
Therapeutic Potential of HLA-I Polyreactive mAbs Mimicking the HLA-I Polyreactivity and Immunoregulatory Functions of IVIg


Abstract
:1. Introduction
- Determine if HLA-E shares antigenic amino acid sequences (epitopes) common to all other HLA-I isoforms;
- Document HLA-I polyreactivity of HLA-E mAbs;
- Document HLA-I polyreactivity of the therapeutic preparations of IVIg;
- Compare immunomodulation by IVIg with polyreactive mAbs
- ○
- Suppression of T-cell proliferation;
- ○
- Suppression of antibody production by B cells;
- ○
- Expansion of Foxp3+ Tregs.
2. HLA-E Shares Antigenic Amino Acid Sequences (Epitopes) Common to All Other HLA-I Isoforms
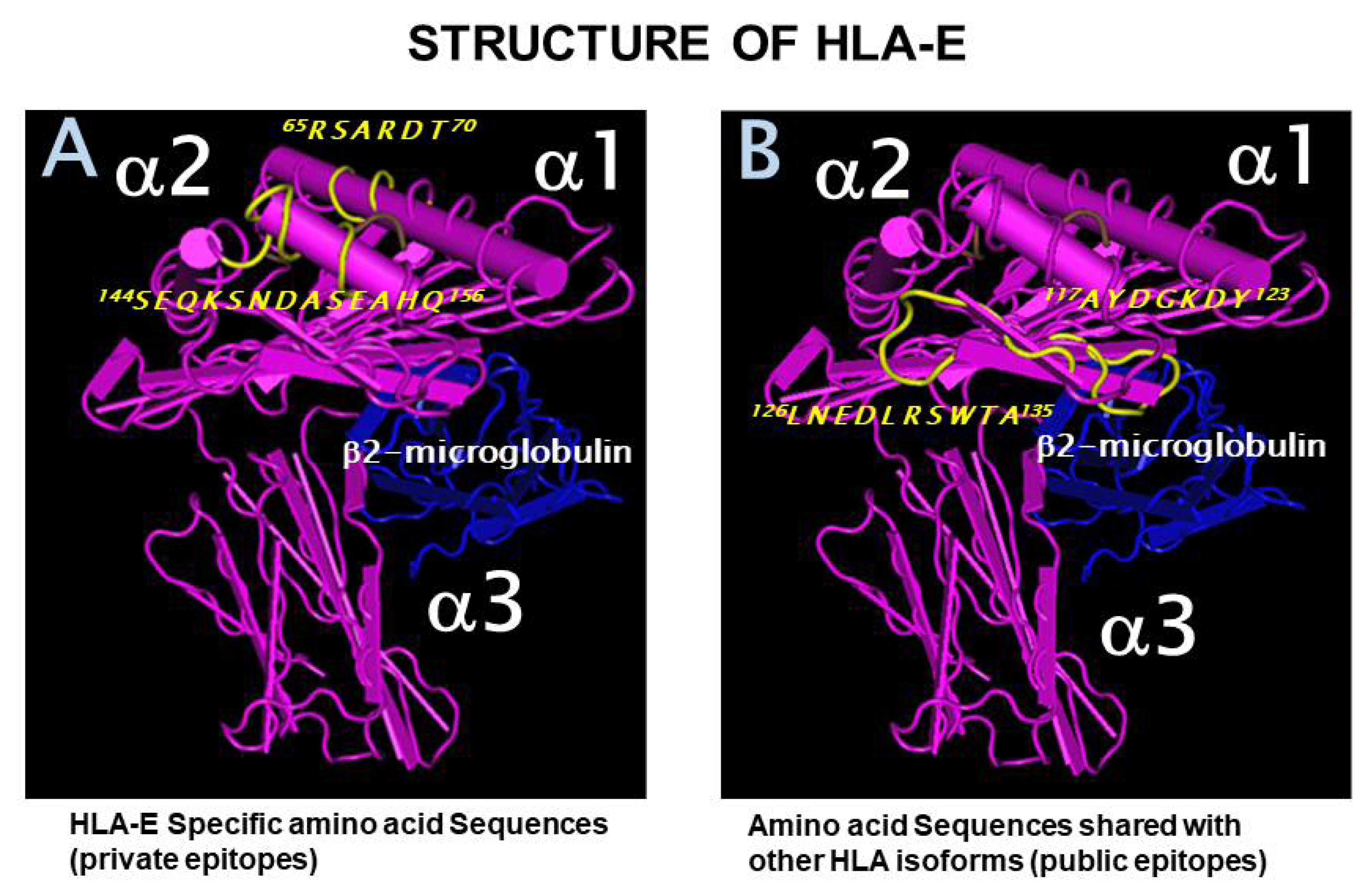
2.1. Private and Public Epitopes of HLA-E
2.2. The Most Widely Shared Public Epitopes Are Cryptic in HLA-I
2.3. Antigenicity Rank of the Most Widely Shared Public Epitopes of HLA-E
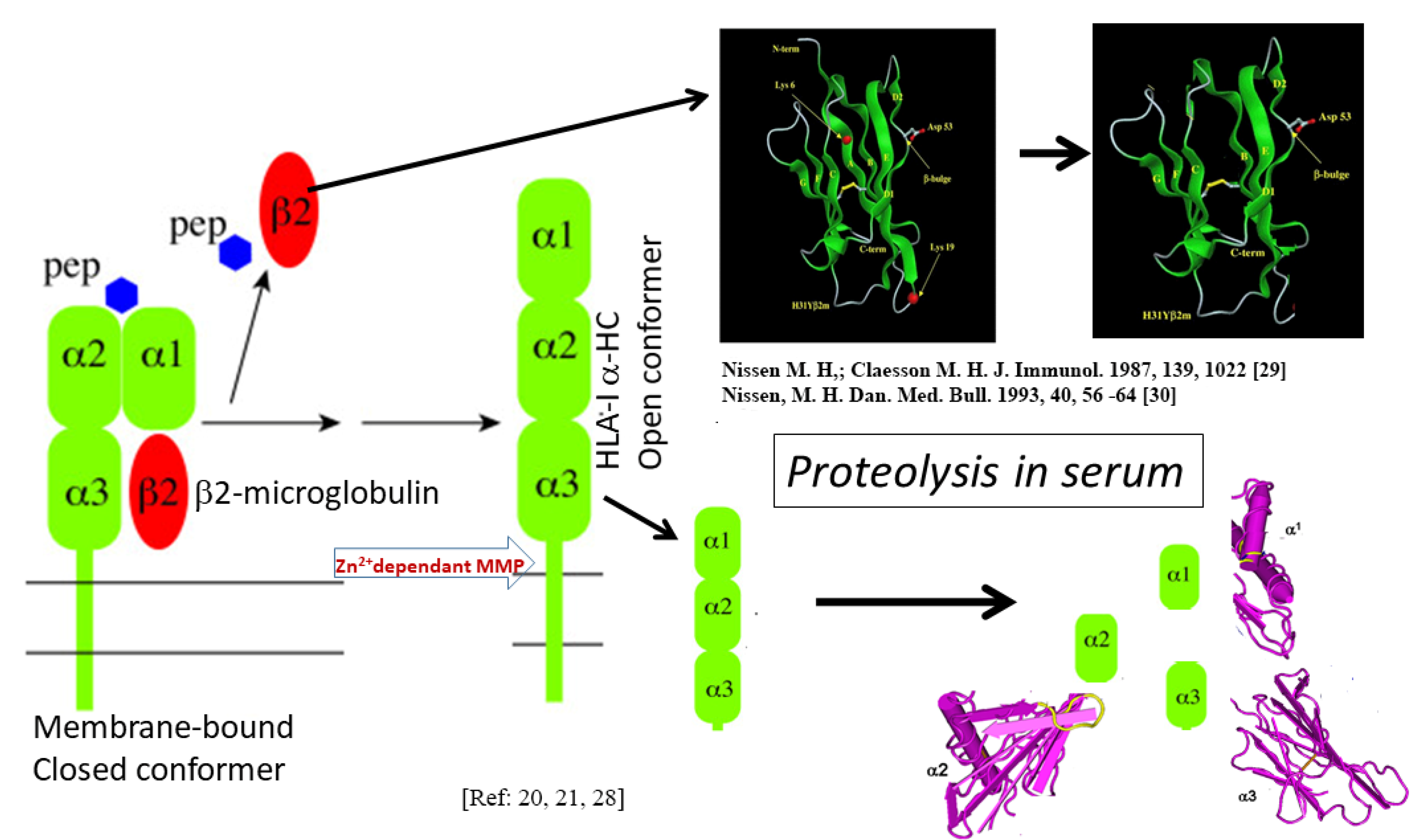
3. Documentation of HLA-I Polyreactivity of HLA-E mAbs
3.1. Methodology–Development and Characterization of HLA-E mAbs
3.1.1. Anti-HLA-E mAb Production
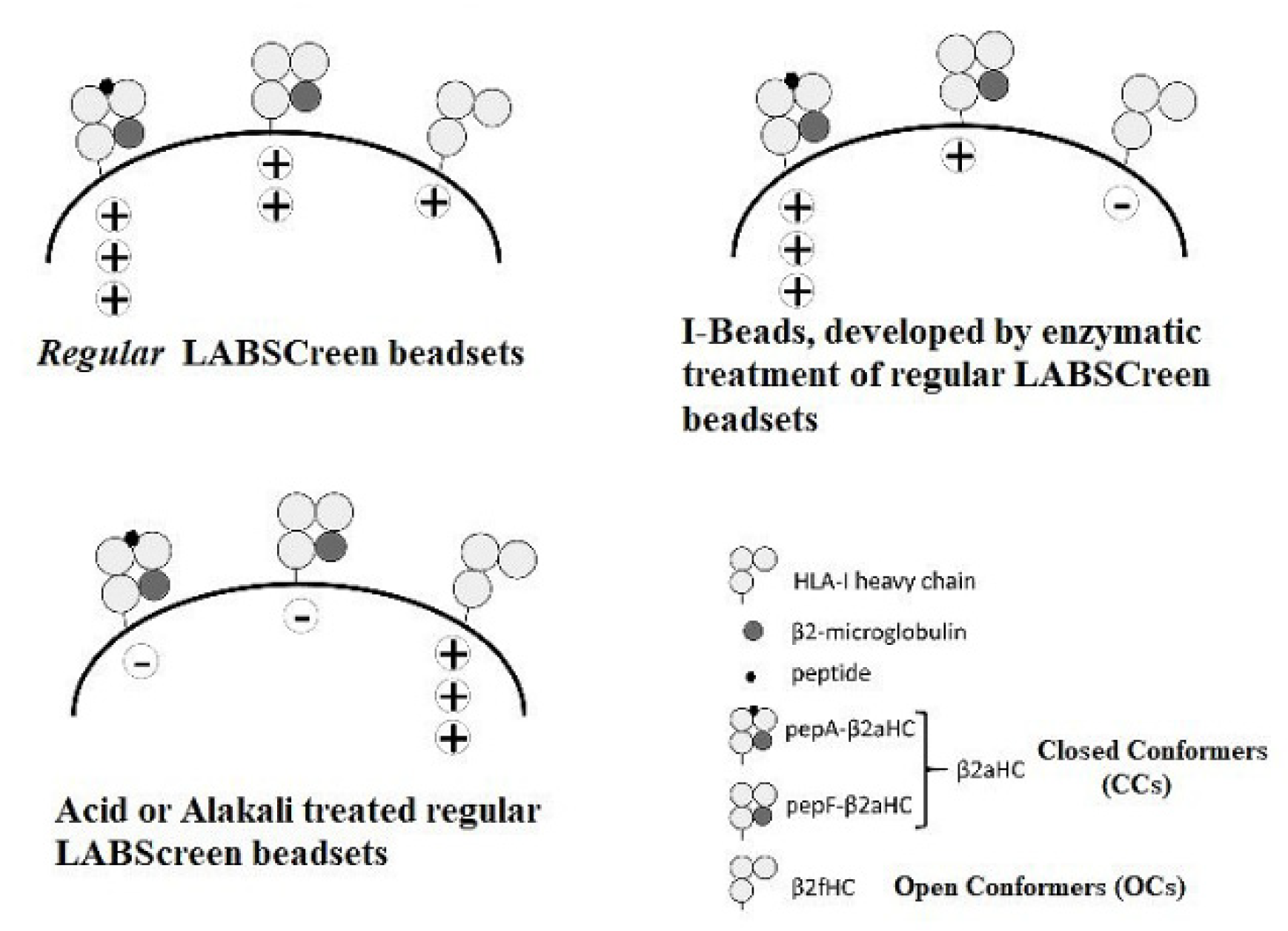
3.1.2. HLA-1 Antigen-Coated Single Antigen Beads (SABs) and the Luminex Platform for Monitoring the Reactivity and the Density of These mAbs
3.2. The HLA Reactivity Groups of mAbs Generated by Recombinant HLA-E OCs
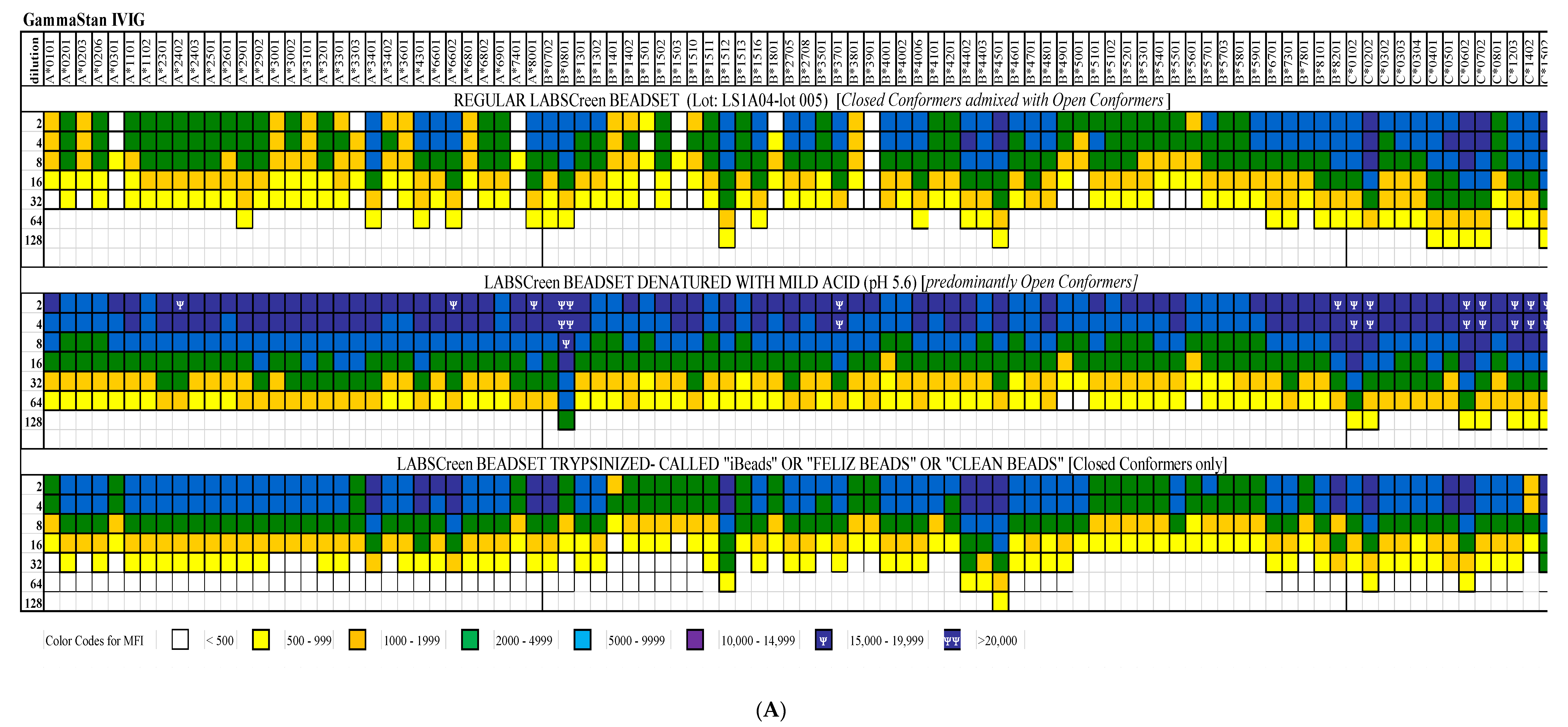
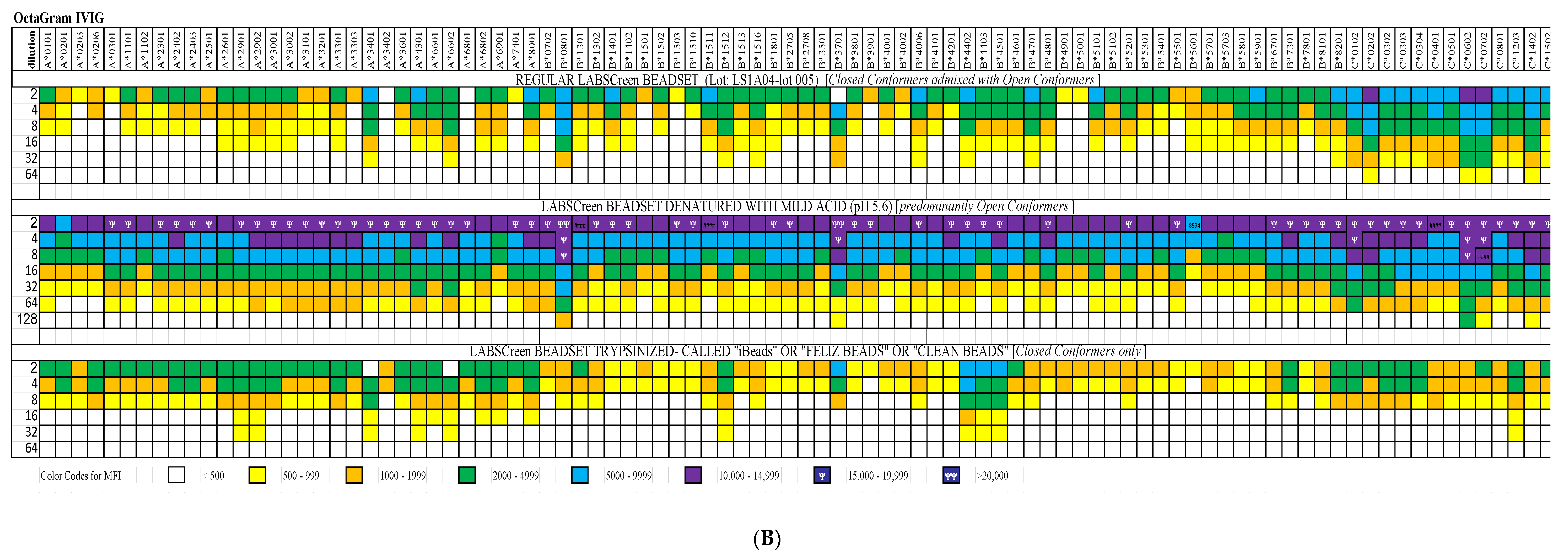
4. Documentation of HLA-I Reactivity of the Therapeutic Preparations of IVIg
5. Immunomodulation By IVIg Compared with Polyreactive Monoclonal Antibodies
5.1. Suppression of T-Cell Proliferation: IVIg vs. HLA-I-Polyreactive mAbs
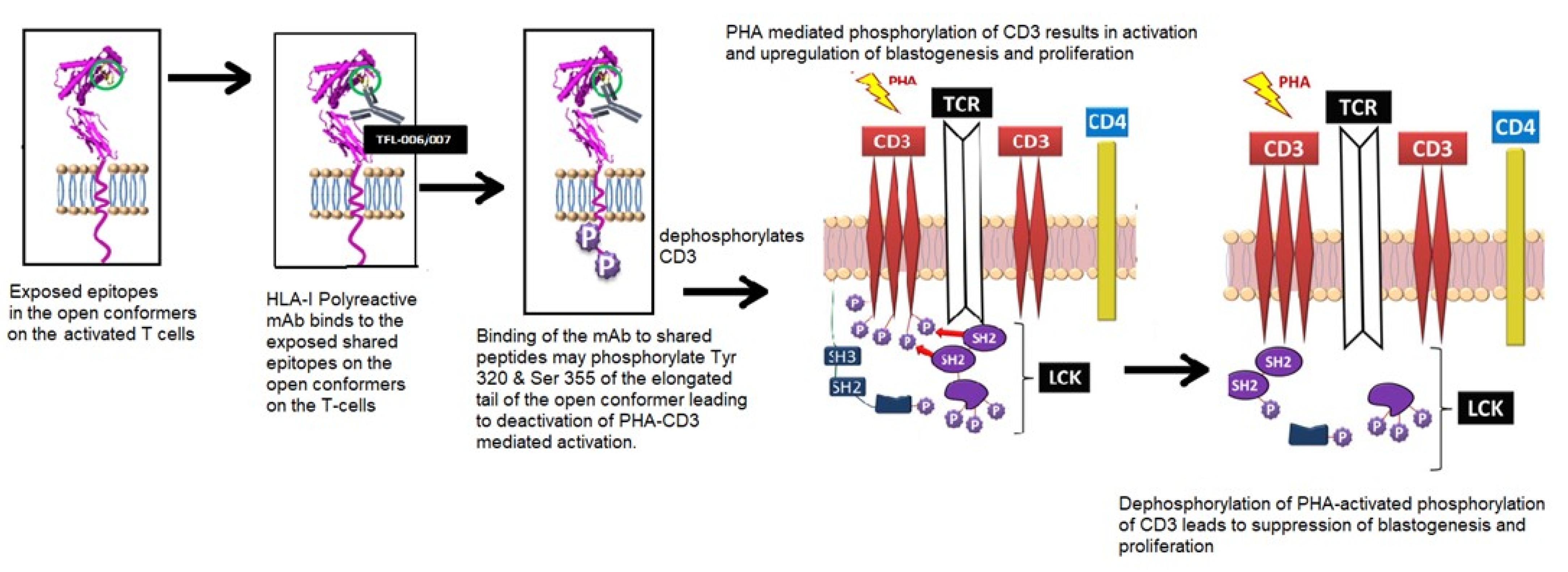
5.1.1. Background and Hypothesis
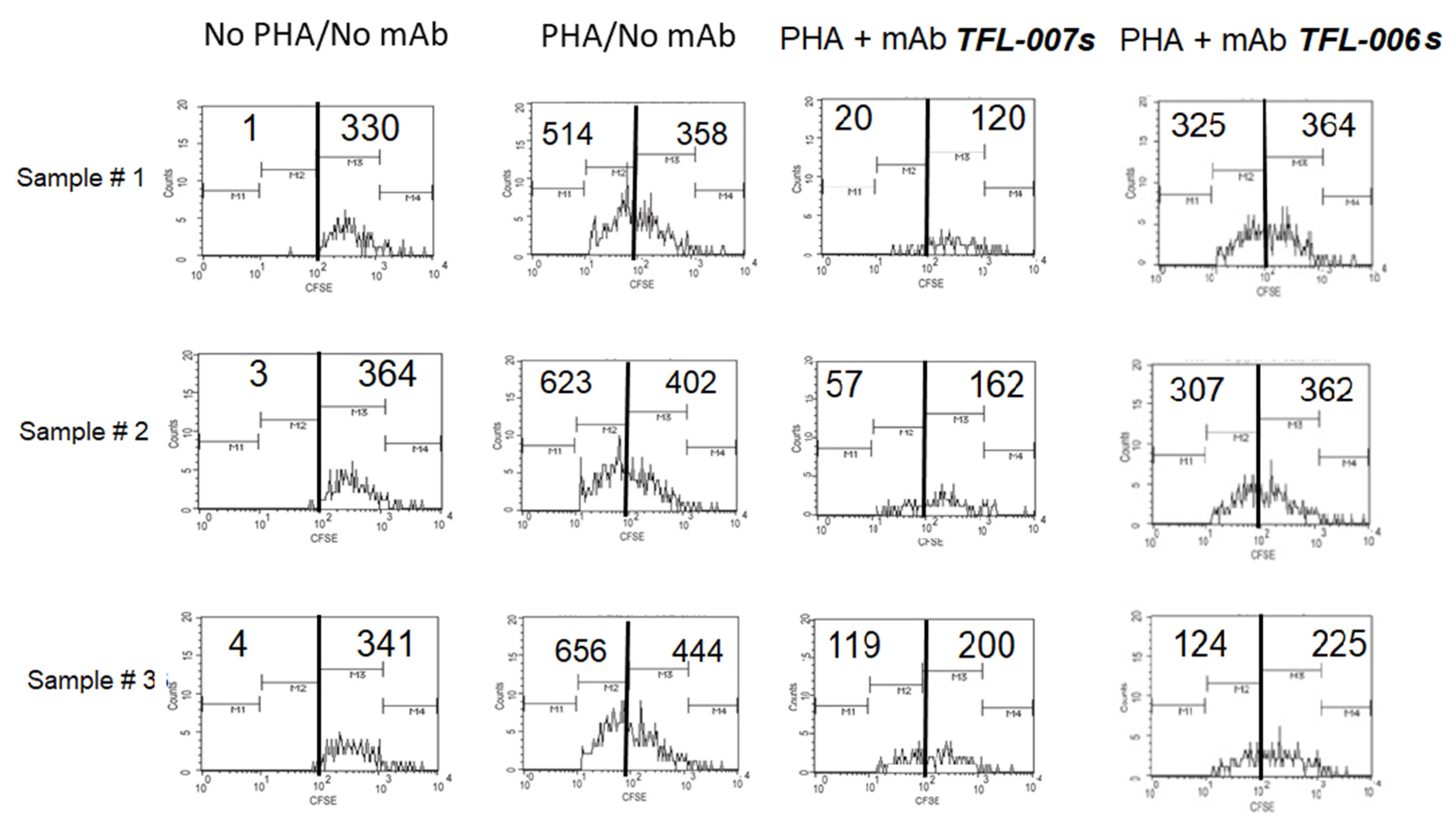
5.1.2. Hypothesis Testing: Measurement of T-Lymphocyte Proliferation
5.1.3. The Suppression of Activated T-Cells by IVIg vs. mAbs
5.2. Suppression of Antibody Production by B-Cells: IVIg vs. HLA-I Polyreactive mAbs
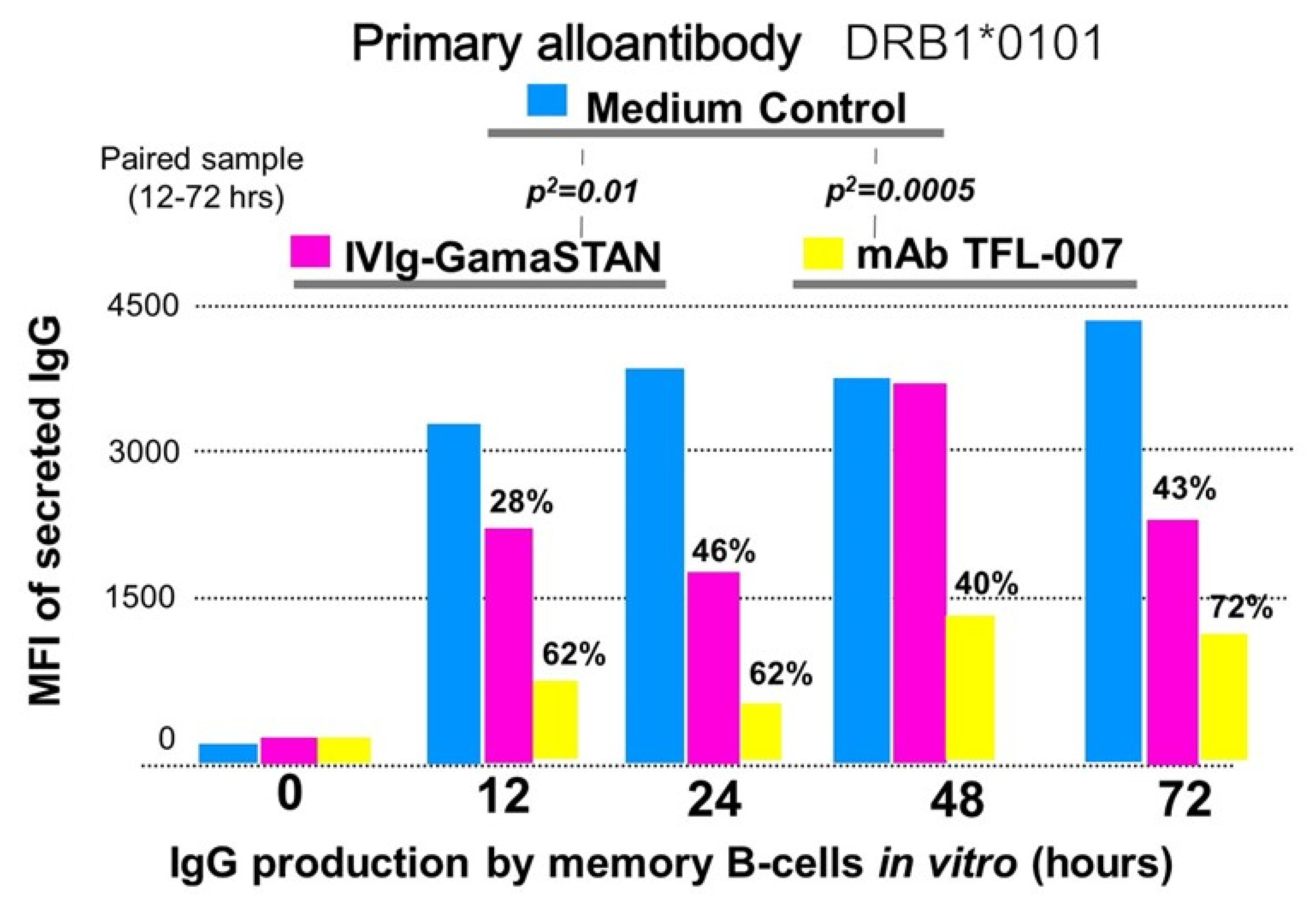
5.2.1. Background and Hypothesis
5.2.2. Methodology to Test the Hypothesis
5.2.3. TFL-007 Suppressed HLA-Antibody Production Better Than That of IVIg
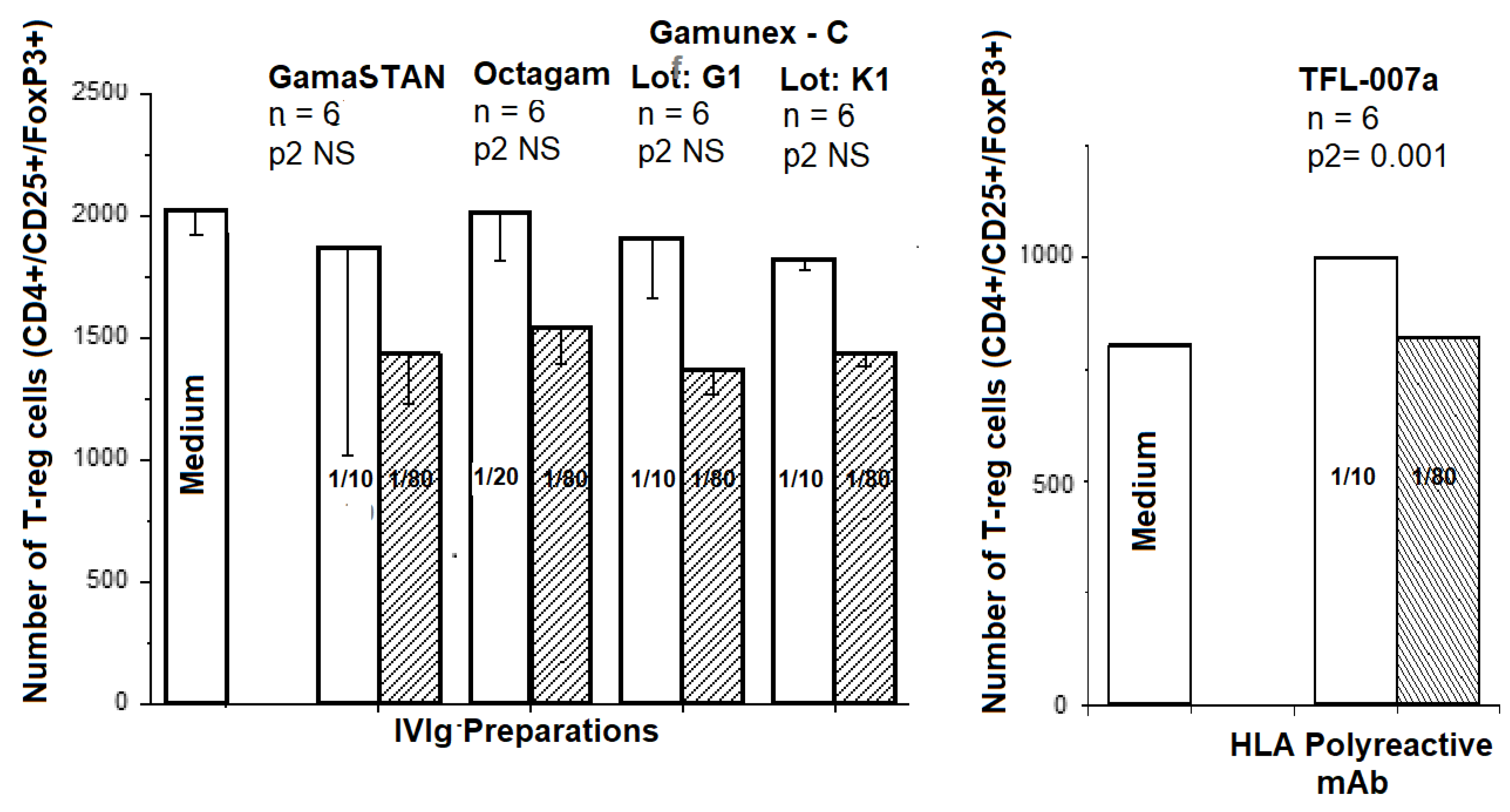
5.3. Expansion of Foxp3+ Tregs In Vitro: IVIg versus HLA-I Polyreactive mAbs
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darabi, K.; OAbdel-Wahab, O.; Dzik, W.H. Current usage of intravenous immune globulin and the rationale behind it: The Massachusetts General Hospital data and a review of the literature. Transfusion 2006, 46, 741–753. [Google Scholar] [CrossRef]
- Glotz, D.; Haymann, J.-P.; Sansonetti, N.; Francois, A.; Menoyo-Calonge, V.; Bariety, J.; Druet, P. Suppression of HLA-specific alloAbs by high-dose intravenous immunoglobulins (IVIg). Transplantation 1993, 56, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Tyan, D.B.; Li, V.A.; Czer, L.; Trento, A.; Jordan, S.C. Intravenous immunoglobulin suppression of HLA alloantibody in highly sensitized transplant candidates and transplantation with a histoincompatible organ. Transplantation 1994, 57, 553–562. [Google Scholar] [CrossRef]
- Sapir, T.; Shoenfeld, Y. Facing the enigma of immunomodulatory effects of intravenous immunoglobulin. Clin. Rev. Aller. Immunol. 2005, 29, 185–199. [Google Scholar] [CrossRef]
- Tongio, M.M.; Falkenrodt, A.; Mitsuishi, Y.; Urlacher, A.; Bergerat, J.P.; North, M.L.; Mayer, S. Natural HLA Abs. Tissue Antigens. 1985, 26, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Luscher, M.A.; Choy, G.; Embree, J.E.; Nagelkerke, N.J.; Bwayo, J.J.; Nienga, S.; Plummer, F.A.; Barber, B.H.; MacDonald, K.S. Anti-HLA alloAb is found in children but does not correlate with a lack of HIV type 1 transmission from infected mothers. AIDS Res. Hum. Retrovir. 1998, 14, 99–107. [Google Scholar] [CrossRef]
- Morales-Buenrostro, L.E.; Terasaki, P.I.; Marino-Vazquez, L.A.; Lee, J.-H.; El-Awar, N.; Alberú, J. “Natural” human leukocyte antigen Abs found in nonalloimmunized healthy males. Transplantation 2008, 86, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Saito, S.; Nakazawa, Y.; Kobayashi, N.; Matsuda, M.; Matsumoto, Y.; Hosoyama, T.; Koike, K. Existence of an immunoglobulin G component of naturally occurring HLA class I Abs that are not directed against self-antigens in human serum. Tissue Antigens. 2008, 72, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, M.H.; Kaneku, H.; El-Awar, N.; Morales-Buenrostro, L.E.; Terasaki, P.I. Abs to HLA-E in alloimmunized males: Pattern of HLA-Ia reactivity of anti-HLA-E-positive sera. J. Immunol. 2010, 185, 1935–1948. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, M.H.; Taniguchi, M.; Chen, C.W.; Ozawa, M.; Kaneku, H.; El-Awar, N.; Cai, J.; Terasaki, P.I. HLA-E mAbs recognize shared peptide sequences on classical HLA class Ia:relevance to human natural HLA Abs. Mol. Immunol. 2010, 47, 1121–1131. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Pham, T.; El-Awar, N.; Kaneku, H.; Terasaki, P.I. Anti-HLA-E mAb 3D12 mimics MEM-E/02 in binding to HLA-B and HLA-C alleles: Web-tools validate the immunogenic epitopes of HLA-E recognized by the Abs. Mol. Immunol. 2011, 48, 423–430. [Google Scholar] [CrossRef]
- Marsh, S.G.E.; Parham, P.; Barber, L.D. The HLA Facts Book; Academic Press: San Diego, CA, USA, 2000; 400p. [Google Scholar]
- Schnabl, E.; Stockinger, H.; Majdic, O.; Gaugitsch, H.; Lindley, I.J.; Maurer, D.; Hajek-Rosenmayr, A.; Knapp, W. Activated human T lymphocytes express MHC class I heavy chains not associated with beta 2 m. J. Exp. Med. 1990, 171, 1431–1442. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Heemels, M.T.; Neefjes, J.J.; Kast, W.M.; Melief, C.J.; Ploegh, H.L. Direct binding of peptide to empty MHC class I molecules on intacT-cells and in vitro. Cell 1990, 62, 563–567. [Google Scholar] [CrossRef]
- Benjamin, R.J.; Madrigal, J.A.; Parham, P. Peptide binding to empty HLA-B27 molecules of viable human cells. Nature 1991, 351, 74–77. [Google Scholar] [CrossRef]
