
Sirtuin 3 Restores Synthesis and Secretion of Very Low-Density Lipoproteins in Cow Hepatocytes Challenged with Nonesterified Fatty Acids In Vitro


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Construction of Recombinant Adenoviruses and siRNA
2.2. Cow Primary Hepatocytes Culture and Treatment
2.3. Quantitative Reverse-Transcription PCR (qRT-PCR) Assay
2.4. Measurement of Very Low-Density Lipoprotein Content
2.5. Statistical Analysis
3. Results
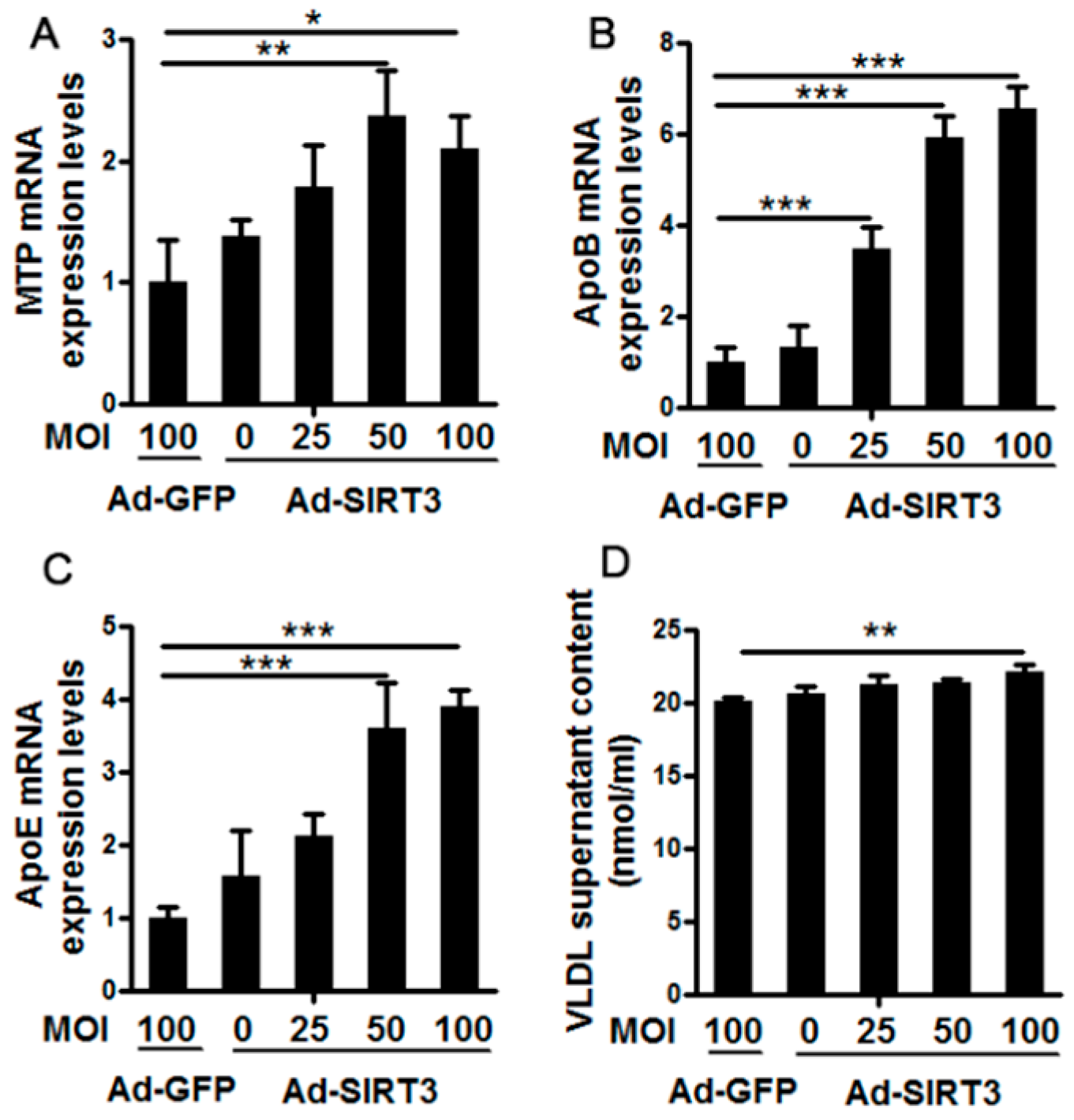
3.1. Ad-SIRT3 Administration Promoted Very Low-Density LipoproteinSynthesis and Assembly in Cow Hepatocytes
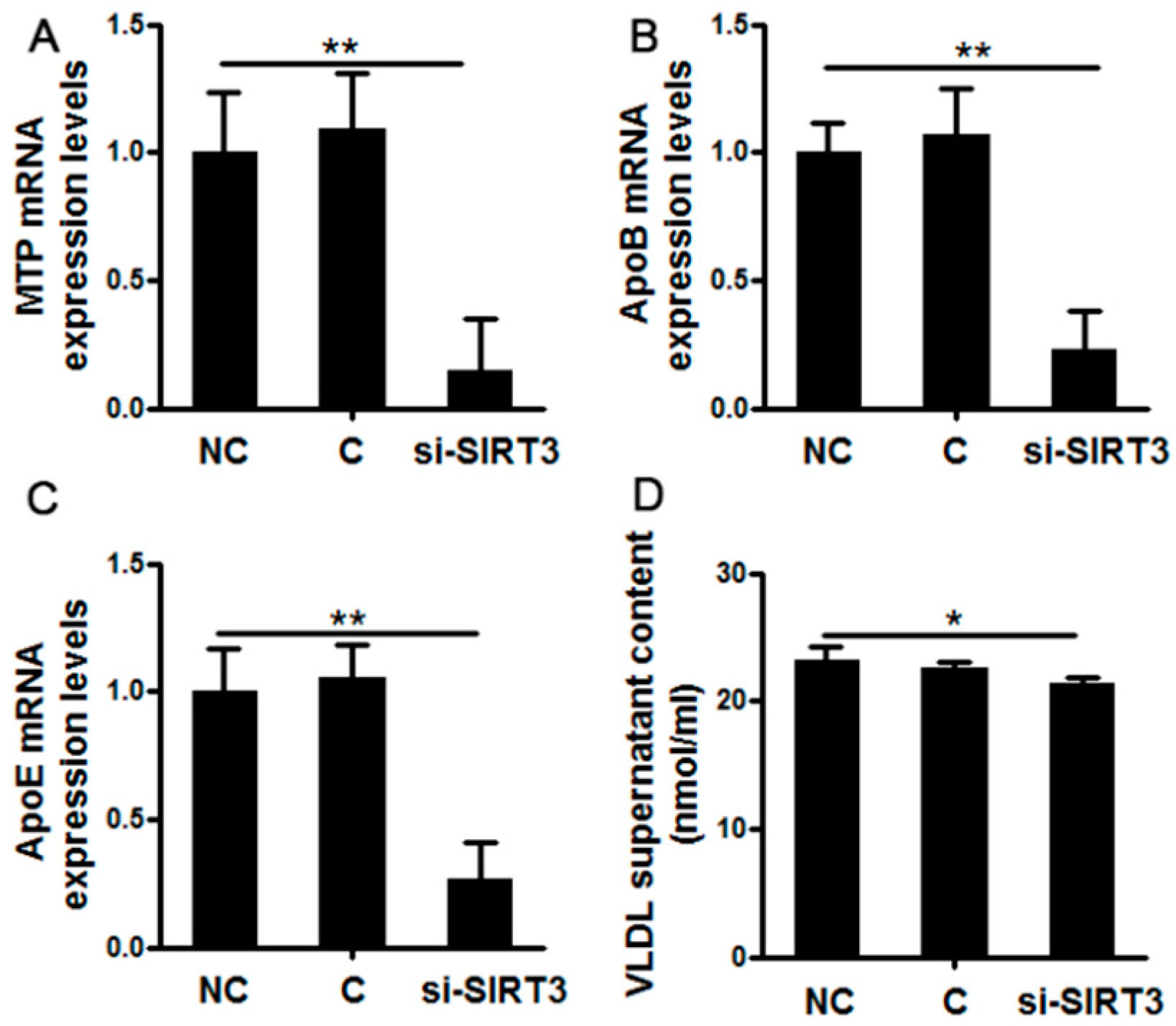
3.2. SIRT3 Silencing Decreased Very Low-Density Lipoprotein Synthesis and Assembly in Cow Hepatocytes
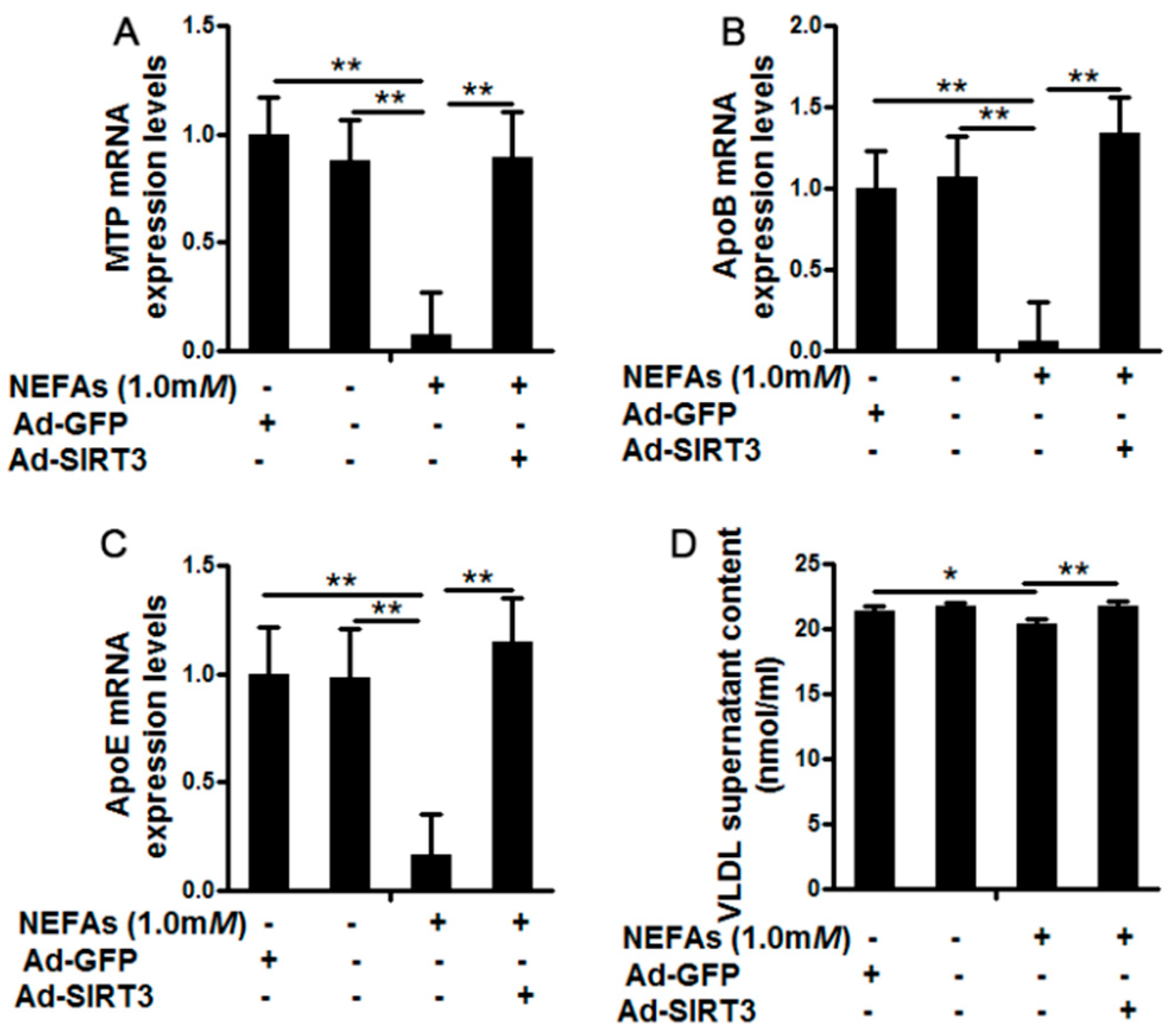
3.3. SIRT3 Reversed the Inhibition of Very Low-Density Lipoprotein Synthesis and Assembly by High Nonesterified Fatty Acids in Cow Hepatocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Herdt, T.H. Fatty Liver in Dairy Cows. Veter. Clin. N. Am. Food Anim. Pract. 1988, 4, 269–287. [Google Scholar] [CrossRef]
- Katoh, N. Relevance of Apolipoproteins in the Development of Fatty Liver and Fatty Liver-Releted Peripartum Diseases in Dairy Cows. J. Veter. Med. Sci. 2002, 64, 293–307. [Google Scholar] [CrossRef] [Green Version]
- Bobe, G.; Young, J.; Beitz, D. Invited Review: Pathology, Etiology, Prevention, and Treatment of Fatty Liver in Dairy Cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef] [Green Version]
- Evink, T.; Endres, M. Management, operational, animal health, and economic characteristics of large dairy herds in 4 states in the Upper Midwest of the United States. J. Dairy Sci. 2017, 100, 9466–9475. [Google Scholar] [CrossRef]
