunological changes. Bull. Eur. Assoc. Fish Pathol. 2002, 22, 288–297. [Google Scholar]
- Xu, D.H.; Klesius, P.H.; Shoemaker, C.A. Protective immunity of Nile tilapia against Ichthyophthirius multifiliis post-immunization with live theronts and sonicated trophonts. Fish Shellfish Immunol. 2008, 25, 124–127. [Google Scholar] [CrossRef]
- Yambot, A.V.; Song, Y.L. Immunization of grouper, Epinephelus coioides, confers protection against a protozoan parasite, Cryptocaryon irritans. Aquaculture 2006, 260, 1–9. [Google Scholar] [CrossRef]
- Grayson, T.H.; John, R.J.; Wadsworth, S.; Greaves, K.; Cox, D.; Roper, J.; Wrathmell, A.B.; Gilpin, M.L.; Harris, J.E. Immunization of Atlantic salmon against the salmon louse: Identification of antigens and effects on louse fecundity. J. Fish Biol. 1995, 47, 85–94. [Google Scholar] [CrossRef]
- Faber, M.N.; Holland, J.W.; Secombes, C.J. Vaccination strategies and IgM responses against PKD in rainbow trout. Fish Shellfish Immunol. 2019, 91, 423. [Google Scholar] [CrossRef]
- Kane, S.N.; Mishra, A.; Dutta, A.K. Preface: International Conference on Recent Trends in Physics (ICRTP 2016). J. Phys. Conf. Ser. 2016, 755. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Poly d,l-lactide-co-glycolic acid (PLGA)-encapsulated vaccine on immune system in Epinephelus bruneus against Uronema marinum. Exp. Parasitol. 2012, 131, 325–332. [Google Scholar] [CrossRef]
- Fontenla, F.; Blanco-Abad, V.; Pardo, B.G.; Folgueira, I.; Noia, M.; Gómez-Tato, A.; Martínez, P.; Leiro, J.M.; Lamas, J. Vaccine-induced modulation of gene expression in turbot peritoneal cells. A microarray approach. Mol. Immunol. 2016, 75, 188–199. [Google Scholar] [CrossRef]
- Méndez, S.; Gurunathan, S.; Kamhawi, S.; Belkaid, Y.; Moga, M.A.; Skeiky, Y.A.W.; Campos-Neto, A.; Reed, S.; Seder, R.A.; Sacks, D. The Potency and Durability of DNA- and Protein-Based Vaccines Against Leishmania major Evaluated Using Low-Dose, Intradermal Challenge. J. Immunol. 2001, 166, 5122–5128. [Google Scholar] [CrossRef] [Green Version]
- Knox, D.P.; Redmond, D.L. Parasite vaccines-Recent progress and problems associated with their development. Parasitology 2006, 133. [Google Scholar] [CrossRef]
- Mutoloki, S.; Munang’andu, H.M.; Evensen, Ø. Oral vaccination of fish-antigen preparations, uptake, and immune induction. Front. Immunol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bøgwald, J.; Dalmo, R.A. Review on immersion vaccines for fish: An update. Microorganisms 2019, 7, 627. [Google Scholar] [CrossRef] [Green Version]
- Plant, K.P.; LaPatra, S.E. Advances in fish vaccine delivery. Dev. Comp. Immunol. 2011, 35, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Horne, M.T. Technical aspects of the administration of vaccines. Dev. Biol. Stand. 1997, 90, 79–89. [Google Scholar]
- Sitjà-Bobadilla, A.; Estensoro, I.; Pérez-Sánchez, J. Immunity to gastrointestinal microparasites of fish. Dev. Comp. Immunol. 2016, 64, 187–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.T.K. Cryptobia (Trypanoplasma) salmositica and salmonid cryptobiosis. J. Fish. Dis. 2003, 26, 627–646. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Palenzuela, O.; Alvarez-Pellitero, P. Immune response of turbot, Psetta maxima (L.) (Pisces: Teleostei), to formalin-killed scuticociliates (Ciliophora) and adjuvanted formulations. Fish Shellfish Immunol. 2008, 24, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchmann, K.; Lindenstrøm, T.; Bresciani, J. Defence mechanisms against parasites in fish and the prospect for vaccines. Acta Parasitol 2001, 46, 76–81. [Google Scholar]
- Katzenback, B.A.; Plouffe, D.A.; Haddad, G.; Belosevic, M. Administration of recombinant parasite β-tubulin to goldfish (Carassius auratus L.) confers partial protection against challenge infection with Trypanosoma danilewskyi Laveran and Mesnil. Vet. Parasitol. 2008, 151, 36–45. [Google Scholar] [CrossRef]