- Majdic, O.; Schnabl, E.; Stockinger, H.; Gadd, S.; Maurer, D.; Radaszkiewics, T. LA45, an activation-induced human lymphocyte antigen with strong homology to MHC class I molecules. In Leukocyte Typing IV; Oxford University Press: Oxford, UK, 1989; Volume 511, 8p. [Google Scholar]
- Madrigal, J.A.; Belich, M.P.; Benjamin, R.J.; Little, A.M.; Hildebrand, W.H.; Mann, D.L.; Parham, P. Molecular definition of a polymorphic antigen (LA45) of free HLA-A and -B heavy chains found on the surfaces of activated B and T-cells. J. Exp. Med. 1991, 174, 1085–1095. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.G.; Simon, J.; Powis, S.J.; Arosa, F.A. Misfolding of Major Histocompatibility Complex Class I Molecules in Activated T-cells Allows cis-Interactions with Receptors and Signaling Molecules and Is Associated with Tyrosine Phosphorylation. J. Biol. Chem. 2004, 279, 53062–53070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arosa, F.A.; Santos, S.G.; Powis, S.J. OC: The hidden face of MHC-I molecules. Trends Immunol. 2007, 28, 115–123. [Google Scholar] [CrossRef]
- Demaria, S.; Schwab, R.; Bushkin, Y. The origin and fate of beta 2m-free MHC class I molecules induced on activated T-cells. Cell Immunol. 1992, 142, 103–113. [Google Scholar] [CrossRef]
- Demaria, S.; Bushkin, Y. CD8 and beta-2 m-free MHC class I molecules in T cell immunoregulation. Int. J. Clin. Lab. Res. 1993, 23, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Raine, T.; Brown, D.; Bowness, P.; Hill Gaston, J.S.; Moffett, A.; Trowsdale, J.; Allen, R.L. Consistent patterns of expression of HLA class I free heavy chains in healthy individuals and raiAllensed expression in spondyloarthropathy patients point to physiological and pathological roles. Rheumatology 2006, 45, 1338–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, R.L.; O’Callaghan, C.A.; McMichael, A.J.; Bowness, P. Cutting edge: HLA-B27 can form a novel beta2m-free H chain homodimer structure. J. Immunol. 1999, 162, 5045–5048. [Google Scholar] [PubMed]
- Khare, S.D.; Hansen, J.; Luthra, H.S.; David, C.S. HLA-B27 heavy chains contribute to spontaneous inflammatory disease in B27/human beta2m (beta2m) double transgenic mice with disrupted mouse beta2m. J. Clin. Investig. 1996, 98, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Capps, G.G.; Robinson, B.E.; Lewis, K.D.; Zúñiga, M.C. In vivo dimeric association of class I MHC heavy chains. Possible relationship to class I MHC heavy chain-beta 2m dissociation. J. Immunol. 1993, 151, 159–169. [Google Scholar] [PubMed]
- Matko, J.; Bushkin, Y.; Wei, T.; Edidin, M. Clustering of class I HLA molecules on the surfaces of activated and transformed human cells. J. Immunol. 1994, 152, 3353–3360. [Google Scholar]
- Allen, R.L.; Raine, T.; Haude, A.; Trowsdale, J.; Wilson, M.J. Leukocyte receptor complex-encoded immunomodulatory receptors show differing specificity for alternative HLA-B27 structures. J. Immunol. 2001, 167, 5543–5547. [Google Scholar] [CrossRef] [Green Version]
- Demaria, S.; Schwab, R.; Gottesman, S.R.; Bushkin, Y. Soluble beta 2-m-free class I heavy chains are released from the surface of activated and leukemia cells by a metalloprotease. J. Biol. Chem. 1994, 269, 6689–6694. [Google Scholar] [CrossRef]
- Nissen, M.H.; Claësson, M.H. Proteolytically modified human beta 2-microglobulin augments the specific cytotoxic activity in murine mixed lymphocyte culture. J. Immunol. 1987, 139, 1022–1029. [Google Scholar]
- Nissen, M.H. Proteolytic modification of beta 2-microglobulin in human serum. Dan. Med. Bull. 1993, 40, 56–64. [Google Scholar]
- Chou, P.Y.; Fasman, G.D. Prediction of the secondary structure of proteins from their amino acid sequence. Adv. Enzymol. Relat. Areas Mol. Biol. 1978, 47, 45–148. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef] [Green Version]
- Karplus, P.A.; Schulz, G.E. Prediction of chain flexibility in proteins—A tool for the selection of peptide antigens. Naturwissenschaft 1985, 72, 212–213. [Google Scholar] [CrossRef]
- Parker, J.M.; Guo, D.; Hodges, R.S. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: Correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry 1986, 25, 5425–5432. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V.; Kawakita, S. Therapeutic preparations of IVIg contain naturally occurring anti-HLA-E Abs that react with HLA-Ia (HLA-A/-B/-Cw) alleles. Blood 2013, 121, 2013–2028. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, M.H.; Zhu, D.; Pham, T.; Jucaud, V.; Hopfield, J.; Kawakita, S.; Terasaki, P.I. Anti-HLA-E mAbs reacting with HLA-la and lb alleles like IVIg as potential IVIg-immunomimetics: An evolving therapeutic concept. Clin. Transpl. 2013, 293–305. [Google Scholar]
- Ward, P.A.; Adams, J.; Faustman, D. Monoclonal Antibody Production A Report of the; Committee on Methods of Producing MAbs Institute for Laboratory Animal Research National Research Council National Academy Press: Washington, DC, USA, 1999; pp. 1–47. [Google Scholar]
- Sasaki, T.; Ravindranath, M.H.; Terasaki, P.I.; Freitas, M.C.; Kawakita, S.; Jucaud, V. Gastric cancer progression may involve a shift in HLA-E profile from an intact heterodimer to β2m-free monomer. Int. J. Cancer 2014, 134, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Parham, P.; Barnstable, C.J.; Bodmer, W.F. Use of a monoclonal antibody (W6/32) in structural studies of HLA-A,B,C, antigens. J. Immunol. 1979, 123, 342–349. [Google Scholar] [PubMed]
- Smith, K.D.; Mace, B.E.; Valenzuela, A.; Vigna, J.L.; McCuthcheon, J.A.; Barbosa, J.A.; Huczko, E.; Engelhard, V.H.; Lutz, C.T. Probing HLA-B7 conformational shifts induced by peptide-binding groove mutations and bound peptide with anti-HLA mAbs. J. Immunol. 1996, 157, 2470–2478. [Google Scholar] [PubMed]
- Vigna, J.L.; Smith, K.D.; Lutz, C.T. Invariant chain association with MHC class I: Preference for HLA class I/beta 2-m heterodimers, specificity, and influence of the MHC peptide-binding groove. J. Immunol. 1996, 157, 4503–4510. [Google Scholar]
- Perosa, F.; Luccarelli, G.; Prete, M.; Favoino, E.; Ferrone, S.; Dammacco, F. Beta 2-m-free HLA class I heavy chain epitope mimicry by monoclonal antibody HC-10-specific peptide. J. Immunol. 2003, 171, 1918–1926. [Google Scholar] [CrossRef] [Green Version]
