Serotonin and Tryptophan Serum Concentrations in Shelter Dogs Showing Different Behavioural Responses to a Potentially Stressful Procedure


{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Setting and Procedure
2.3. Blood Collection, Storage and Analysis
2.4. Statistical Analysis
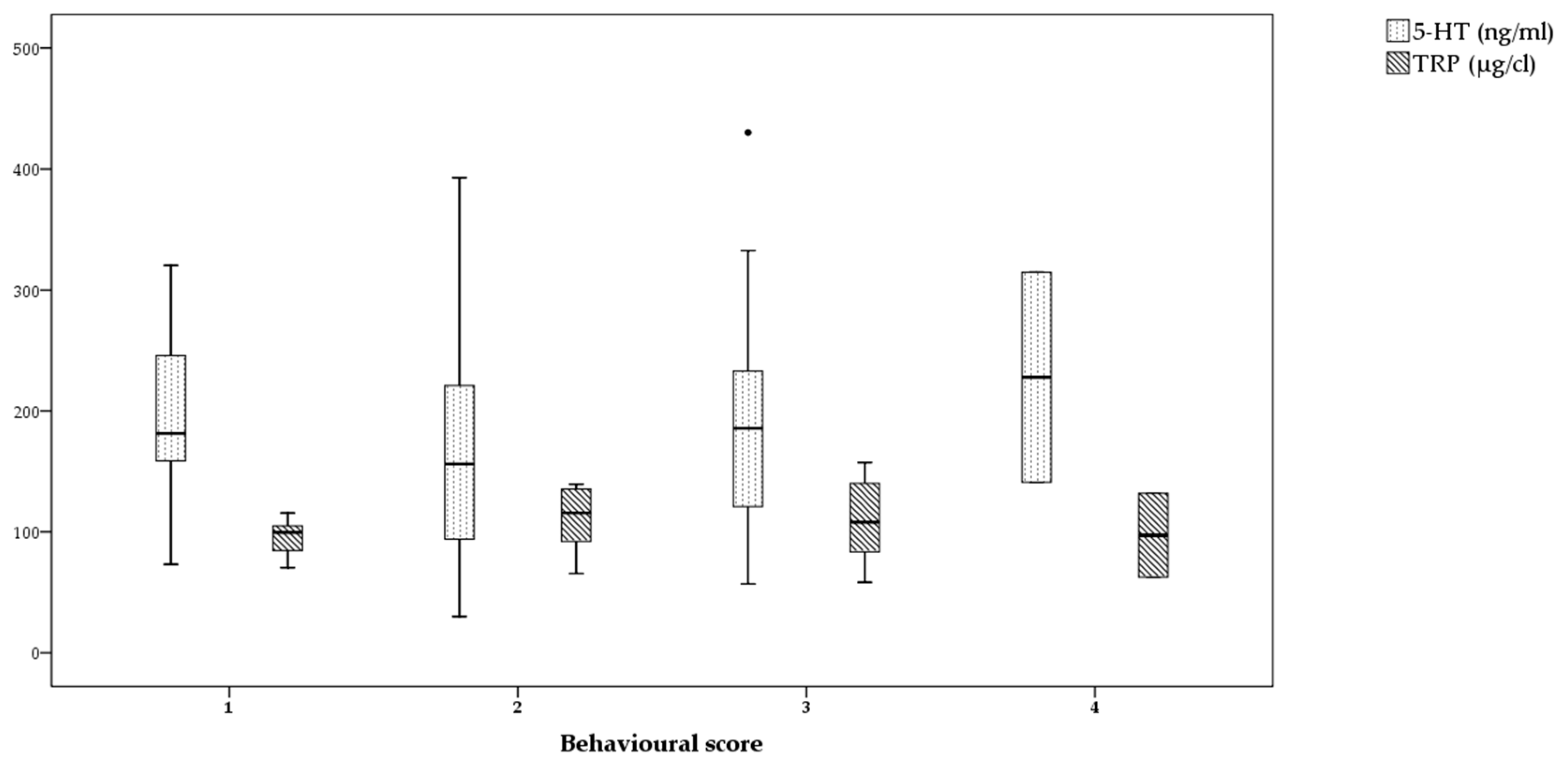
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bocchio, M.; McHugh, S.B.; Bannerman, D.M.; Sharp, T.; Capogna, M. Serotonin, Amygdala and Fear: Assembling the Puzzle. Front. Neural Circuits 2016, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.A.; Sun, E.W.; Martin, A.M.; Keating, D.J. The ever-changing roles of serotonin. Int. J. Biochem. Cell Biol. 2020, 125, 105776. [Google Scholar] [CrossRef] [PubMed]
- Lowry, C.A.; Johnson, P.L.; Hay-schmidt, A.; Mikkelsen, J.; Shekhar, A. Modulation of anxiety circuits by serotonergic systems. Stress 2005, 8, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, P.; Palanza, P.; Parmigiani, S.; De Almeida, R.M.M.; Miczek, K.A. Serotonin and aggressive behavior in rodents and nonhuman primates: Predispositions and plasticity. Eur. J. Pharmacol. 2005, 526, 259–273. [Google Scholar] [CrossRef]
- Howell, S.; Westergaard, G.; Hoos, B.; Chavanne, T.J.; Shoaf, S.E.; Cleveland, A.; Snoy, P.J.; Suomi, S.J.; Higley, J.D.E.E. Serotonergic Influences on Life-History Outcomes in Free-Ranging Male Rhesus Macaques. Am. J. Primatol. 2007, 89, 851–865. [Google Scholar] [CrossRef]
- Goveas, J.S.; Csernansky, J.G.; Coccaro, E.F. Platelet serotonin content correlates inversely with life history of aggression in personality-disordered subjects. Psychiatry Res. 2004, 126, 23–32. [Google Scholar] [CrossRef]
- Lidberg, L.; Tuck, J.R.; Asberg, M.; Scalia-Tomba, G.P.; Bertilsson, L. Homicide, suicide and CSF 5-HIAA. Acta Psychiatr. Scand. 1985, 71, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Ursinus, W.W.; Bolhuis, J.E.; Zonderland, J.J.; Rodenburg, T.B.; De Souza, A.S.; Koopmanschap, R.E.; Kemp, B.; Korte-bouws, G.A.H.; Korte, S.M.; Reenen, C.G. Van Relations between peripheral and brain serotonin measures and behavioural responses in a novelty test in pigs. Physiol. Behav. 2013, 118, 88–96. [Google Scholar] [CrossRef]
- Esler, M.; Lambert, E.; Alvarenga, M.; Socratous, F.; Richards, J.; Barton, D.; Pier, C.; Brenchley, C.; Dawood, T.Y.E.; Hastings, J.; et al. Increased brain serotonin turnover in panic disorder patients in the absence of a panic attack: Reduction by a selective serotonin reuptake inhibitor. Stress 2007, 10, 295–304. [Google Scholar] [CrossRef]
- Çakiroglu, D.; Meral, Y.; Sancak, A.A.; Çifti, G. Relationship between the serum concentrations of serotonin and lipids and aggression in dogs. Vet. Rec. 2007, 161, 59–61. [Google Scholar] [CrossRef]
