ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs
by
 , ,
, ,
Stepan N. Belyakin
1,2,* ,
,
Daniil A. Maksimov
1,
Maria A. Pobedintseva
1,
Petr P. Laktionov
1,3 and
Dinara Voronova
4 1
Genomics Laboratory, Institute of Molecular and Cellular Biology, 630090 Novosibirsk, Russia
2
VetGenomics Laboratory, 630090 Novosibirsk, Russia
3
Laboratory of Epigenetics, Novosibirsk State University, 630090 Novosibirsk, Russia
4
Russian Nihonken Hozonkai (NIPPO) Club, 129626 Moscow, Russia
*
Author to whom correspondence should be addressed.
Vet. Sci. 2022, 9(5), 222; https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050222
Submission received: 23 February 2022
/
Revised: 26 April 2022
/
Accepted: 28 April 2022
/
Published: 3 May 2022
(This article belongs to the Section Veterinary Biomedical Sciences)
Abstract
:Animals exhibit a wide variety of genetically determined coat colors and pigmentation patterns that serve important roles in adaptation and communication. Although the genetics of the main coat colors in dogs have been studied extensively, there are types of coat pigmentation that have not been explained yet. Recently, an association between the variants in the ASIP gene Ventral (VP) and Hair Cycle (HCP) promoters with different coat colors in dogs has been established. Here, we used the new findings as a basis to investigate the genetics of the red sesame coat color in Shiba Inu dogs. Our study revealed that red sesame dogs carry a specific heterozygous ASIP promoter diplotype, VP2-HCP1/VP2-HCP3, where VP2-HCP1 is responsible for the red coat with a dark overlay, and VP2-HCP3 for a tan point-like pattern. This finding explains the inheritance of this coat color pattern and can be used by breeders to produce dogs with this rare phenotype. A comparison of sesame dogs (VP2-HCP1/VP2-HCP3) to a dog homozygous for the VP2-HCP1 promoter haplotype suggests that the incomplete dominance between the ASIP alleles may be involved in the sesame coat formation. These results are in good agreement with the new model explaining how different levels of ASIP gene expression affect the regulation of pigment synthesis in melanocytes.
1. Introduction
The genetics of coat color in domestic dogs is an example of an extensively characterized biological system with peculiar genetic interactions including several epistases [1,2]. The diversity of the coat colors in dogs results from different combinations of two pigments—dark eumelanin and yellow/red pheomelanin—that are synthesized in melanocytes and loaded into the growing hair [3,4,5].
The Agouti Signaling Protein produced by the ASIP gene switches the type of pigment that is produced in melanocytes and thereby plays a central role in coat color formation in different animals [6,7,8]. The ASIP protein is a ligand of the Melanocortin 1 Receptor (MC1R) on the surface of melanocytes. In the absence of ASIP, MC1R promotes eumelanin synthesis. The binding of ASIP to MC1R switches melanocyte to pheomelanin production [9,10,11].
In many animal species, the concentration of ASIP oscillates in the skin on the dorsal side of the body, thereby resulting in a banded pattern of pigmentation along the hair [1,12]. For example, at the starting phase of hair growth, the concentration of ASIP in the skin is low, and eumelanin is predominantly produced in melanocytes and loaded into the hair tip. At some point, the ASIP concentration increases and melanocytes are switched to pheomelanin synthesis, resulting in the deposition of red pigment as the hair continues to grow. When the amount of ASIP reduces again, a new dark band may appear in the hair.
In domestic dogs, the alleles of the ASIP gene are known to produce a number of different patterns. When other genes participating in coat color formation are intact, the dominant allele Ay leads to a solid red coat pattern. There are variants that produce black-and-tan or saddle tan patterns. The most recessive allele in the set (allele a) is responsible for the solid black coat color [13]. The wild-type allele aw forms the wolf-sable coat color, characterized by a pheomelanistic ventrum and a banded pattern of hair pigmentation on the dorsum due to ASIP oscillation in the skin.
The different alleles of the ASIP gene were previously associated with particular DNA polymorphisms or variants (Table 1). These variants are routinely used in many commercial testing laboratories as markers of the corresponding ASIP alleles that explain most of the ASIP gene effects on coat pigmentation in dogs. However, according to a recent report [14], these variants are not causative; they are rather linked to the corresponding alleles and, therefore, important exceptions exist that cannot be comprehensively explained by the old tests interrogating the imperfectly associated variants.