- Carreno, B.M.; Hansen, T.H. Exogenous peptide ligand influences the expression and half-life of free HLA class I heavy chains ubiquitously detected at the cell surface. Eur. J. Immunol. 1994, 24, 1285–1292. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; El Hilali, F. Monospecific and Polyreactive MAbs against Human Leukocyte Antigen-E: Diagnostic and Therapeutic Relevance. In Monoclonal Antibodies; Resaei, N., Ed.; Intech Open: London, UK, 2021; 38p, ISBN 978-1-83968-370-1. [Google Scholar]
- Jucaud, V.; Ravindranath, M.H.; Terasaki, P.I. Conformational Variants of the Individual HLA-I Antigens on Luminex Single Antigen Beads Used in Monitoring HLA Abs: Problems and Solutions. Transplantation 2017, 101, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Jucaud, V. Conformational variants of HLA-I antigens on Luminex Single Antigen Beads for monitoring Abs. Transplantation 2017, 101, e153–e154. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Jucaud, V.; Ferrone, S. Monitoring native HLA-I trimer specific Abs in Luminex multiplex single antigen bead assay: Evaluation of beadsets from different manufacturers. J. Immunol. Methods 2017, 450, 73–80, Erratum in 2018, 460, 125. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H. HLA Class Ia and Ib Polyreactive Anti-HLA-E IgG2a MAbs (TFL-006 and TFL-007) Suppress Anti-HLA IgG Production by CD19+ B-cells and Proliferation of CD4+ T-cells While Upregulating Tregs. J. Immunol. Res. 2017, 2017, 3475926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, S.C.; Quartel, A.W.; Czer, L.S.; Admon, D.; Chen, G.E.; Fishbein, M.C.; Schwieger, J.; Steiner, R.W.; Davis, C.; Tyan, D.B. Posttransplant therapy using high-dose human immunoglobulin (intravenous gammaglobulin) to control acute humoral rejection in renal and cardiac allograft recipients and potential mechanism of action. Transplantation 1998, 66, 800–805. [Google Scholar] [CrossRef]
- Jordan, S.C.; Vo, A.; Bunnapradist, S.; Toyoda, M.; Peng, A.; Puliyanda, D.; Kamil, E.; Tyan, D. Intravenous immune globulin treatment inhibits crossmatch positivity and allows for successful transplantation of incompatible organs in living-donor and cadaver recipients. Transplantation 2003, 76, 631–636. [Google Scholar] [CrossRef]
- Perez, E.E.; Orange, J.S.; Bonilla, F.; Chinen, J.; Chinn, I.K.; Dorsey, M.; El-Gamal, Y.; Harville, T.O.; Hossny, E.; Mazer, B.; et al. Update on the use of immunoglobulin in human disease: A review of evidence. J. Allergy Clin. Immunol. 2017, 139, S1–S46. [Google Scholar] [CrossRef] [Green Version]
- Jordan, S.C.; Toyoda, M.; Vo, A.A. Intravenous immunoglobulin a natural regulator of immunity and inflammation. Transplantation 2009, 88, 1–6. [Google Scholar] [CrossRef]
- Zachary, A.A.; Montgomery, R.A.; Ratner, L.E.; Samaniego-Picota, M.; Haas, M.; Kopchaliiska, D.; Leffell, M.S. Specific and durable elimination of antibody to donor HLA antigens in renal-transplant patients. Transplantation 2003, 76, 1519–1525. [Google Scholar] [CrossRef]
- Nair, V.; Sawinski, D.; Akalin, E.; Friedlander, R.; Ebcioglu, Z.; Sehgal, V.; Dinavahi, R.; Khaim, R.; Ames, S.; Lerner, S.; et al. Effect of high-dose intravenous immunoglobulin on anti-HLA Abs in sensitized kidney transplant candidates. Clin. Transplant. 2012, 26, E261–E268. [Google Scholar] [CrossRef]
- Alachkar, N.; Lonze, B.E.; Zachary, A.A.; Holechek, M.J.; Schillinger, K.; Cameron, A.M.; Desai, N.M.; Dagher, N.N.; Segev, D.L.; Montgomery, R.A.; et al. Infusion of high-dose intravenous immunoglobulin fails to lower the strength of human leukocyte antigen Abs in highly sensitized patients. Transplantation 2012, 94, 165–171. [Google Scholar] [CrossRef]
- Marfo, K.; Ling, M.; Bao, Y.; Calder, B.; Ye, B.; Hayde, N.; Greenstein, S.; Chapochnick-Friedman, J.; Glicklich, D.; De Boccardo, G.; et al. Lack of effect in desensitization with intravenous immunoglobulin and rituximab in highly sensitized patients. Transplantation 2012, 94, 345–351. [Google Scholar] [CrossRef]
- ELHilali, F.; Jucaud, V.; ELHilali, H.; Bhuiyan, M.H.; Mancuso, A.; LiuSullivan, N.; Elidrissi, A.; Mazouz, H. Characterization of the Anti-HLA Class I and II IgG Abs in Moroccan IVIg Using Regular Beads and Ibeads in Luminex Multiplex Single Antigen Immunoassay. Int. J. Immunol. 2017, 5, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V.; Kawakita, S. Suppression of blastogenesis and proliferation of activated CD4(+) T-cells: Intravenous immunoglobulin (IVIg) versus novel anti-human leucocyte antigen (HLA)-E mAbs mimicking HLA-I reactivity of IVIg. Clin. Exp. Immunol. 2014, 178, 154–177. [Google Scholar] [CrossRef]
- Kaveri, S.; Vassilev, T.; Hurez, V.; Lengagne, R.; Lefranc, C.; Cot, S.; Pouletty, P.; Glotz, D.; Kazatchkine, M.D. Antibodies to a conserved region of HLA class I molecules, capable of modulating CD8 T cell-mediated function, are present in pooled normal immunoglobulin for therapeutic use. J. Clin. Investig. 1996, 97, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Klaesson, S.; Ringdén, O.; Markling, L.; Remberger, M.; Lundkvist, I. Immune modulatory effects of immunoglobulns on cellmediated immune responses in vitro. Scand. J. Immunol. 1993, 38, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, F.; Horiuchi, H.; Nagata, I.S.; Kitahara, S.; Kiyoki, M.; Komoriya, K.; Yuki, N. Fc portion of intravenous immunoglobulins suppresses the induction of experimental allergic neuritis. J. Neuroimmunol. 1997, 78, 127–131. [Google Scholar] [CrossRef]
- Sali, A.D.; Karakasiliotis, I.; Evangelidou, M.; Avarameas, S.; Lymberi, P. Immunological evidence and regulatory potential for cell-penetrating antibodies in intravenous immunoglobulin. Clin. Transl. Immunol. 2015, 4, e42. [Google Scholar] [CrossRef] [PubMed]
- van Schaik, I.N.; Lundkvist, I.; Vermeulen, M.; Brand, A. Polyvalent immunoglobulin for intravenous use interferes with cell proliferation in vitro. J. Clin. Immunol. 1992, 12, 325–334. [Google Scholar] [CrossRef]