- Buchmann, K. Immune response to Ichthyophthirius multifiliis and role of IgT. Parasite Immunol. 2020, 42, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Mo, Z.Q.; Li, Y.W.; Wang, H.Q.; Wang, J.L.; Ni, L.Y.; Yang, M.; Lao, G.F.; Luo, X.C.; Li, A.X.; Dan, X.M. Comparative transcriptional profile of the fish parasite Cryptocaryon irritans. Parasites Vectors 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzon, M.C.; Wiegertjes, G.F.; Leiro, J.; Lamas, J. Turbot resistance to Philasterides dicentrarchi is more dependent on humoral than on cellular immune responses. Fish Shellfish Immunol. 2011, 30, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.L.; Khader, S.A. Novel vaccine approaches for protection against intracellular pathogens. Curr. Opin. Immunol. 2014, 28, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Bethony, J.; Loukas, A.; Smout, M.; Brooker, S.; Mendez, S.; Plieskatt, J.; Goud, G.; Bottazzi, M.E.; Zhan, B.; Wang, Y.; et al. Antibodies against a secreted protein from hookworm larvae reduce the intensity of hookworm infection in humans and vaccinated laboratory animals. FASEB J. 2005, 19, 1743–1745. [Google Scholar] [CrossRef] [PubMed]
- Riveau, G.; Deplanque, D.; Remoué, F.; Schacht, A.M.; Vodougnon, H.; Capron, M.; Thiry, M.; Martial, J.; Libersa, C.; Capron, A. Safety and immunogenicity of rSh28GST antigen in humans: Phase 1 randomized clinical study of a vaccine candidate against urinary schistosomiasis. PLoS Negl. Trop. Dis. 2012, 6, e1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lofthouse, S.A.; Andrews, A.E.; Elhay, M.J.; Bowles, V.M.; Meeusen, E.N.T.; Nash, A.D. Cytokines as adjuvants for ruminant vaccines. Int. J. Parasitol. 1996, 26, 835–842. [Google Scholar] [CrossRef]
- Brown, G.V.; Reeder, J.C. Malaria vaccines. Med. J. Aust. 2002, 177, 230–231. [Google Scholar] [CrossRef] [PubMed]
- Arkoosh, M.R.; Kaattari, S.L. Development of immunological memory in rainbow trout (Oncorhynchus mykiss). I. An immunochemical and cellular analysis of the B cell response. Dev. Comp. Immunol. 1991, 15, 279–293. [Google Scholar] [CrossRef]
- Ferguson, H.W.; BalL, H.J. Epidemiological aspects of proliferative kidney disease amongst rainbow trout Salmo gairdneri Richardson in Northern Ireland. J. Fish Dis. 1979, 2, 219–225. [Google Scholar] [CrossRef]
- Findly, R.C.; Zhao, X.; Noe, J.; Camus, A.C.; Dickerson, H.W. B cell memory following infection and challenge of channel catfish with Ichthyophthirius multifiliis. Dev. Comp. Immunol. 2013, 39, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Gorgoglione, B.; Wang, T.; Secombes, C.J.; Holland, J.W. Immune gene expression profiling of Proliferative Kidney Disease in rainbow trout Oncorhynchus mykiss reveals a dominance of anti-inflammatory, antibody and T helper cell-like activities. Vet. Res. 2013, 44, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fast, M.D. Fish immune responses to parasitic copepod (namely sea lice) infection. Dev. Comp. Immunol. 2014, 43, 300–312. [Google Scholar] [CrossRef]
- Dzikowski, R.; Deitsch, K. Antigenic variation by protozoan parasites: Insights from Babesia bovis. Mol. Microbiol. 2006, 59, 364–366. [Google Scholar] [CrossRef]
- El-Matbouli, M.; Hoffmann, R.W.; Schoel, H.; McDowell, T.S.; Hedrick, R.P. Whirling disease: Host specificity and interaction between the actinosporean stage of Myxobolus cerebralis and rainbow trout Oncorhynchus mykiss. Dis. Aquat. Organ. 1999, 35, 1–12. [Google Scholar] [CrossRef]
- Clark, T.G.; Mcgraw, R.A.; Dickerson, H.W. Developmental expression of surface antigen genes in the parasitic ciliate Ichthyophthirius multifiliis. Proc. Natl. Acad. Sci. USA 1992, 89, 6363–6367. [Google Scholar] [CrossRef] [Green Version]
- Palti, Y. Toll-like receptors in bony fish: From genomics to function. Dev. Comp. Immunol. 2011, 35, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Nóbrega de Sousa, T.; de Menezes Neto, A.; Alves de Brito, C.F. “Omics” in the study of the major parasitic diseases malaria and schistosomiasis. Infect. Genet. Evol. 2013, 19, 258–273. [Google Scholar] [CrossRef]