- Bremmer, D.; Trower, S.; Bertics, S.; Besong, S.; Bernabucci, U.; Grummer, R. Etiology of Fatty Liver in Dairy Cattle: Effects of Nutritional and Hormonal Status on Hepatic Microsomal Triglyceride Transfer Protein. J. Dairy Sci. 2000, 83, 2239–2251. [Google Scholar] [CrossRef]
- Gruffat-Mouty, D.; Graulet, B.; Durand, D.; Samson-Bouma, M.-E.; Bauchart, D. Apolipoprotein B Production and Very Low Density Lipoprotein Secretion by Calf Liver Slices. J. Biochem. 1999, 126, 188–193. [Google Scholar] [CrossRef]
- E Brumby, P.; Anderson, M.; Tuckley, B.; E Storry, J.; Hibbit, K.G. Lipid metabolism in the cow during starvation-induced ketosis. Biochem. J. 1975, 146, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Durand, D.; Martinaud, M.; Gruffat, D.; Leplaix-Charlat, L.; Lefaivre, J.; Ollier, A.; Robert, J.; Chilliard, Y.; Bauchart, D. Plasma and hepatic lipids and lipoproteins in the underfed high-yielding dairy cow during early lactation. Ann. Zootech. 1994, 43, 46–47. [Google Scholar] [CrossRef]
- Marcos, E.; Mazur, A.; Cardot, P.; Rayssiguier, Y. Serum apolipoproteins B and A-I and naturally occurring fatty liver in dairy cows. Lipids 1990, 25, 575–577. [Google Scholar] [CrossRef]
- Liu, L.; Li, X.; Li, Y.; Guan, Y.; Song, Y.; Yin, L.; Chen, H.; Lei, L.; Liu, J.; Li, X.; et al. Effects of nonesterified fatty acids on the synthesis and assembly of very low density lipoprotein in bovine hepatocytes in vitro. J. Dairy Sci. 2014, 97, 1328–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xiang, H.; Liu, J.; Chen, Y.; He, R.