- Sbrana, S.; Ruocco, L.; Vanacore, R.; Azzarà, A.; Ambrogi, F. In vitro effects of an immunoglobulin preparation for intravenous use (IVIG) on T-cells activation. Allerg. Immunol. 1993, 25, 35–37. [Google Scholar]
- Hurez, V.; Kaveri, S.V.; Mouhoub, A.; Dietrich, G.; Mani, J.C.; Klatzmann, D.; Kazatchkine, M.D. Anti-CD4 activity of normal human immunoglobulin G for therapeutic use (intravenous immunoglobulin, IVIg). Ther. Immunol. 1994, 1, 269–277. [Google Scholar] [PubMed]
- Amran, D.; Renz, H.; Lack, G.; Bradley, K.; Gelfand, E.W. Suppression of cytokine-dependent human T-cell proliferation by intravenous immunoglobulin. Clin. Immunol. Immunopathol. 1994, 73, 180–186. [Google Scholar] [CrossRef]
- Andersson, J.; Skansén-Saphir, U.; Sparrelid, E.; Andersson, U. Intravenous immune globulin affects cytokine production in T lymphocytes and monocytes/macrophages. Clin. Exp. Immunol. 1996, 104 (Suppl. 1), 10–20. [Google Scholar] [CrossRef]
- Prasad, N.K.; Papoff, G.; Zeuner, A.; Bonnin, E.; Kazatchkine, M.D.; Ruberti, G.; Kaveri, S.V. Therapeutic preparations of normal polyspecific IgG (IVIg) induce apoptosis in human lymphocytes and monocytes: A novel mechanism of action of IVIg involving the Fas apoptotic pathway. J. Immunol. 1998, 161, 3781–3790. [Google Scholar] [PubMed]
- Aktas, O.; Waiczies, S.; Grieger, U.; Wendling, U.; Zschenderlein, R.; Zipp, F. Polyspecific immunoglobulins (IVIg) suppress proliferation of human (auto)antigen-specific T-cells without inducing apoptosis. J. Neuroimmunol. 2001, 114, 160–167. [Google Scholar] [CrossRef]
- MacMillan, H.F.; Lee, T.; Issekutz, A.C. Intravenous immunoglobulin G-mediated inhibition of T-cell proliferation reflects an endogenous mechanism by which IgG modulates T-cell activation. Clin. Immunol. 2009, 132, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Aubin, E.; Lemieux, R.; Bazin, R. Indirect inhibition of in vivo and in vitro T-cell responses by intravenous immunoglobulins due to impaired antigen presentation. Blood 2010, 115, 1727–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawfik, D.S.; Cowan, K.R.; Walsh, A.M.; Hamilton, W.S.; Goldman, F.D. Exogenous immunoglobulin downregulates T-cell receptor signaling and cytokine production. Pediatr. Allergy Immunol. 2012, 23, 88–95. [Google Scholar] [CrossRef]
- Koffman, B.M.; Dalakas, M.C. Effect of high-dose intravenous immunoglobulin on serum chemistry, hematology, and lymphocyte subpopulations: Assessments based on controlled treatment trials in patients with neurological diseases. Muscle Nerve 1997, 20, 1102–1107. [Google Scholar] [CrossRef]
- Rodriguez, M.A.; De Sanctis, J.B.; Blasini, A.; Leon-Ponte, M.; Abadi, I. Human IFN-gamma up-regulates IL-2 receptors in mitogen-activated T lymphocytes. Immunology 1990, 69, 554–557. [Google Scholar]
- Santos, S.G.; Antoniou, A.N.; Sampaio, P.; Powis, S.J.; Arosa, F.A. Lack of tyrosine 320 impairs spontaneous endocytosis and enhances release of HLA-B27 molecules. J. Immunol. 2006, 176, 2942–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guild, B.C.; Strominger, J.L. Human and murine class I MHC antigens share conserved serine 335, the site of HLA phosphorylation in vivo. J. Biol. Chem. 1984, 259, 9235–9240. [Google Scholar] [CrossRef]
- Mustelin, T.; Vang, T.; Bottini, T.N. Protein tyrosine phosphatases and the immune response. Nat. Rev. Immunol. 2005, 5, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Buckley, R.H.; Schiff, R.I. The use of intravenous immune globulin in immuno-deficiency diseases. N. Engl. J. Med. 1991, 325, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Kaveri, S.V.; Dietrich, G.; Hurez, V.; Kazatchkine, M.D. Intravenous immunoglobulins (IVIg) in the treatment of autoimmune diseases. Clin. Exp. Immunol. 1991, 86, 192–198. [Google Scholar] [CrossRef]
- McIntyre, J.A.; Higgins, N.; Britton, R.; Faucett, S.; Johnson, S.; Beckman, D.; Hormuth, D.; Fehrenbacher, J.; Halbrook, H. Utilization of intravenous immunoglobulin to ameliorate all-Abs in a highly sensitized patient with a cardiac assist device awaiting heart transplantation. Fluorescence-activated cell sorter analysis. Transplantation 1996, 62, 691–693. [Google Scholar] [CrossRef]
- Ippoliti, F.; De Santis, W.; Volterrani, A.; Canitano, N.; Frattolillo, D.; Lucarelli, S.; Frediani, S.; Frediani, T. Psychological stress affects response to sublingual immunotherapy in asthmatic children allergic to house dust mite. Pediatr. Allergy Immunol. 2006, 17, 337–345. [Google Scholar] [CrossRef]
- Glotz, D.; Antoine, C.; Julia, P.; Suberbielle-Boissel, C.; Boudjeltia, S.; Fraoui, R.; Hacen, C.; Duboust, A.; Bariety, J. Desensitization and subsequent kidney transplantation of patients using intravenous immunoglobulins (IVIg). Am. J. Transplant. 2002, 2, 758–760. [Google Scholar] [CrossRef]
- Sethi, S.; Choi, J.; Toyoda, M.; Vo, A.; Peng, A.; Jordan, S.C. Desensitization: Overcoming the Immunologic Barriers to Transplantation. J. Immunol. Res. 2017, 2017, 6804678. [Google Scholar] [CrossRef]
- Jordan, S.C.; Tyan, D.; Stablein, D.; McIntosh, M.; Rose, S.; Vo, A.; Toyoda, M.; Davis, C.; Shapiro, R.; Adey, D.; et al. Evaluation of intravenous immunoglobulin as an agent to lower allosensitization and improve transplantation in highly sensitized adult patients with end-stage renal disease: Report of the NIH IG02 trial. J. Am. Soc. Nephrol. 2004, 15, 3256–3262. [Google Scholar] [CrossRef]
- Ross, C.N.; Gaskin, G.I.; Gregor-Macgregor, S.U.; Patel, A.A.; Davey, N.J.; Lechler, R.I.; Williams, G.O.; Rees, A.J.; Pusey, C.D. Renal transplantation following immunoadsorption in highly sensitized recipients. Transplantation 1993, 5, 785–789. [Google Scholar] [CrossRef]
- larabi, A.; Backman, U.; Wikstrom, B.; Sjoberg, O.; Tufveson, G. Pretransplantation plasmapheresis in HLA-sensitized patients: Five years experience. Transplant. Proc. 1995, 27, 3448. [Google Scholar]
- Stegall, M.D.; Gloor, J.; Winters, J.L.; Moore, S.B.; Degoey, S. A comparison of plasmapheresis versus high-dose IVIG desensitization in renal allograft recipients with high levels of donor specific alloantib ody. Am. J. Transplant. 2006, 6, 346–351. [Google Scholar] [CrossRef]