- Amambua-Ngwa, A.; Tetteh, K.K.A.; Manske, M.; Gomez-Escobar, N.; Stewart, L.B.; Deerhake, M.E.; Cheeseman, I.H.; Newbold, C.I.; Holder, A.A.; Knuepfer, E.; et al. Population Genomic Scan for Candidate Signatures of Balancing Selection to Guide Antigen Characterization in Malaria Parasites. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [Green Version]
- Hull, R.; Dlamini, Z. The role played by alternative splicing in antigenic variability in human endo-parasites. Parasites Vectors 2014, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Yeoh, L.M.; Goodman, C.D.; Mollard, V.; McHugh, E.; Lee, V.V.; Sturm, A.; Cozijnsen, A.; McFadden, G.I.; Ralph, S.A. Alternative splicing is required for stage differentiation in malaria parasites. Genome Biol. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Haase, S.; Cabrera, A.; Langer, C.; Treeck, M.; Struck, N.; Herrmann, S.; Jansen, P.W.; Bruchhaus, I.; Bachmann, A.; Dias, S.; et al. Characterization of a conserved rhoptry-associated leucine zipper-like protein in the malaria parasite Plasmodium falciparum. Infect. Immun. 2008, 76, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Lindner, S.E.; Swearingen, K.E.; Harupa, A.; Vaughan, A.M.; Sinnis, P.; Moritz, R.L.; Kappe, S.H.I. Total and putative surface proteomics of malaria parasite salivary gland sporozoites. Mol. Cell. Proteom. 2013, 12, 1127–1143. [Google Scholar] [CrossRef] [Green Version]
- Doolan, D.L. Plasmodium Immunomics. Int. J. Parasitol 2011, 41, 3–20. [Google Scholar] [CrossRef]
- Doolan, B.D.L.; Sedegah, M.; Hedstrom, R.C.; Charoenvit, P.H.; Hoffrnan, S.L. Circumventing Genetic Restriction of Protection against Malaria with Multigene DNA Immunization: CD8 + T Cell, Interferon, and Nitric Oxide-Dependent Immunity. J. Exp. Med. 1996, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osier, F.H.A.; Fegan, G.; Polley, S.D.; Murungi, L.; Verra, F.; Tetteh, K.K.A.; Lowe, B.; Mwangi, T.; Bull, P.C.; Thomas, A.W.; et al. Breadth and magnitude of antibody responses to multiple Plasmodium falciparum merozoite antigens are associated with protection from clinical malaria. Infect. Immun. 2008, 76, 2240–2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecílio, P.; Pérez-Cabezas, B.; Fernández, L.; Moreno, J.; Carrillo, E.; Requena, J.M.; Fichera, E.; Reed, S.G.; Coler, R.N.; Kamhawi, S.; et al. Pre-clinical antigenicity studies of an innovative multivalent vaccine for human visceral leishmaniasis. PLoS Negl. Trop. Dis. 2017, 11, e0005951. [Google Scholar] [CrossRef] [Green Version]
- Jayaraj, R.; Piedrafita, D.; Dynon, K.; Grams, R.; Spithill, T.W.; Smooker, P.M. Vaccination against fasciolosis by a multivalent vaccine of stage-specific antigens. Vet. Parasitol. 2009, 160, 230–236. [Google Scholar] [CrossRef]
- Kumar, G.; Kocour, M. Applications of next-generation sequencing in fisheries research: A review. Fish. Res. 2017, 186, 11–22. [Google Scholar] [CrossRef]
- Sudhagar, A.; Ertl, R.; Kumar, G.; El-Matbouli, M. Transcriptome profiling of posterior kidney of brown trout, Salmo trutta, during proliferative kidney disease. Parasites Vectors 2019, 12, 1–17. [Google Scholar] [CrossRef]
- Kumar, G.; Ertl, R.; Bartholomew, J.L.; El-Matbouli, M. First transcriptome analysis of bryozoan Fredericella sultana, the primary host of myxozoan parasite Tetracapsuloides bryosalmonae. PeerJ 2020. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Ertl, R.; Bartholomew, J.L.; El-Matbouli, M. Transcriptome analysis elucidates the key responses of bryozoan Fredericella sultana during the development of Tetracapsuloides bryosalmonae (Myxozoa). Int. J. Mol. Sci. 2020, 21, 5910. [Google Scholar] [CrossRef]
- Kumar, G.; Ertl, R.; Nilsen, F.; Bartholomew, J.L.; El-Matbouli, M. Data of de novo transcriptome assembly of the myxozoan parasite Tetracapsuloides bryosalmonae. Data Br. 2021, 35, 106831. [Google Scholar] [CrossRef]