Even prior to the discovery of the different ASIP promoter variants [14], it was already noted that the widely used commercial tests for the associated variants do not always yield the expected genotypes [19]. The new framework of ASIP promoter variants provides a plausible and consistent explanation for the discrepancies observed in the Dreger et al. study [14].
A recent study determined that, apart from the a allele, which is indeed the missense loss-of-function mutation (chr24:23393552C>T [20]), other alleles are associated with different promoter haplotypes of the ASIP gene [14]. Two ASIP gene promoters are known to operate in many animals [12]. The Ventral Promoter (VP) predominantly activates the ASIP gene on the ventral part of the body. Its activity gradually decreases toward the dorsal surface. The transcription from VP is permanent and, therefore, the coat color on the ventral surface of the body is normally red due to pheomelanin deposition. Another promoter, which was called Hair Cycle Promoter (HCP), regulates ASIP gene activity mostly on the dorsal surface of the body. The activity of this promoter normally circulates, leading to a banded hair pattern [1,12]. It was suggested that these promoters affect ASIP gene activity differentially, thereby mediating the phenotypes caused by its different alleles [14].
As has been shown recently, there are at least two different alleles of VP and five alleles of HCP. The combinations of these functional haplotypes accurately determine the different alleles of the ASIP gene [14], as summarized in Table 1. Remarkably, the black-and-tan coat pattern in rabbits has been associated with a deletion in the HCP of the ASIP gene [21], implying that this deletion causes an effect similar to the at allele in dogs.
The hierarchy of the ASIP gene alleles can be presented as follows: Ay > Ays > aw > asa = at > a. Two alleles—asa and at—are semidominant: the dogs with the asa/at genotype show an intermediate coat pattern between black and tan and saddle tan. The black pigmentation on their heads forms a characteristic “widow’s peak”-shaped pattern [17].
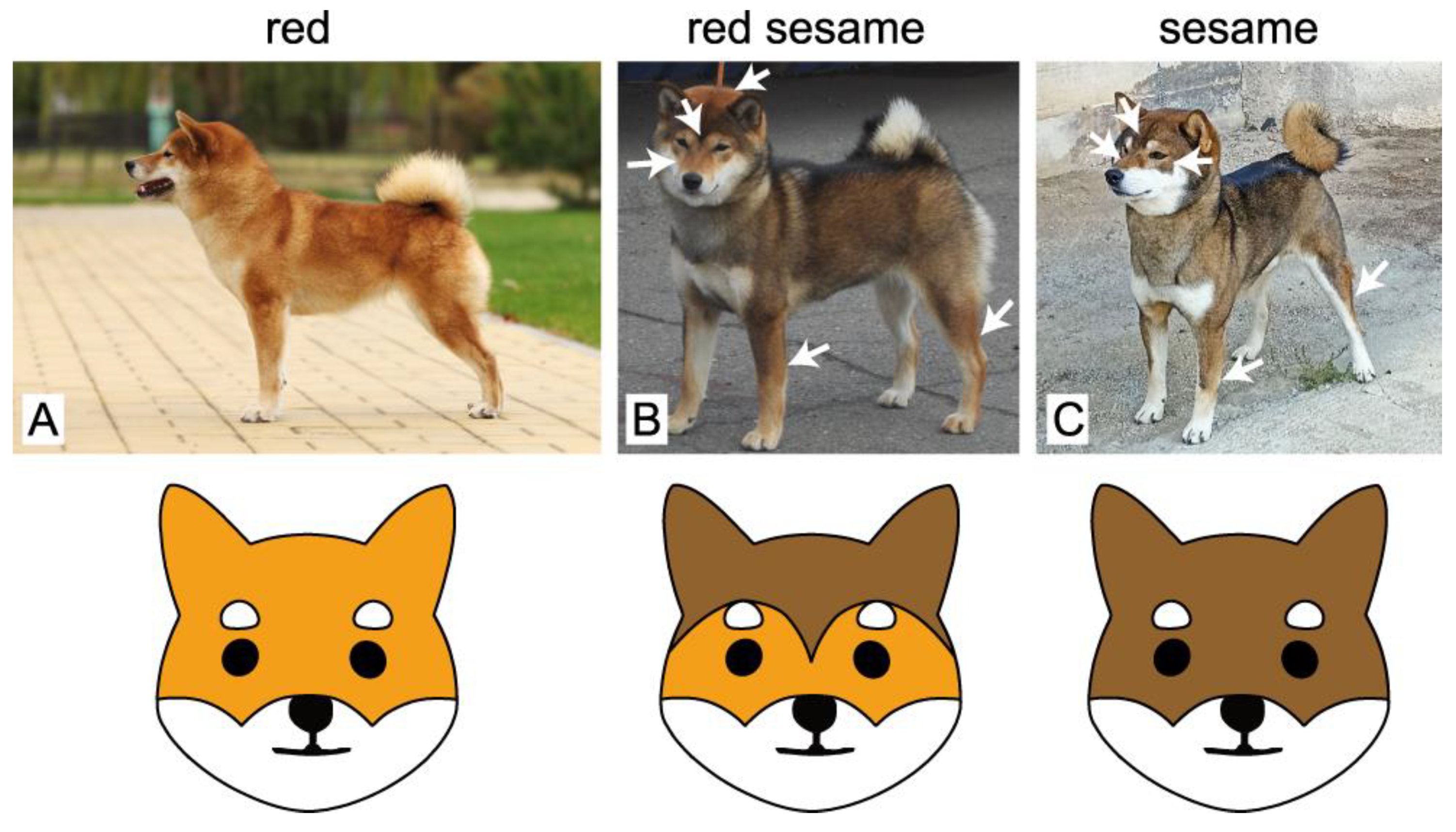
In Shiba Inu dogs, there is a specific coat pattern that is referred to as sesame (Figure 1). This pattern resembles the black and tan, with the solid black areas replaced by the black overlay. According to the FCI breed standard, there are three distinct subtypes of this coat pattern that are classified as sesame, black sesame, or red sesame. These subtypes are primarily differentiated by the amount of black shading, with red sesame demonstrating considerably more red than sesame or black sesame (Figure 1).