- Onrust, S.V.; Lamb, H.M.; Balfour, J.A. Rituximab. Drugs 1999, 58, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Vo, A.A.; Lukovsky, M.; Toyoda, M.; Wang, J.; Reinsmoen, N.L.; Lai, C.H.; Peng, A.; Villicana, R.; Jordan, S.C. Rituximab and intravenous immune globulin for desensitization during real transplantation. N. Engl. J. Med. 2008, 359, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Vo, A.A.; Peng, A.; Toyoda, M.; Kahwaji, J.; Cao, K.; Lai, C.H.; Reinsmoen, N.L.; Villicana, R.; Jordan, S.C. Use of intravenous immune globulin and rituximab for desensitization of highly HLA-sensitized patients awaiting kidney transplantation. Transplantation 2010, 89, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Vo, A.A.; Choi, J.; Cisneros, K.; Reinsmoen, N.; Haas, M.; Ge, S.; Toyoda, M.; Kahwaji, J.; Peng, A.; Villicana, R.; et al. Benefits of rituximab combine with intravenous immunoglobulin for desensitization of kidney transplant recipients. Transplantation 2014, 98, 312–319. [Google Scholar] [CrossRef]
- Zhu, D.; Ravindranath, M.H.; Terasaki, P.I.; Miyazaki, T.; Pham, T.; Jucaud, V. Suppression of allo-human leucocyteantigen (HLA) Abs secreted by B memory cells in vitro: Intravenous immunoglobulin (IVIg) versus a monoclonal anti-HLA-E IgG that mimics HLA-I reactivities of IVIg. Clin. Exp. Immunol. 2014, 177, 464–477. [Google Scholar] [CrossRef]
- Baecher-Allan, C.; Brown, J.A.; Freeman, G.J.; Hafler, D.A. CD4+CD25+ high regulatory cells in human peripheral blood. J. Immunol. 2001, 167, 1245–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, W.F.; Duggan, P.J.; Ponchel, F.; Matarese, G.; Lombardi, G.; Edwards, A.D.; Isaacs, J.D.; Lechler, R.I. Human CD4+CD25+ cells: A naturally occurring population of regulatory T-cells. Blood 2001, 98, 2736–2744. [Google Scholar] [CrossRef]
- Li, Y.; Koshiba, T.; Yoshizawa, A.; Yonekawa, Y.; Masuda, K.; Ito, A.; Ueda, M.; Mori, T.; Kawamoto, H.; Tanaka, Y.; et al. Analyses of peripheral blood mononuclear cells in operational tolerance after pediatric living donor liver Transplantation. Am. J. Transplant. 2004, 4, 2118–2125. [Google Scholar] [CrossRef] [Green Version]
- Pons, J.A.; Revilla-Nuin, B.; Baroja-Mazo, A.; Ramírez, P.; Martínez-Alarcón, L.; Sánchez-Bueno, F.; Robles, R.; Rios, A.; Aparicio, P.; Parrilla, P. FoxP3 in peripheral blood is associated with operational tolerance in liver transplant patients during immunosuppression withdrawal. Transplantation 2008, 86, 1370–1378. [Google Scholar] [CrossRef]
- Baroja-Mazo, A.; Revilla-Nuin, B.; Parrilla, P.; Martinez-Alarcon, L.; Ramirez, P.; Pons, J.A. Tolerance in liver Transplantation: Biomarkers and clinical relevance. World J. Gastroenterol. 2016, 22, 7676–7691. [Google Scholar] [CrossRef] [PubMed]
- Baroja-Mazo, A.; Revilla-Nuin, B.; de Bejar, Á.; Martínez-Alarcón, L.; Herrero, J.I.; El-Tayeb, A.; Müller, C.E.; Aparicio, P.; Pelegrín, P.; Pons, J.A. Extracellular adenosine reversibly inhibits the activation of human regulatory T-cells and negatively influences the achievement of the operational tolerance in liver Transplantation. Am. J. Transplant. 2019, 9, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Jhun, J.; Lee, S.H.; Lee, S.K.; Kim, H.Y.; Jung, E.S.; Kim, D.G.; Choi, J.; Bae, S.H.; Yoon, S.K.; Chung, B.H.; et al. Serial monitoring of immune markers being represented regulatory T cell/T helper 17 cell ratio: Indicating tolerance for tapering immunosuppression after liver Transplantation. Front. Immunol. 2018, 9, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sablik, K.A.; Jordanova, E.S.; Pocorni, N.; Clahsen-van Groningen, M.C.; Betjes, M.G.H. Immune Cell Infiltrate in Chronic-Active Antibody-Mediated Rejection. Front. Immunol. 2019, 10, 3106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, H.; Zheng, Y.; Thomson, A.W.; Rogers, N.M. Transplant Tolerance Induction: Insights from the Liver. Front. Immunol. 2020, 11, 1044. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Kingsley, C.I.; Niimi, M.; Read, S.; Turvey, S.E.; Bushell, A.R.; Morris, P.J.; Powrie, F.; Wood, K.J. IL-10 is required for regulatory T-cells to mediate tolerance to alloantigens in vivo. J. Immunol. 2001, 166, 3789–3796. [Google Scholar] [CrossRef] [PubMed]
- Levings, M.; Sangregorio, R.; Roncarolo, M.G. Human CD25+CD4+ T regulatory cells suppress naive and memory T-cell proliferation and can be expanded in vitro without loss of function. J. Exp. Med. 2001, 193, 1295–1301. [Google Scholar] [CrossRef]
- van Maurik, K.; Wood, J.; Jones, N. Cutting edge: CD4+CD25+ alloantigen-specific immunoregulatory cells that can prevent CD8+ T-cell-mediated graft rejection: Implications for anti-CD154 immunotherapy. J. Immunol. 2002, 169, 5401–5404. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-Y.; Graca, L.; Cobbold, S.P.; Waldmann, H. Dominant transplantation tolerance impairs CD8+ T-cell function but not expansion. Nat. Immunol. 2002, 3, 1208–1213. [Google Scholar] [CrossRef]
- Graca, L.; Cobbold, S.P.; Waldmann, H. Identification of regulatory T-cells in tolerated allografts. J. Exp. Med. 2002, 195, 1641–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobbold, S.P.; Waldmann, H. Regulatory cells and transplantation tolerance. Cold Spring Harb. Perspect. Med. 2013, 3, a015545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, J.; Hashizume, M.; Kaneko, Y.; Yoshimoto, K.; Nishina, N.; Takeuchi, T. Peripheral blood CD4(+) CD25(+) CD127 (low) regulatory T-cells are significantly increased by tocilizumab treatment in patients with rheumatoid arthritis: Increase in regulatory T-cells correlates with clinical response. Arthritis Res. Ther. 2015, 17, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ephrem, A.; Chamat, S.; Miquel, C.; Fisson, S.; Mouthon, L.; Caligiuri, G.; Delignat, S.; Elluru, S.; Bayry, J.; Lacroix-Desmazes, S.; et al. Expansion of CD4+CD25+regulatory T-cells by intravenous immunoglobulin: A critical factor in controlling experimental autoimmune encephalomyelitis. Blood 2008, 111, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Filippone, E.J.; Das, B.; Norin, A.J.; Ravindranath, M.H. Optimizing the assessment of pathogenic anti-HLA antibodies. Am. J. Transplant. 2021, 21, 431–432. [Google Scholar] [CrossRef]