- Ronza, P.; Robledo, D.; Bermúdez, R.; Losada, A.P.; Pardo, B.G.; Sitjà-Bobadilla, A.; Quiroga, M.I.; Martínez, P. RNA-seq analysis of early enteromyxosis in turbot (Scophthalmus maximus): New insights into parasite invasion and immune evasion strategies. Int. J. Parasitol. 2016, 46, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Hartigan, A.; Kosakyan, A.; Pecková, H.; Eszterbauer, E.; Holzer, A.S. Transcriptome of Sphaerospora molnari (Cnidaria, Myxosporea) blood stages provides proteolytic arsenal as potential therapeutic targets against sphaerosporosis in common carp. BMC Genom. 2020, 21, 404. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.S.; Neuhof, M.; Rubinstein, N.D.; Diamant, A.; Philippe, H.; Huchon, D.; Cartwright, P. Genomic insights into the evolutionary origin of Myxozoa within Cnidaria. Proc. Natl. Acad. Sci. USA 2015, 112, 14912–14917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, Z.Q.; Yang, M.; Wang, H.Q.; Xu, Y.; Huang, M.Z.; Lao, G.F.; Li, Y.W.; Li, A.X.; Luo, X.C.; Dan, X.M. Grouper (Epinephelus coioides) BCR signaling pathway was involved in response against Cryptocaryon irritans infection. Fish Shellfish Immunol. 2016, 57, 198–205. [Google Scholar] [CrossRef]
- Kumar, G.; Gotesman, M.; El-Matbouli, M. Interaction of Tetracapsuloides bryosalmonae, the causative agent of proliferative kidney disease, with host proteins in the kidney of Salmo trutta. Parasitol. Res. 2015, 114, 1721–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piriatinskiy, G.; Atkinson, S.D.; Park, S.; Morgenstern, D.; Brekhman, V.; Yossifon, G.; Bartholomew, J.L.; Lotan, T. Functional and proteomic analysis of Ceratonova shasta (Cnidaria: Myxozoa) polar capsules reveals adaptations to parasitism. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Saleh, M.; Kumar, G.; Abdel-Baki, A.A.S.; Dkhil, M.A.; El-Matbouli, M.; Al-Quraishy, S. Quantitative proteomic profiling of immune responses to Ichthyophthirius multifiliis in common carp skin mucus. Fish Shellfish Immunol. 2019, 84, 834–842. [Google Scholar] [CrossRef]
- Mai, Y.Z.; Li, Y.W.; Li, R.J.; Li, W.; Huang, X.Z.; Mo, Z.Q.; Li, A.X. Proteomic analysis of differentially expressed proteins in the marine fish parasitic ciliate Cryptocaryon irritans. Vet. Parasitol. 2015, 211, 1–11. [Google Scholar] [CrossRef]
- Coyne, R.S.; Hannick, L.; Shanmugam, D.; Hostetler, J.B.; Brami, D.; Joardar, V.S.; Johnson, J.; Radune, D.; Singh, I.; Badger, J.H.; et al. Comparative genomics of the pathogenic ciliate Ichthyophthirius multifiliis, its free-living relatives and a host species provide insights into adoption of a parasitic lifestyle and prospects for disease control. Genome Biol. 2011, 12, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Wang, J.L.; Huang, S.Y.; Yang, W.B.; Zhu, W.N.; Zhu, X.Q. Immune responses and protection after DNA vaccination against Toxoplasma gondii calcium-dependent protein kinase 2 (TgCDPK2). Parasite 2017, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Zhou, A.; Lu, G.; Meng, M.; Sun, M.; Bai, Y.; Han, Y.; Wang, L.; Zhou, H.; Cong, H.; et al. Identification and characterization of Toxoplasma gondii aspartic protease 1 as a novel vaccine candidate against toxoplasmosis. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef] [PubMed]
- National Centre for Biotechnology Information. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/bioproject/PRJNA241036/ (accessed on 14 April 2020).
- National Centre for Biotechnology Information. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/sra/SRX4615721/ (accessed on 14 April 2020).
- Hahn, C.; Fromm, B.; Bachmann, L. Comparative genomics of flatworms (Platyhelminthes) reveals shared genomic features of ecto-and endoparastic neodermata. Genome Biol. Evol. 2014, 6, 1105–1117. [Google Scholar] [CrossRef] [Green Version]
- National Centre for Biotechnology Information. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/sra/?term=SRX2417163 (accessed on 14 April 2020).
- National Centre for Biotechnology Information. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/sra/?term=SRX2417144 (accessed on 14 April 2020).