Most of the dogs in the Shiba Inu breed are red; a low percentage of them are black and tan (also see the statistics in [22]). The sesame coat colors are the rarest ones. The sesame coat pattern in Shiba Inu dogs is an amazing example of the ASIP gene’s effects on coat pigmentation. It is known that the sesame coat color is determined by the wild-type allele of the ASIP gene, which is very rare in the Shiba Inu breed. In turn, red sesame dogs were shown to possess the double mutation (chr24:23393510G>T and chr24:23393514G>A), as well as the SINE insertion at chr24:23365297, as detected by a widely used genetic test for ASIP alleles (see “Associated genetic variant” column in Table 1). The problem is that the dogs with this genotype can be red sesame or red with hardly any black shading (Figure 2). The reason for this variation and the inheritance of the red sesame coat color has remained elusive.
The inheritance of the sesame coat color is unclear and has puzzled breeders and dog owners for decades. Here, we tested whether the promoter-based model of ASIP alleles can be applied to explain the sesame coat color formation in Shiba Inu.
2. Materials and Methods
2.1. Samples
Shiba Inu buccal swabs were used as a source of genomic DNA for this study. The samples were sent to the VetGenomics laboratory by the owners for genetic testing. The dogs used in this study were initially selected using the coat color information indicated in the Shiba Inu pedigree database [23]. Red and sesame dogs were initially selected. The owners of all the dogs in this study signed an agreement for their participation in the study and sent photographs of their dogs to confirm the coat color. Based on this information, the phenotypes of sesame dogs were classified as sesame and red sesame. The resulting cohort included 57 dogs (42 red, 11 red sesame, 3 sesame dogs, and one dog that looked like a red sesame but had a lighter coat). All of these dogs were tested for causative promoter haplotypes [14] and for the associated genetic variants indicated in Table 1.
2.2. Genotyping of ASIP Promoters
Genomic DNA was isolated from the swabs and used in PCR (BioMaster HS-Taq PCR-Color (2×) Mastermix, Biolabmix LLC, Novosibirsk, Russian Federation) with the primers for VP and HCP [14]. The genotypes were determined by assessing the lengths of PCR products in 1% agarose gel in 1 × Tris–Acetate–EDTA buffer. The primer pairs and resulting fragment lengths are summarized in Table 2.