- Arosa, F.A.; Esgalhado, A.J.; Padrão, C.A.; Cardoso, E.M. Divide, Conquer, and Sense: CD8+CD28- T-cells in Perspective. Front. Immunol. 2017, 7, 665. [Google Scholar] [CrossRef] [Green Version]
- Arosa, F.A.; de Jesus, O.; Porto, G.; Carmo, A.M.; de Sousa, M. Calreticulin is expressed on the cell surface of activated human peripheral blood T lymphocytes in association with major histocompatibility complex class I molecules. J. Biol. Chem. 1999, 274, 16917–16922. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.L.; Trowsdale, J. Recognition of classical and heavy chain forms of HLA-B27 by leukocyte receptors. Curr. Mol. Med. 2004, 4, 59–65. [Google Scholar] [CrossRef]
- Jones, D.C.; Kosmoliaptsis, V.; Apps, R.; Lapaque, N.; Smith, I.; Kono, A.; Chang, C.; Boyle, L.H.; Taylor, C.J.; Trowsdale, J.; et al. HLA class I allelic sequence and conformation regulate leukocyte Ig-like receptor binding. J. Immunol. 2011, 186, 2990–2997. [Google Scholar] [CrossRef] [Green Version]
- Tamir, I.; Cambier, J.C. Antigen receptor signaling: Integration of protein tyrosine kinase functions. Oncogene 1988, 17, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Angulo, R.; Fulcher, D.A. Measurement of Candida-specific blastogenesis: Comparison of carboxyfluorescein succinimidyl ester labelling of T-cells, thymidine incorporation, and CD69 expression. Cytometry 1998, 34, 143–151. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | 43 | 44 | 45 | 46 | 47 | 48 | 49 | 50 | |
| HLA-E | G | S | H | S | L | K | Y | F | H | T | S | V | S | R | P | G | R | G | E | P | R | F | I | S | V | G | Y | V | D | D | T | Q | F | V | R | F | D | N | D | A | A | S | P | R | M | V | P | R | A | P |
| HLA-G | G | S | H | S | M | R | Y | F | S | A | A | V | S | R | P | G | R | G | E | P | R | F | I | A | M | G | Y | V | D | D | T | Q | F | V | R | F | D | S | D | S | A | C | P | R | M | E | P | R | A | P |
| HLA-F | G | S | H | S | L | R | Y | F | S | T | A | V | S | R | P | G | R | G | E | P | R | Y | I | A | V | E | Y | V | D | D | T | Q | F | L | R | F | D | S | D | A | A | I | P | R | M | E | P | R | E | P |
| a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | a1 | |||||||||||||||||||||
| 51 | 52 | 53 | 54 | 55 | 56 | 57 | 58 | 59 | 60 | 61 | 62 | 63 | 64 | 65 | 66 | 67 | 68 | 69 | 70 | 71 | 72 | 73 | 74 | 75 | 76 | 77 | 78 | 79 | 80 | 81 | 82 | 83 | 84 | 85 | 86 | 87 | 88 | 89 | 90 | 91 | 92 | 93 | 94 | 95 | 96 | 97 | 98 | 99 | 100 | |
| HLA-E | W | M | E | Q | E | G | S | E | Y | W | D | R | E | T | R | S | A | R | D | T | A | Q | L | F | R | V | N | L | R | T | L | R | G | Y | Y | N | Q | S | E | A | G | S | H | T | L | Q | W | M | H | G |
| HLA-G | W | V | E | Q | E | G | P | E | Y | W | E | E | E | T | R | N | T | K | A | H | A | Q | T | D | R | M | N | L | Q | T | L | R | G | Y | Y | N | Q | S | E | A | S | S | H | T | L | Q | W | M | I | G |
| HLA-F | W | V | E | Q | E | G | P | Q | Y | W | E | W | T | T | G | Y | A | K | A | N | A | Q | T | D | R | V | A | L | R | N | L | L | R | R | Y | N | Q | S | E | A | G | S | H | T | L | Q | G | M | N | G |
| a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | |||||||||||||||||
| 101 | 102 | 103 | 104 | 105 | 106 | 107 | 108 | 109 | 110 | 111 | 112 | 113 | 114 | 115 | 116 | 117 | 118 | 119 | 120 | 121 | 122 | 123 | 124 | 125 | 126 | 127 | 128 | 129 | 130 | 131 | 132 | 133 | 134 | 135 | 136 | 137 | 138 | 139 | 140 | 141 | 142 | 143 | 144 | 145 | 146 | 147 | 148 | 149 | 150 | |
| HLA-E | C | E | L | G | P | D | R | R | F | L | R | G | Y | E | Q | F | A | Y | D | G | K | D | Y | L | T | L | N | E | D | L | R | S | W | T | A | V | D | T | A | A | Q | I | S | E | G | K | S | N | D | A |
| HLA-G | C | D | L | G | S | D | G | R | L | L | R | G | Y | E | Q | Y | A | Y | D | G | K | D | Y | L | A | L | N | E | D | L | R | S | W | T | A | A | D | T | A | A | Q | I | S | K | R | K | C | E | A | A |
| HLA-F | C | D | M | G | P | D | G | R | L | L | R | G | Y | H | Q | H | A | Y | D | G | K | D | Y | I | S | L | N | E | D | L | R | S | W | T | A | A | D | T | V | A | Q | I | T | Q | R | F | Y | E | A | E |
| SHARED WITH HLA-A/-B & -C | SHARED WITH HLA-A, HLA-B & HLA-C | |||||||||||||||||||||||||||||||||||||||||||||||||
| a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | a2 | |||||||||||||||||||||||||||||||
| 151 | 152 | 153 | 154 | 155 | 156 | 157 | 158 | 159 | 160 | 161 | 162 | 163 | 164 | 165 | 166 | 167 | 168 | 169 | 170 | 171 | 172 | 173 | 174 | 175 | 176 | 177 | 178 | 179 | 180 | 181 | 182 | 183 | 184 | 185 | 186 | 187 | 188 | 189 | 190 | 191 | 192 | 193 | 194 | 195 | 196 | 197 | 198 | 199 | 200 | |
| HLA-E | S | E | A | E | H | Q | R | A | Y | L | E | D | T | C | V | E | W | L | H | K | Y | L | E | K | G | K | E | T | L | L | H | L | E | P | P | K | T | H | V | T | H | H | P | I | S | D | H | E | A | T |
| HLA-G | N | V | A | E | Q | R | R | A | Y | L | E | G | T | C | V | E | W | L | H | R | Y | L | E | N | G | K | E | M | L | Q | R | A | D | P | P | K | T | H | V | T | H | H | P | V | F | D | Y | E | A | T |
| HLA-F | E | Y | A | E | E | F | R | T | Y | L | E | G | E | C | L | E | L | L | R | R | Y | L | E | N | G | K | E | T | L | Q | R | A | D | P | P | K | A | H | V | A | H | H | P | I | S | D | H | E | A | T |
| 201 | 202 | 203 | 204 | 205 | 206 | 207 | 208 | 209 | 210 | 211 | 212 | 213 | 214 | 215 | 216 | 217 | 218 | 219 | 220 | 221 | 222 | 223 | 224 | 225 | 226 | 227 | 228 | 229 | 230 | 231 | 232 | 233 | 234 | 235 | 236 | 237 | 238 | 239 | 240 | 241 | 242 | 243 | 244 | 245 | 246 | 247 | 248 | 249 | 250 | |
| HLA-E | L | R | C | W | A | L | G | F | Y | P | A | E | I | T | L | T | W | Q | Q | D | G | E | G | H | T | Q | D | T | E | L | V | E | T | R | P | A | G | D | G | T | F | Q | K | W | A | A | V | V | V | P |
| HLA-G | L | R | C | W | A | L | G | F | Y | P | A | E | I | I | L | T | W | Q | R | D | G | E | D | Q | T | Q | D | V | E | L | V | E | T | R | P | A | G | D | G | T | F | Q | K | W | A | A | V | V | V | P |