- Leòn, M.; Rosado, B.; Garcìa-Belenguer, S.; Chacòn, G.; Villegas, A.; Palacio, J. Assessment of serotonin in serum, plasma, and platelets of aggressive dogs. J. Vet. Behav. Clin. Appl. Res. 2012, 7, 348–352. [Google Scholar] [CrossRef]
- Rosado, B.; Garcìa-Belenguer, S.; Leòn, M.; Chacòn, G.; Villegas, A.; Palacio, J. Blood concentrations of serotonin, cortisol and dehydroepiandrosterone in aggressive dogs. Appl. Anim. Behav. Sci. 2010, 123, 124–130. [Google Scholar] [CrossRef]
- Amat, M.; Le Brech, S.; Camps, T.; Torrente, C.; Mariotti, V.M.; Le Brech, S.; Ruiz, L.; Manteca, X. Differences in serotonin serum concentration between aggressive English cocker spaniels and aggressive dogs of other breeds. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 19–25. [Google Scholar] [CrossRef]
- Reisner, I.R.; Mann, J.J.; Stanley, M.; Huang, Y.; Houpt, K.A. Comparison of cerebrospinal fluid monoamine metabolite levels in dominant-aggressive and non-aggressive dogs. Brain Res. 1996, 714, 57–64. [Google Scholar] [CrossRef]
- Riva, J.; Bondiolotti, G.; Michelazzi, M.; Verga, M.; Carenzi, C. Anxiety related behavioural disorders and neurotransmitters in dogs. Appl. Anim. Behav. Sci. 2008, 114, 168–181. [Google Scholar] [CrossRef]
- Friedman, M. Analysis, Nutrition, and Health Benefits of Tryptophan. Int. J. Tryptophan Res. 2018, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, T.A.; Nguyen, J.C.D.; Polglaze, K.E.; Bertrand, P.P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef]
- Höglund, E.; Øverli, Ø.; Winberg, S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef]
- Gazzano, A.; Ogi, A.; Torracca, B.; Mariti, C.; Casini, L. Plasma Tryptophan/Large Neutral Amino Acids Ratio in Domestic Dogs Is Affected by a Single Meal with High Carbohydrates Level. Animals 2018, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Fernstrom, J.D. Review Aromatic amino acids and monoamine synthesis in the central nervous system: Influence of the diet. Hournal Nutr. Biochem. 1990, 1, 508–517. [Google Scholar] [CrossRef]
- Mateos, S.S.; Sánchez, C.L.; Paredes, S.D.; Barriga, C.; Rodríguez, A.B. Circadian Levels of Serotonin in Plasma and Brain after Oral Administration of Tryptophan in Rats. Basic Clin. Pharmacol. Toxicol. 2008, 104, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Kantak, K.M.; Hegstrand, L.R.; Whitman, J.; Eichelman, B. Effects of Dietary Supplements and a Tryptophan-Free Diet on Aggressive Behavior in Rats. Pharmacol. Biochem. Behav. 1980, 12, 173–179. [Google Scholar] [CrossRef]
- Chamberlain, B.; Ervin, F.R.; Pihl, R.O.; Youg, S.N. The Effect of Raising or Lowering Tryptophan Levels on Aggression in Vervet Monkeys. Pharmacol. Biochem. Behav. 1987, 28, 503–510. [Google Scholar] [CrossRef]