2.3. Sequencing the MC1R Gene
The coding sequence of the MC1R gene was amplified using the following primers: Forward: 5′-cctcaccaggaacatagcac-3′, Reverse: 5′-ctgagcaagacacctgagag-3′. The 1007 bp PCR product was purified by Polyethylene Glycol (PEG) precipitation [24] in order to remove primers. The purified PCR product was used for Sanger sequencing on a 3500 Genetic Analyser (Applied Biosystems, Foster City, CA, USA).
3. Results
We genotyped ASIP promoters in Shiba Inu dogs with red and sesame coat colors using a PCR-based approach to detect the different VP and HCP variants (Materials and Methods).
This analysis provided a clear concordance between the coat colors and genotypes determined using the new findings about ASIP gene promoters (Table 3). All of the dogs having red coats possessed the most dominant Ay allele in their genotypes (in this case, Ay = VP1-HCP1). Genotyping of the red sesame dogs revealed the genotype Ays/at (Ays = VP2-HCP1), which clearly distinguished them from the red Ay/at dogs. The phenotypically darker sesame dogs were confirmed to have the aw/at genotype. In these cases, the aw allele was directly determined by the VP2-HCP2 promoter combination. No dogs with aw/aw or Ays/aw genotypes were found in the present study.
Both VP1-HCP1 (Ay) and VP2-HCP1 (Ays) combinations are linked to the (chr24:23393510T, chr24:23393514A) variant (Table 1). This explains why routine testing using the previously reported SNV markers could not distinguish between clear red and red sesame dogs, as illustrated in Figure 2.
Our findings explain the puzzling inheritance of the red sesame coat color: red sesame pups may be born if one parent bears the Ays allele and another carries the at allele. To illustrate this, we genotyped a family that produced a red sesame dog. Both parents were red. The genotyping of ASIP gene promoters revealed that the sire had the Ay/at genotype (VP1-HCP1/VP2-HCP2) while the dam was Ay/Ays (VP1-HCP1/VP2-HCP1). Their red sesame daughter was confirmed to have the Ays/at genotype (VP2-HCP1/VP2-HCP3; Figure 3).
To check for the presence of mutations in the MC1R gene that could also be involved in the shaded coat color formation, we sequenced this gene in these three dogs. No mutations were detected in the MC1R gene, thereby suggesting that mutations in locus E apparently do not have a major impact on the sesame phenotype formation in Shiba Inu.
The typical red sesame Shiba Inu dogs that were used in this study have the Ays/at genotype. This suggests that an incomplete dominance of the Ays allele over the at allele could exist. Alternatively, the red sesame coat color would be produced by Ays alone and the recessive at allele would not be manifested in the phenotype. In the latter scenario, Ays/at dogs should not be phenotypically different from Ays/Ays animals.
The Ays allele is quite rare in Shiba Inu dogs. However, we identified one dog with the Ays/Ays genotype (Table 2). As shown in Figure 4, this dog has a dark overlay that is similar to a typical red sesame coat color, but this overlay is generally thinner and much more red pigmentation is observed compared to the red sesame Ays/at dogs that were analyzed in our study. This example suggests that the Ays allele is incompletely dominant over the at allele, yet individual variation in allele expressivity cannot be ruled out. A more representative analysis of confirmed Ays/Ays Shiba Inu dogs would help to validate this.
We also performed the complete sequencing of MC1R coding sequences in this Ays/Ays dog and found two synonymous mutations (chr5:63695055C>T and chr5:63695037C>T). These mutations led to AAG > AAA and TCG > TCA codon substitutions, which did not affect the amino acid sequence of the MC1R protein (Supplementary Material). The frequencies of the resulting codons in the dog genome (AAA—0.437, TCA—0.146) were not dramatically different from the frequencies of the ancestral codons (AAG—0.563, TCG—0.059), thereby suggesting that the translation efficiency would not be significantly affected by these replacements. This analysis indicates that the MC1R gene is most probably not involved in the lighter shade of the coat color that is observed in the Ays/Ays dog.