-R.; Liu, B. Mitochondrial Sirtuin 3: New emerging biological function and therapeutic target. Theranostics 2020, 10, 8315–8342. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yao, L.; Peng, T.; Wen, L.; Cai, W.; Jia, X.; He, J. Hepatic Sirt3 expression declines postpartum in dairy goats. J. Dairy Res. 2018, 85, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xing, D.; Du, X.; Peng, T.; McFadden, J.W.; Wen, L.; Lei, H.; Dong, W.; Liu, G.; Wang, Z.; et al. Sirtuin 3 improves fatty acid metabolism in response to high nonesterified fatty acids in calf hepatocytes by modulating gene expression. J. Dairy Sci. 2020, 103, 6557–6568. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.E.; Gaughan, E.M. Partial Hepatic Resection for Treatment of a Single Liver Abscess in a Dairy Heifer. Veter. Surg. 1988, 17, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Bernabucci, U.; Ronchi, B.; Basiricò, L.; Pirazzi, D.; Rueca, F.; Lacetera, N.; Nardone, A. Abundance of mRNA of Apolipoprotein B100, Apolipoprotein E, and Microsomal Triglyceride Transfer Protein in Liver from Periparturient Dairy Cows. J. Dairy Sci. 2004, 87, 2881–2888. [Google Scholar] [CrossRef] [Green Version]
- Ooi, E.; Janus, E.D.; Grant, S.J.; Sinclair, L.M.; Barrett, P.H. Effect of apolipoprotein E genotype on apolipoprotein B-100 metabolism in normolipidemic and hyperlipidemic subjects. J. Lipid Res. 2010, 51, 2413–2421. [Google Scholar] [CrossRef] [Green Version]
- Herdt, T.H.; Liesman, J.S.; Gerloff, B.J.; Emery, R.S. Reduction of serum triacylglycerol-rich lipoprotein concentrations in cows with hepatic lipidosis. Am. J. Veter. Res. 1983, 44, 293–296. [Google Scholar]