- Rouvinen, K.; Archbold, S.; Laffin, S.; Harri, M. Long-term effects of tryptophan on behavioural response and growing-furring performance in silver fox (Vulpes vulpes). Applied 1999, 63, 65–77. [Google Scholar] [CrossRef]
- Weld, K.P.; Mench, J.A.; Woodward, A.; Bolesta, M.S.; Suomi, S.J.; Higley, J.D. Effect of Tryptophan Treatment on Self-Biting and Central Nervous System Serotonin Metabolism in Rhesus Monkeys (Macaca mulatta). Neuropsychopharmacology 1998, 19, 314–321. [Google Scholar] [CrossRef]
- Koopmans, S.J.; Ruis, M.; Dekker, R.; Van Diepen, H.; Korte, M.; Mroz, Z.; Korte, S.M. Surplus dietary tryptophan reduces plasma cortisol and noradrenaline concentrations and enhances recovery after social stress in pigs. Physiol. Behav. 2005, 85, 469–478. [Google Scholar] [CrossRef]
- DeNapoli, J.S.; Dodman, N.H.; Shuster, L.; Rand, W.M.; Gross, K.L. Effect of dietary protein content and tryptophan supplementation on dominance aggression, territorial aggression, and hyperactivity in dogs. J. Am. Vet. Med. Assoc. 2000, 217, 505–508. [Google Scholar] [CrossRef]
- Anzola, B.; Ibañez, M.; Morillas, S.; Benedetti, R.; Pérez, J.; Farìas, D. The Use of Tryptophan in Shelter Dogs to Treat Stress-related Anxiety Disorders DE. Rev. Cient. Vet. 2013, 1, 26–32. [Google Scholar]
- Kaulfuß, P.; Drescher, D.; Failing, K.; Hintze, S.; Würbel, H. The effect of trypthophan as dietary supplementation on dogs with abnormal-repetitive behaviours. Prakt. Tierarzt 2014, 95, 24–39. [Google Scholar]
- Bosch, G.; Beerda, B.; Beynen, A.C.; van der Borg, J.A.M.; van der Poel, A.F.B.; Hendriks, W.H. Dietary tryptophan supplementation in privately owned mildly anxious dogs. Appl. Anim. Behav. Sci. 2009, 121, 197–205. [Google Scholar] [CrossRef]
- Templeman, J.R.; Davenport, G.M.; Cant, J.P.; Osborne, V.R.; Shoveller, A. The effect of graded concentrations of dietary tryptophan on canine behavior in response to the approach of a familiar or unfamiliar individual. Can. J. Vet. Res. 2018, 82, 294–305. [Google Scholar] [PubMed]
- Pietraszek, M.H.; Takada, Y.; Yan, D.; Urano, T.; Senzawa, K.; Takada, A. Relationship between serotoninergic measurs in periphery and the brain of mouse. Life Sci. 1992, 51, 75–82. [Google Scholar] [CrossRef]
- Gazzano, A.; Ogi, A.; Macchioni, F.; Gatta, D.; Preziuso, G.; Baragli, P.; Curadi, M.C.; Giuliotti, L.; Sergi, V.; Casini, L. Blood serotonin concentrations in phobic dogs fed a dissociated carbohydrate-based diet: A pilot study. Dog Behav. 2019, 2, 9–17. [Google Scholar]