- Ivory, C.; Chadee, K. DNA vaccines: Designing strategies against parasitic infections. Genet. Vaccines Ther. 2004, 2, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budiño, B.; Lamas, J.; Pata, M.P.; Arranz, J.A.; Sanmartín, M.L.; Leiro, J. Intraspecific variability in several isolates of Philasterides dicentrarchi (syn. Miamiensis avidus), a scuticociliate parasite of farmed turbot. Vet. Parasitol. 2011, 175, 260–272. [Google Scholar] [CrossRef]
Figure 1.
Conceptual representation of multivalent/multiepitope vaccine formulation for fish parasites in the omics era. Analysis of genomic, transcriptomic and proteomic data of parasite and the data obtained from host–parasite interactions enable identifying a suitable antigen as vaccine candidates. Genome sequence contains the entire genetic repertoire of antigens from which novel vaccine targets can be selected. Transcriptome analysis provides insights on the parasite gene expression profile leading to successful establishment and pathology in host. Proteomic analysis provides information on protein expression under specified conditions. It is useful in identification of proteins that are expressed by parasite during infection and the subset of proteins which are present on parasite surface (surface proteome). Surface exposed proteins which are immunogenic in the host can be suitable vaccine candidate. Structural genomics helps to know the three-dimensional structure of proteins produced by an organism and how they interact with antibodies or drugs. Immunoproteomics provides information on the proteins or epitopes which interact with host antibodies.
Figure 1.
Conceptual representation of multivalent/multiepitope vaccine formulation for fish parasites in the omics era. Analysis of genomic, transcriptomic and proteomic data of parasite and the data obtained from host–parasite interactions enable identifying a suitable antigen as vaccine candidates. Genome sequence contains the entire genetic repertoire of antigens from which novel vaccine targets can be selected. Transcriptome analysis provides insights on the parasite gene expression profile leading to successful establishment and pathology in host. Proteomic analysis provides information on protein expression under specified conditions. It is useful in identification of proteins that are expressed by parasite during infection and the subset of proteins which are present on parasite surface (surface proteome). Surface exposed proteins which are immunogenic in the host can be suitable vaccine candidate. Structural genomics helps to know the three-dimensional structure of proteins produced by an organism and how they interact with antibodies or drugs. Immunoproteomics provides information on the proteins or epitopes which interact with host antibodies.

Figure 2.
Workflow for identification of vaccine candidates and vaccine formulation using high throughput omics techniques. The material of interest (DNA, RNA or protein) can be extracted from the desired parasite or from-infected host fish tissue. The isolated material serves as sample for next generation sequence analysis (for DNA and RNA) and spectrophotometry and Edman sequencing (for protein). The obtained sequences can be subjected to different bioinformatic tools for analysis and functional annotation of parasite molecules. Based on molecular function and biological pathway analysis, the immunogenic targets (capable of eliciting host immune response) can be selected and their protective epitopes predicted. These molecules can then be produced on a large scale and combined with suitable adjuvants to form vaccine. Subsequently, the vaccine thus produced can be used for conducting trials in suitable fish and its efficacy and appropriate route of administration can be determined.
Figure 2.
Workflow for identification of vaccine candidates and vaccine formulation using high throughput omics techniques. The material of interest (DNA, RNA or protein) can be extracted from the desired parasite or from-infected host fish tissue. The isolated material serves as sample for next generation sequence analysis (for DNA and RNA) and spectrophotometry and Edman sequencing (for protein). The obtained sequences can be subjected to different bioinformatic tools for analysis and functional annotation of parasite molecules. Based on molecular function and biological pathway analysis, the immunogenic targets (capable of eliciting host immune response) can be selected and their protective epitopes predicted. These molecules can then be produced on a large scale and combined with suitable adjuvants to form vaccine. Subsequently, the vaccine thus produced can be used for conducting trials in suitable fish and its efficacy and appropriate route of administration can be determined.