- Loor, J.J.; Everts, R.E.; Bionaz, M.; Dann, H.M.; Morin, D.E.; Oliveira, R.; Rodriguez-Zas, S.L.; Drackley, J.K.; Lewin, H.A. Nutrition-induced ketosis alters metabolic and signaling gene networks in liver of periparturient dairy cows. Physiol. Genom. 2007, 32, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Li, X.; Liu, G.; Loor, J.; Bucktrout, R.; Sun, X.; Li, G.; Shu, X.; Dong, J.; Wang, Y.; et al. Perilipin 5 promotes hepatic steatosis in dairy cows through increasing lipid synthesis and decreasing very low density lipoprotein assembly. J. Dairy Sci. 2019, 102, 833–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikawa, S.; Katoh, N. Decreases in serum apolipoprotein B-100 and A-I concentrations in cows with milk fever and downer cows. Can. J. Veter Res. Rev. Can. Rech. Veter. 2002, 66, 31–34. [Google Scholar]
- Mazur, A.; Ayrault-Jarrier, M.; Chilliard, Y.; Rayssiguier, Y. Lipoprotein metabolism in fatty liver dairy cows. Diabete Metab. 1992, 18, 145–149. [Google Scholar]
- Wetterau, J.R.; Lin, M.C.; Jamil, H. Microsomal triglyceride transfer protein. Biochim. Biophys. Acta BBA Lipids Lipid Metab. 1997, 1345, 136–150. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.M.; Shi, J.; Dreizen, P. Microsomal triglyceride transfer protein and its role in apoB-lipoprotein assembly. J. Lipid Res. 2003, 44, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Kalaitzakis, E.; Panousis, N.; Roubies, N.; Giadinis, N.; Kaldrymidou, E.; Georgiadis, M.; Karatzias, H. Clinicopathological evaluation of downer dairy cows with fatty liver. Can. Veter. J. Rev. Veter. Can. 2010, 51, 615–622. [Google Scholar]