| HLA-F | L | R | C | W | A | L | G | F | Y | P | A | E | I | T | L | T | W | Q | R | D | G | E | E | Q | T | Q | D | T | E | L | V | E | T | R | P | A | G | D | G | T | F | Q | K | W | A | A | V | V | V | P |
| HLA-E Peptide Sequences | HLA Alleles | Method 1 | Method 2 | Method 3 | Method 4 | Rank of Antigenicity | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Prediction SCORES | |||||||||||
| Classical HLA-Ib | Non-Classical HLA-Ib | Specificity | Beta-Turn | Antigenicity | Flexibility | Hydrophilicity | |||||
| [total number of amino acids] | A | B | Cw | F | G | Chou & Fasman (1978) | Kolaskar & Tangaonkar (1990) | Karplus & Schulz (1985) | Parker (1986) | ||
| 47PRAPWMEQE55 [9] | 1 | 0 | 0 | 0 | 0 | A*3306 | 0.993 | 0.948 | 0.969 | 0.586/1.143/1.657 | |
| 58EYWDRETR65 [8] | 5 | 0 | 0 | 0 | 0 | A restricted | 0.993 | 0.915 | 1.024 | 3.301/2.786 | 10 |
| 90AGSHTLQW97 [8] | 1 | 10 | 48 | 0 | 0 | Polyspecific | 1.019 | 1.033 | 0.989 | 2.629/0.901 | 6 |
| 108RFLRGYE114 [7] | 24 | 0 | 0 | 0 | 0 | A restricted | 0.933 | 0.996 | 0.996 | 0.229 | 8 |
| 115QFAYDGKDY123 [9] | 1 | 104 | 75 | 0 | 0 | Polyspecific | 1.059 | 1.001 | 0.993 | 2.629/3.201 | 5 |
| 117AYDGKDY123 [7] | 491 | 831 | 271 | 21 | 30 | Polyspecific | 1.204 | 0.989 | 1.061 | 4.243 | 1 |
| 126LNEDLRSWTA135 [10] | 239 | 219 | 261 | 21 | 30 | Polyspecific | 1.046 | 0.983 | 1.039 | 2.443/2.329 | 2 |
| 137DTAAQI142 [6] | 0 | 824 | 248 | 0 | 30 | Polyspecific | 0.813 | 1.065 | 0.978 | 1.957 | 3 |
| 137DTAAQIS143 [7] | 0 | 52 | 4 | 0 | 30 | Polyspecific | 0.946 | 1.012 | 0.97 | 3.414 | 7 |
| 157RAYLED162 [6] | 0 | 1 | 0 | 0 | 0 | B*8201 | 0.929 | 0.996 | 0.969 | 2.601 | |
| 163TCVEWL168 [6] | 282 | 206 | 200 | 0 | 30 | Polyspecific | 0.841 | 1.115 | 0.929 | −0.914 | 4 |
| 183EPPKTHVT190 [8] | 0 | 0 | 19 | 0 | 0 | C restricted | 1.029 | 1.044 | 1.042 | 3.043 | 9 |
| 65RSARDTA71 [7] | 0 | 0 | 0 | 0 | 0 | E restricted | 1.011 | 0.952 | 1.038 | 4.901 | 2 |
| 143SEQKSNDASE152 [10] | 0 | 0 | 0 | 0 | 0 | E restricted | 1.231 | 0.923 | 1.222 | 7.071/6.443/6.257/6.514 | 1 |
| IMMUNOGEN HLA-ER107 OR HLA-EG107 | |||||||
|---|---|---|---|---|---|---|---|
| Groups | Number | HLA-CLASS Ib | HLA-CLASS IC | ||||
| of mAbs | of mAbs | HLA-E | HLA-F | HLA-G | HLA-A | HLA-B | HLA-C |
| Group 1 | 24 | + | − | − | − | − | − |
| Group 2 | 1 | + | + | − | − | − | − |
| Group 3 | 1 | + | − | + | − | − | − |
| Group 4 | 8 | + | + | + | − | − | − |
| Group 5 | 4 | + | − | − | − | + | − |
| Group 6 | 31 | + | − | − | − | + | + |
| Group 7 | 109 | + | − | − | + | + | + |
| Group 8 | 11 | + | + | − | + | + | + |
| Group 9 | 18 | + | − | + | + | + | + |
| Group 10 | 7 | + | + | + | + | + | + |
| mAbs | Monospecific | Polyreactive | mAbs | Monospecific | Polyreactive | mAbs | Monospecific | Polyreactive | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TFL-033 | TFL-006 | TFL-007 | TFL-033 | TFL-006 | TFL-007 | TFL-033 | TFL-006 | TFL-007 | |||
| IgG1 | IgG2a | IgG2a | IgG1 | IgG2a | IgG2a | IgG1 | IgG2a | IgG2a | |||
| Neg | 3 | 15 | 7 | B* alleles | C* alleles | ||||||
| Pos | 71 | 88 | 85 | B*0702 | 1331 | 841 | C*0102 | 7242 | 3268 | ||
| HLA-E | 24411 | 22522 | 21618 | B*0801 | 2092 | 1033 | C*0202 | 10690 | 6084 | ||
| HLA-F | 12650 | 11035 | B*1301 | 5654 | 3979 | C*0302 | 5917 | 3062 | |||
| HLA-G | 7193 | 2670 | B*1302 | 2237 | 1426 | C*0303 | 7114 | 4250 | |||
| A* alleles | B*1401 | 11319 | 8767 | C*0304 | 6584 | 3891 | |||||
| A*0101 | 2395 | 1037 | B*1402 | 4414 | 2558 | C*0401 | 2843 | 1272 | |||
| A*0201 | 856 | B*1501 | 1097 | C*0501 | 16131 | 13096 | |||||
| A*0203 | 1095 | B*1502 | 6256 | 4497 | C*0602 | 9396 | 4274 | ||||
| A*0206 | 1494 | 843 | B*1503 | 2831 | 1926 | C*0702 | 12251 | 6919 | |||
| A*0301 | 818 | B*1510 | 2616 | 1470 | C*0801 | 13456 | 10733 | ||||
| A*1101 | 10190 | 8476 | B*1511 | 9041 | 5902 | C*1203 | 5055 | 2102 | |||
| A*1102 | 860 | B*1512 | 1624 | 996 | C*1402 | 8727 | 4936 | ||||
| A*2301 | 614 | B*1513 | 5326 | 3365 | C*1502 | 6030 | 3225 | ||||
| A*2402 | 3133 | 2011 | B*1516 | 5614 | 3443 | C*1601 | 8462 | 4364 | |||
| A*2403 | 3151 | 1967 | B*1801 | 6990 | 4890 | C*1701 | 13521 | 9069 | |||
| A*2501 | 1230 | 692 | B*2705 | 2591 | 1576 | C*1802 | 17918 | 15207 | |||
| A*2601 | 3368 | 1638 | B*2708 | 4437 | 2671 | C* alleles | 0 | 16 | 16 | ||
| A*2901 | 3194 | 2256 | B*3501 | 10205 | 8594 | ||||||
| A*2902 | 2235 | 1136 | B*3701 | 6472 | 4338 | ||||||
| A*3001 | 2229 | 1237 | B*3801 | 3844 | 1820 | ||||||
| A*3002 | 3353 | 2211 | B*3901 | 7093 | 5304 | ||||||
| A*3101 | 858 | B*4001 | 5743 | 3758 | |||||||
| A*3201 | 2237 | 1508 | B*4002 | 6118 | 4675 | ||||||
| A*3301 | 2791 | 1627 | B*4006 | 15643 | 13758 | ||||||
| A*3303 | 4212 | 2961 | B*4101 | 7191 | 5277 | ||||||
| A*3401 | 6268 | 3968 | B*4201 | 636 | |||||||
| A*3402 | 1399 | 893 | B*4402 | 7062 | 4059 | ||||||
| A*3601 | 5806 | 3826 | B*4403 | 7256 | 5638 | ||||||
| A*4301 | 4420 | 2364 | B*4501 | 9535 | 7646 | ||||||
| A*6601 | 3644 | 1526 | B*4601 | 6491 | 4130 | ||||||
| A*6602 | 1395 | 789 | B*4701 | 6528 | 3895 | ||||||
| A*6801 | 1314 | 859 | B*4801 | 4365 | 2716 | ||||||
| A*6802 | 2078 | 1276 | B*4901 | ||||||||
| A*6901 | 1964 | 917 | B*5001 | 741 | |||||||
| A*7401 | 723 | B*5101 | 6205 | 3724 | |||||||
| A*8001 | 2841 | 1430 | B*5102 | 5251 | 3579 | ||||||
| A* alleles | 0 | 32 | 24 | B*5201 | 4524 | 2728 | |||||
| B*5301 | 8807 | 7323 | |||||||||
| B*5401 | 5556 | 4153 | |||||||||
| B*5501 | 2829 | 1887 | |||||||||
| B*5601 | 1386 | 777 | |||||||||
| B*5701 | |||||||||||
| B*5703 | 1229 | 600 | |||||||||
| B*5801 | 10160 | 8047 | |||||||||
| B*5901 | 5646 | 3001 | |||||||||
| B*6701 | 675 | ||||||||||
| B*7301 | 3347 | 2171 | |||||||||
| B*7801 | 6089 | 4597 | |||||||||
| B*8101 | 1352 | 729 | |||||||||
| B*8201 | 4367 | 3069 | |||||||||
| B* alleles | 0 | 48 | 44 | ||||||||
| TFL-006 (20 ug/mL) | |||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HLA-A | NC | PC | A*01:01 | A*02:01 | A*02:03 | A*03:01 | A*11:01 | A*11:02 | A*23:01 | A*24:02 | A*24:03 | A*25:01 | A*26:01 | A*29:01 | A*29:02 | A*30:01 | A*31:01 | A*32:01 | A*33:01 | A*33:03 | A*34:02 | A*36:01 | A*43:01 | A*66:01 | A*66:02 | A*68:01 | A*68:02 | A*69:01 | A*74:01 | A*80:01 | |||||||||||||