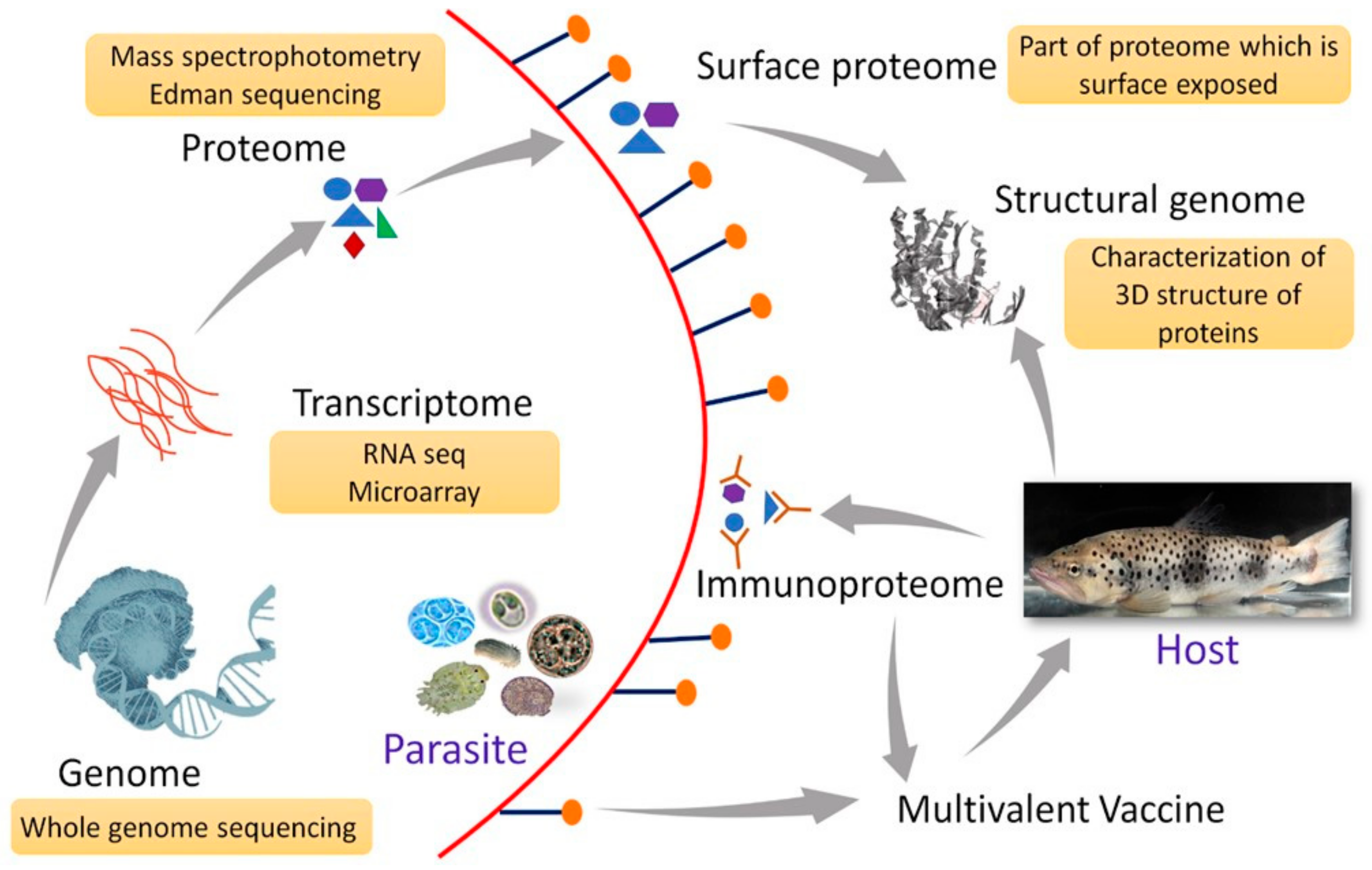{kind=link}
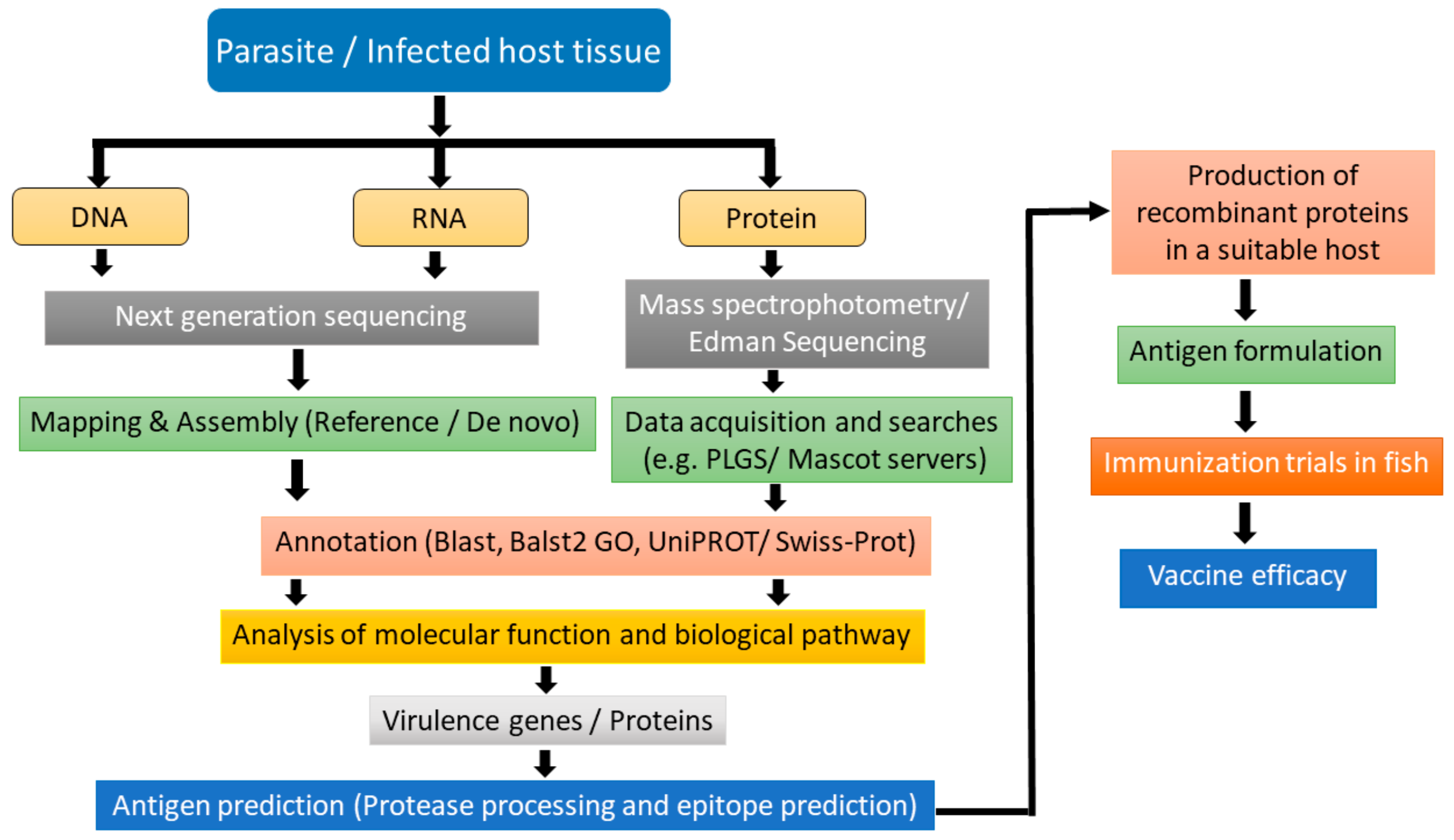{kind=link}
Table 1.
Some economically important fish parasites: Table provides information on mortality and economic loss caused by important parasites of fish. NA: not available, a: mortality, b: economic loss.
Table 1.
Some economically important fish parasites: Table provides information on mortality and economic loss caused by important parasites of fish. NA: not available, a: mortality, b: economic loss.
| Parasite Group | Parasite | Disease | Host | Mortality a/ Economic Loss b | Reference |
|---|---|---|---|---|---|
| Freshwater Endoparasites | |||||
| Cnidaria | T. bryosalmonae | Proliferative kidney disease | Rainbow trout | 95% a | [26] |
| M. cerebralis | Whirling disease | Rainbow trout | 90% a | [27] | |
| S. molnari | Sphaerosporosis | Carps | NA | [28] | |
| Platyhelminthes | Bothriocephalus acheilo gnathi | Ulcer, catarrhal enteritis | Carps | 100% a | [29] |
| Freshwater Ectoparasites | |||||