- Lloyd, J.K.F. Minimising Stress for Patients in the Veterinary Hospital: Why It Is Important and What Can Be Done about it. Vet. Sci. 2017, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, P.T.; Hazel, S.J.; Browne, M.; Serpell, J.A.; Mcarthur, L.; Smith, B.P. Investigating risk factors that predict a dog’ s fear during veterinary consultations. PLoS ONE 2019, 14, e0215416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doring, D.; Roscher, A.; Scheipl, F.; Ku, H.; Erhard, M.H. Fear-related behaviour of dogs in veterinary practice. Vet. J. 2009, 182, 38–43. [Google Scholar] [CrossRef]
- Mills, D.S.; Ramos, D.; Gandia Estelles, M.; Hargrave, C. A triple blind placebo-controlled investigation into the assessment of the effect of Dog Appeasing Pheromone (DAP) on anxiety related behaviour of problem dogs in the veterinary clinic. Appl. Anim. Behav. Sci. 2006, 98, 114–126. [Google Scholar] [CrossRef]
- Bearcroft, C.Y.; Farthing, M.J.G.; Perretts, D. Determination of 5=Hydroxytryptamine, in Plasma and Urine by HPLC with 5-Hydroxyindoleacetic Acid and Tryptophan Fluorimetric Detection. Biomed. Chromatogr. 1995, 9, 23–27. [Google Scholar] [CrossRef]
- Atkinson, W.; Lockhart, S.J.; Houghton, L.A.; Keevil, B.G. Validation of the measurement of low concentrations of 5-hydroxytryptamine in plasma using high performance liquid chromatography. J. Chromatogr. B 2006, 832, 173–176. [Google Scholar] [CrossRef]
- Yamamoto, T.; Castell, L.M.; Botella, J.; Powell, H.; Hall, G.M.; Young, A.; Newsholme, E.A.; Medical, H.; Nw, L.; Hospital, F.; et al. Changes in the Albumin Binding of Tryptophan During Postoperative Recovery: A Possible Link With Central Fatigue? Brain Res. Bull. 1997, 43, 43–46. [Google Scholar] [CrossRef]
- Badawy, A.A. Modulation of Tryptophan and Serotonin Metabolism as a Biochemical Basis of the Behavioral Effects of Use and Withdrawal of Androgenic-Anabolic Steroids and Other Image- and Performance-Enhancing Agents. Int. J. Tryptophan Res. 2018, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badawy, A. Plasmafree tryptophan revisited: What you need to know and do before measuring it. J. Psychopharmacol. 2008, 24, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Dodman, N.H.; Reisner, I.R.; Shuster, L.; Rand, W.; Luescher, U.A.; Robinson, I.; Houpt, K.A. Effect of dietary protein content on behavior in dogs. J. Am. Vet. Med. Assoc. 1996, 208, 376–379. [Google Scholar] [PubMed]
- Audhya, T.; Adams, J.B.; Johansen, L. Correlation of serotonin levels in CSF, platelets, plasma, and urine. Biochim. Biophys. Acta 2012, 1820, 1496–1501. [Google Scholar] [CrossRef] [PubMed]
- Sarrias, M.J.; Cab, P.; Martinez, E.; Artigas, F. Relationship Between Serotoninergic Measures in Blood and Cerebrospinal Fluid Simultaneously Obtained in Humans. J. Neurochem. 1990, 53, 783–786. [Google Scholar] [CrossRef]