- Zhao, B.; Luo, C.; Zhang, M.; Xing, F.; Luo, S.; Fu, S.; Sun, X. Knockdown of phosphatase and tensin homolog (PTEN) inhibits fatty acid oxidation and reduces very low density lipoprotein assembly and secretion in calf hepatocytes. J. Dairy Sci. 2020, 103, 10728–10741. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xin, T.; Li, D.; Wang, C.; Zhu, H.; Zhou, H. Therapeutic effect of Sirtuin 3 on ameliorating nonalcoholic fatty liver disease: The role of the ERK-CREB pathway and Bnip3-mediated mitophagy. Redox Biol. 2018, 18, 229–243. [Google Scholar] [CrossRef]
- Sun, R.; Kang, X.; Zhao, Y.; Wang, Z.; Wang, R.; Fu, R.; Li, Y.; Hu, Y.; Wang, Z.; Shan, W.; et al. Sirtuin 3-mediated deacetylation of acyl- CoA synthetase family member 3 by protocatechuic acid attenuates non-alcoholic fatty liver disease. Br. J. Pharmacol. 2020, 177, 4166–4180. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Yang, J.; Hu, O.; Huang, J.; Ran, L.; Chen, M.; Zhang, Y.; Zhou, X.; Zhu, J.; Zhang, Q.; et al. Dihydromyricetin Ameliorates Nonalcoholic Fatty Liver Disease by Improving Mitochondrial Respiratory Capacity and Redox Homeostasis Through Modulation of SIRT3 Signaling. Antioxid. Redox Signal. 2019, 30, 163–183. [Google Scholar] [CrossRef]
- Xu, X.; Zhu, X.-P.; Bai, J.-Y.; Xia, P.; Li, Y.; Lu, Y.; Li, X.-Y.; Gao, X. Berberine alleviates nonalcoholic fatty liver induced by a high-fat diet in mice by activating SIRT3. FASEB J. 2019, 33, 7289–7300. [Google Scholar] [CrossRef]