| Ciliophora | I. multifiliis | White spot | Rainbow trout, Carps | NA | [30] |
| Arthropoda | Argulus | Argulosis | Indian Major Carps | 5.41 million USD b | [31] |
| Platyhelminthes | Gyrodactylus salaris | Gyrodactylosis | Atlantic salmon | NA | [32] |
| Marine Endoparasites | |||||
| Cnidaria | K. thyrsites | Soft flesh syndrome | Salmonids | 6 million CAD b | [33] |
| C. shashta | Ceratomyxosis | Salmonids | NA | [34] | |
| E. leei | Enteromyxosis | Sharpsnout seabream | NA | [35] | |
| Ciliophora | Uronema marinum, Philasterides dicentrarchi | Scuticociliatosis | Seabass, Turbot | NA | [36] |
| Marine Ectoparasites | |||||
| Ciliophora | C. irritans | White spot | Greater Amberjack | 834 USD b | [3] |
| Arthropoda | Caligus rogercresseyi, Lepeophtheirus salmonis | Sea louse disease | Salmonids | 100 million USD b | [37] |
Table 2.
Overview of immunization trials in fish for ectoparasites: Table provides information on immunization trials for important ectoparasites of fish. IM: Intramuscular, IP: Intraperitoneal.
Table 2.