- Liu, M.; Xiang, J.; Zhou, J.; Ding, H. A disposable amperometric sensor for rapid detection of serotonin in the blood and brain of the depressed mice based on Nafion membrane-coated colloidal gold screen-printed electrode. J. Electroanal. Chem. 2010, 640, 1–7. [Google Scholar] [CrossRef]
- Nakatani, Y.; Sato-suzuki, I.; Tsujino, N.; Nakasato, A.; Seki, Y.; Fumoto, M.; Arita, H. Augmented brain 5-HT crosses the blood—brain barrier through the 5-HT transporter in rat. Eur. J. Neurosci. 2008, 27, 2466–2472. [Google Scholar] [CrossRef]
- Brust, P.; Friedrich, A.; Krizbai, I.A.; Bergmann, R.; Roux, F.; Ganapathy, V.; Johannsen, B. Functional Expression of the Serotonin Transporter in Immortalized Rat Brain Microvessel Endothelial Cells. J. Neurochem. 2000, 74, 1241–1248. [Google Scholar] [CrossRef]
- Wakayama, K.; Ohtsuki, S.; Takanaga, H.; Hosoya, K.; Terasaki, T. Localization of norepinephrine and serotonin transporter in mouse brain capillary endothelial cells. Neurosci. Res. 2002, 44, 173–180. [Google Scholar] [CrossRef]
- Young, L.W.; Darios, E.S.; Watts, S.W. An immunohistochemical analysis of SERT in the blood—brain barrier of the male rat brain. Histochem. Cell Biol. 2015, 144, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Young, S.N.; Leyton, M. The role of serotonin in human mood and social interaction Insight from altered tryptophan levels. Pharmacol. Biochem. Behav. 2002, 71, 857–865. [Google Scholar] [CrossRef]
- Walz, J.C.; Stertz, L.; Fijtman, A.; dos Santos, B.T.M.Q.; de Almeida, R.M. Tryptophan diet reduces aggressive behavior in male mice. Psychol. Neurosci. 2013, 6, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Rayment, D.J.; Peters, R.A.; Marston, L.C.; De Groef, B. Relationships between serum serotonin, plasma cortisol, and behavioral factors in a mixed-breed, -sex, and -age group of pet dogs. J. Vet. Behav. Clin. Appl. Res. 2020, 38, 96–102. [Google Scholar] [CrossRef]
- Young, S.N. The effect of raising and lowering tryptophan levels on human mood and social behaviour. Philos. Trans. R. Soc. B 2013, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, S.E.; Kristensen, A.; Reimann, M.J.; Eriksen, N.B.; Petersen, S.F.; Marschner, C.B.; Tarnow, I.; Oyama, M.A.; Olsen, L.H. Plasma and serum serotonin concentrations and surface-bound platelet serotonin expression in Cavalier King Charles Spaniels with myxomatous mitral valve disease. Am. J. Vet. Res. 2015, 76, 12–14. [Google Scholar] [CrossRef]
- Alberghina, D.; Tropia, E.; Piccione, G.; Giannetto, C.; Panzera, M. Serum serotonin (5-HT) in dogs (Canis familiaris): Preanalytical factors and analytical procedure for use of reference values in behavioral medicine. J. Vet. Behav. Clin. Appl. Res. 2019, 32, 72–75. [Google Scholar] [CrossRef]
- Arndt, J.W.; Reynolds, C.A.; Singletary, G.E.; Connolly, J.M.; Levy, R.J.; Oyama, M.A. Serum Serotonin Concentrations in Dogs with Degenerative Mitral Valve Disease. J. Vet. Intern. Med. 2009, 23, 1208–1213. [Google Scholar] [CrossRef]