| REGULAR LABSCreen BEADSET (Lot: LS1A04-lot 10) [Closed Conformers admixed with Open Conformers] | |||||||||||||||||||||||||||||||||||||||||||
| 0 | 12 | 933 | 339 | 1018 | 193 | 4782 | 537 | 133 | 716 | 2516 | 194 | 2221 | 1017 | 778 | 1496 | 396 | 515 | 1038 | 554 | 1535 | 1353 | 2479 | 1886 | 1454 | 713 | 1185 | 3128 | 652 | 3132 | ||||||||||||||
| LIFECODES BEADSET (Lot # 3005619) (Closed Conformers only) | |||||||||||||||||||||||||||||||||||||||||||
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||||||||||||||
| HLA-B | B*07:02 | B*08:01 | B*13:02 | B*14:01 | B*14:02 | B*15:01 | B*15:02 | B*15:03 | B*15:12 | B*15:13 | B*15:16 | B*18:01 | B*27:05 | B*27:08 | B*35:01 | B*37:01 | B*38:01 | B*39:01 | B*40:01 | B*40:02 | B*41:01 | B*42:01 | B*44:02 | B*44:03 | B*45:01 | B*46:01 | B*47:01 | B*48:01 | B*49:01 | B*50:01 | B*51:01 | B*52:01 | B*53:01 | B*54:01 | B*55:01 | B*56:01 | B*57:01 | B*58:01 | B*59:01 | B*67:01 | B*73:01 | B*78:01 | B*81:01 |
| REGULAR LABSCreen BEADSET (Lot: LS1A04-lot 10) [Closed Conformers admixed with Open Conformers] | |||||||||||||||||||||||||||||||||||||||||||
| 862 | 1226 | 2514 | 7805 | 1831 | 335 | 1935 | 1822 | 770 | 3135 | 3076 | 3096 | 634 | 1659 | 6128 | 2650 | 2521 | 704 | 3429 | 2697 | 3739 | 347 | 3650 | 1829 | 1736 | 3572 | 2152 | 3262 | 1554 | 1799 | 2461 | 2146 | 5442 | 1662 | 2519 | 3662 | 2089 | 5268 | 3553 | 406 | 1423 | 2996 | 1525 | |
| LIFECODES BEADSET (Lot # 3005619) (Closed Conformers only) | |||||||||||||||||||||||||||||||||||||||||||
| 0 | 0 | 12 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 8 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | |
| HLA-C | C*01:02 | C*02:02 | C*03:03 | C*03:04 | C*04:01 | C*05:01 | C*06:02 | C*07:02 | C*08:01 | C*14:02 | C*15:02 | C*16:01 | C*17:01 | ||||||||||||||||||||||||||||||
| REGULAR LABSCreen BEADSET (Lot: LS1A04-lot 10) [Closted Conformers admixed with Open Conformers] | |||||||||||||||||||||||||||||||||||||||||||
| 4066 | 7446 | 2458 | 4504 | 3337 | 9124 | 5644 | 8702 | 6090 | 3937 | 4465 | 4648 | 8296 | |||||||||||||||||||||||||||||||
| LIFECODES BEADSET (Lot # 3005619) (Closed Conformers only) | |||||||||||||||||||||||||||||||||||||||||||
| 1 | 20 | 0 | 0 | 8 | 24 | 4 | 97 | 1 | 0 | 4 | 0 | 10 | |||||||||||||||||||||||||||||||
| Therapeutic Preparations of IVIg | Reactivity of Different HLA Class I Antigens | |||||
|---|---|---|---|---|---|---|
| Classical HLA-Ia Alleles | Non-Classical HLA-Ib | |||||
| A | B | Cw | E | F | G | |
| IVIg (GamaSTAN, Talecris Biotherapeutics, Inc., Research Triangle Park, NC, USA) | 31 | 50 | 16 | Positive | Positive | Positive |
| IVIg (Octogam, Octapharma S.A. Argentina Poniente, Mexico, D.F.) | 30 | 47 | 16 | Positive | Positive | Positive |
| IVIg (Sandoglobulin, CSL Behring, Kankakee, IL, USA) | 30 | 47 | 16 | Positive | Positive | Positive |
| IVIg (GlobEx, Bangalore, India) | 20 | 39 | 16 | Positive | Positive | Positive |
| IVIg (IV-LFB-CNTs LFB Biomedicaments, Courtaboeuf Cedex, France) | 31 | 50 | 16 | Positive | Positive | Positive |
| Source, Nature, and Functions | Intravenous Immunoglobulin (IVIg) | HLA-I Polyreactive mAbs, TFL-006 & TFL-007 |
| Manufacturer | Several pharmaceutical firms | Terasaki Foundation Laboratory, U.S. Patent No. 10,800.847;10/13/20 |
| Source | Purified from pooled plasma of 10,000 blood donors | Immunized in mice with a heavy chain of HLA-ER107 |
| Nature of antibody | Human, polyclonal IgG with trace levels of IgA | Murine purified monoclonal IgG |
| Subclass of IgG antibodies | IgG1, IgG2a, IgG3, IgG4 | IgG2a |
| Purity | Contains soluble HLA antigens and other non-IgG proteins | 100% purified protein of IgG2a [44,45] |
| Cytokines, chemokines | ||
| Antibody reactivity | CCs & OCs of HLA-A, HLA-B, HLA-Cw, HLA-E, HLA-F | OCs but not CCs of HLA-A, HLA-B, HLA-Cw, HLA-E, HLA-F, HLA-G |
| HLA-G, HLA-DR, HLA-DQA/DQB, HLA-DPA/DPB | None | |
| Fc receptors: FcgI, FcgII, FcgIII, FcgIV (tested) [96] | FcgII (anticipated) | |
| Blood groups: A, B, Rh | No | |
| Escherichia coli bacterial antigens ranging from | No | |
| antigens by different preparations of IVIg | No | |
| Human albumin | No | |
| Phospholipids | No | |
| Binding site | Binds to both closed and open conformers | Binds only to open conformers |
| Stabilizer | Many, including sucrose in some preparations | None |
| Protein concentration | Highly variable, from 2 to 12% | Protein concentration adjusted to requirement |
| CD4+ T-cell suppression | PHA- or cytokine-activated T cells by apoptosis and necrosis | PHA-activated T cells |
| CD8+ T-cell proliferation | PHA-activated T-cells | PHA-activated T cells |
| B-cell proliferation | May induce differentiation | None |
| Anti-HLA antibody suppression | PRA antibody reduction | Suppress production of anti-HLA-I and anti-HLA-II IgG |
| Suppress selected HLA-II antibody production | ||
| Promote selected HLA-II antibody production | ||
| Expansion of Tregs | Yes | Yes |
| Special application | Not applicable | To monitor the presence of open conformers admixed with |
| closed conformers on the bead sets. (e.g., LABScreen vs | ||
| LIFECODE bead sets used in monitoring HLA antibodies |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravindranath, M.H.; Hilali, F.E.; Filippone, E.J. Therapeutic Potential of HLA-I Polyreactive mAbs Mimicking the HLA-I Polyreactivity and Immunoregulatory Functions of IVIg. Vaccines 2021, 9, 680. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060680
Ravindranath MH, Hilali FE, Filippone EJ. Therapeutic Potential of HLA-I Polyreactive mAbs Mimicking the HLA-I Polyreactivity and Immunoregulatory Functions of IVIg. Vaccines. 2021; 9(6):680. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060680
Chicago/Turabian StyleRavindranath, Mepur H., Fatiha El Hilali, and Edward J. Filippone. 2021. "Therapeutic Potential of HLA-I Polyreactive mAbs Mimicking the HLA-I Polyreactivity and Immunoregulatory Functions of IVIg" Vaccines 9, no. 6: 680. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9060680