Overview of immunization trials in fish for ectoparasites: Table provides information on immunization trials for important ectoparasites of fish. IM: Intramuscular, IP: Intraperitoneal.
| Disease | Parasite | Fish Host | Vaccine Trial Type | Antigen Target | Delivery Method | Reference |
|---|---|---|---|---|---|---|
| White spot disease (Ich) | Ichthyophthirius multifiliis | Channel catfish | DNA | (i-antigen) immobilization antigen | IM | [99] |
| Channel catfish | Live | Theront | IP | [100] | ||
| Rainbow trout | Subunit | Recombinant proteins (#5, #10, & #11) | IP | [101] | ||
| DNA | (i-antigen) immobilization antigen & Cysteine protease | IM, Needle free injection, Gene gun delivery | [102] | |||
| Live | Theronts | IP | [103] | |||
| Killed | Sonicated formalin killed trophonts | IP | [104] | |||
| Nile Tilapia | Live | Theront and sonicated trophonts | Immersion and IP | [105] | ||
| Channel catfish | Live | Tomites | IP | [98] | ||
| Killed | Trophonts and tomites | IP | ||||
| Marine White spot | Cryptocaryon irritans | Grouper | Live | Theronts | Bath | [106] |
| Killed | Formalin-killed theronts | IP | [107] | |||
| Sea louse infestations | Lepeophtheirus salmonis | Atlantic salmon | Crude parasite extract | Adult female parasite | IP |
Table 3.
Overview of immunization trials in fish against parasites: Table provides information on immunization trials for important endoparasites. NA: Not Available, IM: Intramuscular, IP: Intraperitoneal.
Table 3.
Overview of immunization trials in fish against parasites: Table provides information on immunization trials for important endoparasites. NA: Not Available, IM: Intramuscular, IP: Intraperitoneal.
| Disease | Parasite | Fish Host | Vaccine Type | Antigen Target | Delivery Method | Reference |
|---|---|---|---|---|---|---|
| Freshwater Endoparasites | ||||||
| PKD | Tetracapsuloides bryosalmonae | Rainbow trout | DNA | Micro-exon gene (TB-MEG1) | Not Available | [108] |
| Myxobolosis | Myxobolus koi | Gold fish | Subunit | Crude protein spore | Immersion | [109] |
| Marine Endoparasites | ||||||
| Scuticociliatosis | Uronema marinum | Grouper | Subunit | (i-antigen) immobilization antigen | IP | [110] |
| Philasterides dicentrarchi | Turbot | Subunit | Membrane proteins | IP | [111] | |
Table 4.
Available sequence data for parasites: Table enlists the fish parasites for which genome and transcriptome data are available. a: Transcriptome size, b: Genome size, c: Number of contigs, NA: Not Available.
Table 4.
Available sequence data for parasites: Table enlists the fish parasites for which genome and transcriptome data are available. a: Transcriptome size, b: Genome size, c: Number of contigs, NA: Not Available.
| Parasite | Molecular Data Type | Sequencing Platform | Size/No. of Contigs | Accession No. | Reference |
|---|---|---|---|---|---|
| Endoparasite | |||||
| Tetracapsuloides bryosalmonae | Transcriptome | NextSeq 550 | 25908 c | [155] | |
| Myxobolus cerebralis | Transcriptome | HiSeq 2000 | 52972 c | GBGI00000000 | [158] |
| Kudoa iwatai | Transcriptome | HiSeq 2000 | 1637 c | GBKL00000000 | [158] |
| Ceratomyxa shasta | Transcriptome | Illumina HiSeq 3000 | NA | SRX3741971 | [168] |
| Sphaerospora molnari | Transcriptome | Illumina HiSeq | 29560 c | PRJNA522909 | [157] |
| Myxobolus squamalis | Transcriptome | Illumina HiSeq 3000 | NA | SRX4615721 | [169] |
| Ectoparasite | |||||
| Gyrodactylus salaris | Whole genome | Roche 454 FLX Titanium Illumina GAII | 120 Mb b | JJOG00000000 | [170] |
| Cryptocaryon irritans | Transcriptome (Trophont) | Illumina HiSeq 2000 | 2.6 Gb a | SRX2417163 | [171] |
| Transcriptome (Theront) | Illumina HiSeq 2000 | 3.2 Gb a | SRX2417144 | [172] | |
| Argulus siamensis | Transcriptome | Illumina HiSeq 2000 | 46352 c | SRX150806 | [31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shivam, S.; El-Matbouli, M.; Kumar, G. Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities. Vaccines 2021, 9, 179. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020179
AMA Style
Shivam S, El-Matbouli M, Kumar G. Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities. Vaccines. 2021; 9(2):179. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020179
Chicago/Turabian StyleShivam, Saloni, Mansour El-Matbouli, and Gokhlesh Kumar. 2021. "Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities" Vaccines 9, no. 2: 179. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020179
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.