- Höglund, K.; Häggström, J.; Hanas, S.; Merveille, A.; Gouni, V.; Wiberg, M.; Lundgren Willesen, J.; Mc Entee, K.M.; Mejer Sørensen, L.; Tiret, L.; et al. Interbreed variation in serum serotonin (5-hydroxytryptamine) concentration in healthy dogs. J. Vet. Cardiol. 2018, 20, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.; Yu, J.H.; Jeong, E.; Yoo, H.J.; Kim, M. Meal-related oscillations in the serum serotonin levels in healthy young men. Clin. Endocrinol. 2018, 88, 549–555. [Google Scholar] [CrossRef]
- Alberghina, D.; Piccione, G.; Pumilia, G.; Gioè, M.; Rizzo, M.; Raffo, P.; Panzera, M. Daily fluctuation of urine serotonin and cortisol in healthy shelter dogs and influence of intraspecific social exposure. Physiol. Behav. 2019, 206, 1–6. [Google Scholar] [CrossRef]
- Piccione, G.; Assenza, A.; Fazio, F.; Percipalle, M.; Caola, G. Central fatigue and nycthemeral change of serum tryptophan and serotonin in the athletic horse. J. Circadian Rhythm. 2005, 3, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csoltova, E.; Martineau, M.; Boissy, A.; Gilbert, C. Behavioral and physiological reactions in dogs to a veterinary examination: Owner-dog interactions improve canine well-being. Physiol. Behav. 2017, 177, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Mariti, C.; Pierantoni, L.; Sighieri, C.; Gazzano, A. Guardians’ Perceptions of Dogs’ Welfare and Behaviors Related to Visiting the Veterinary Clinic. J. Appl. Anim. Welf. Sci. 2017, 20, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Voigt, J.; Sekeris, C.E. Induction of tryptophan oxygenase and tyrosine aminotransferase by metabolites of hydrocortisone. Biochim. Biophys. Acta 1980, 633, 422–435. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riggio, G.; Mariti, C.; Sergi, V.; Diverio, S.; Gazzano, A. Serotonin and Tryptophan Serum Concentrations in Shelter Dogs Showing Different Behavioural Responses to a Potentially Stressful Procedure. Vet. Sci. 2021, 8, 1. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8010001
Riggio G, Mariti C, Sergi V, Diverio S, Gazzano A. Serotonin and Tryptophan Serum Concentrations in Shelter Dogs Showing Different Behavioural Responses to a Potentially Stressful Procedure. Veterinary Sciences. 2021; 8(1):1. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8010001
Chicago/Turabian StyleRiggio, Giacomo, Chiara Mariti, Valeria Sergi, Silvana Diverio, and Angelo Gazzano. 2021. "Serotonin and Tryptophan Serum Concentrations in Shelter Dogs Showing Different Behavioural Responses to a Potentially Stressful Procedure" Veterinary Sciences 8, no. 1: 1. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8010001