- Xu, S.; Gao, Y.; Zhang, Q.; Wei, S.; Chen, Z.; Dai, X.; Zeng, Z.; Zhao, K.-S. SIRT1/3 Activation by Resveratrol Attenuates Acute Kidney Injury in a Septic Rat Model. Oxidative Med. Cell. Longev. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Park, W.; Jin, J.; Kim, W.; Park, S.K.; Kang, K.P. Sirtuin 3 Activation by Honokiol Decreases Unilateral Ureteral Obstruction-Induced Renal Inflammation and Fibrosis via Regulation of Mitochondrial Dynamics and the Renal NF-κB-TGF-β1/Smad Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Chen, L.; Lu, Y.; Lei, S.; Peng, M.; Xia, Z. SIRT3 activator honokiol ameliorates surgery/anesthesia-induced cognitive decline in mice through anti-oxidative stress and anti-inflammatory in hippocampus. CNS Neurosci. Ther. 2018, 25, 355–366. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, D.; Wang, B.; Lu, H.; Peng, T.; Su, J.; Lei, H.; He, J.; Zhou, Y.; Liu, L. Sirtuin 3 Restores Synthesis and Secretion of Very Low-Density Lipoproteins in Cow Hepatocytes Challenged with Nonesterified Fatty Acids In Vitro. Vet. Sci. 2021, 8, 121. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8070121
Xing D, Wang B, Lu H, Peng T, Su J, Lei H, He J, Zhou Y, Liu L. Sirtuin 3 Restores Synthesis and Secretion of Very Low-Density Lipoproteins in Cow Hepatocytes Challenged with Nonesterified Fatty Acids In Vitro. Veterinary Sciences. 2021; 8(7):121. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8070121
Chicago/Turabian StyleXing, Dongmei, Baogen Wang, Hong Lu, Tao Peng, Jianming Su, Hongyu Lei, Jianhua He, Yingfang Zhou, and Lei Liu. 2021. "Sirtuin 3 Restores Synthesis and Secretion of Very Low-Density Lipoproteins in Cow Hepatocytes Challenged with Nonesterified Fatty Acids In Vitro" Veterinary Sciences 8, no. 7: 121. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8070121