4. Discussion
Here, we used the recently published model of the ASIP gene effects on coat pigmentation in dogs [14] in order to explain the inheritance of the red sesame coat color in Shiba Inu dogs.
Our results uncover a peculiar interplay between ASIP alleles and the coat color phenotypes. All red sesame Shiba Inu dogs in this study were Ays/at heterozygous (VP2-HCP1/VP2-HCP3). Therefore, this coat color appears to be a product of the intricate regulation of the ASIP gene in the skin. Two copies of the VP2 promoter ensure the permanent expression of this gene on the ventral side of the body. However, a single copy of the HCP1 promoter apparently is unable to maintain the high level of ASIP protein, which would be sufficient to mask the manifestation of the non-functional HCP3 promoter on the dorsal side of the body.
This suggestion is further supported by the phenotype of the Ays/Ays dog, which had a coat color that was similar to but clearly distinct from red sesame. Compared to red sesame dogs, this example could be serve as important evidence of the incomplete dominance of the Ays allele over the at allele in Shiba Inu. Although this observation is based on a single Ays/Ays dog and requires a more representative analysis, it seems to be in line with the previous reports of the interactions between different ASIP alleles. Another example of the incomplete dominance of ASIP alleles was described in Welsh Corgi Pembroke: asa/at combination produces a “widow’s peak”-shaped pattern on the dog’s head, which is intermediate between saddle tan and black-and-tan [17]. These results align very well with the new model explaining the action of ASIP alleles via the differential activity of Ventral and Hair Cycle promoters [14].
Our work was focused on the genotyping of ASIP gene promoters. This design cannot completely exclude the involvement of other genes in sesame coat color. For example, a recently characterized eA allele of the MC1R gene showed a somewhat similar pattern of pigmentation in different dog breeds [25]. As this allele is ancient in canine lineage, it is possible that it also exists in Shiba Inu dogs. However, the direct sequencing of MC1R in the red sesame dog detected no mutations within its CDS. Taken together with the perfect correlation between the red sesame coat color and the Ays/at genotype, this suggests that locus E plays no role in the sesame coat color.
Finally, our conclusions support the recently published promoter-centered model of ASIP alleles and provide a useful tool to the cynological community and particularly to Shiba Inu breeders.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/vetsci9050222/s1, Supplementary File contains the sequence of the MC1R from the Ays/Ays Shiba Inu dog. The detected polymorphisms are indicated in bold.
Author Contributions
S.N.B.: Conceptualization, Funding acquisition, Project administration, Writing—original draft; D.A.M.: Data curation, Formal analysis; M.A.P.: Investigation, Methodology; P.P.L.: Investigation, Resources, Formal analysis; D.V.: Resources, Visualization. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Fundamental Scientific Research Project: FWGZ-2021-0017.
Institutional Review Board Statement
The study protocol was approved by the Ethics Committee on Animal and Human Research of the Institute of Molecular and Cellular Biology (Novosibirsk, Russia), Protocol No. 02/21, on 04/08/2021.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: http://www.shiba-pedigree.ru (accessed on 21 October 2021).
Acknowledgments
The authors are grateful to all of the Shiba Inu breeders and owners who agreed to participate in this study anonymously or publicly, and particularly to Elena Gerasimova, Elena Shubnikova, Karine Kaliyan, Evgeniya Makaricheva, Dmitri Mikhailov, Nadezhda Sadovnikova, Julija Tarhanova, and Andrea Fedele. S.N.B., D.A.M. and P.P.L. were partially supported by the Russian Fundamental Scientific Research Project (FWGZ-2021-0017).
Conflicts of Interest
S.N.B. is employed at the VetGenomics Company (Novosibirsk, Russia), which provides commercial genetic tests for domestic animals.
References
- Kaelin, C.B.; Barsh, G.S. Genetics of pigmentation in dogs and cats. Annu. Rev. Anim. Biosci. 2013, 1, 125–156. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Berryere, T.G. Genes affecting coat colour and pattern in domestic dogs: A review. Anim. Genet. 2007, 38, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes are transferred from melanocytes to keratinocytes through the processes of packaging, release, uptake, and dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Tobin, D.J. Human hair pigmentation--biological aspects. Int. J. Cosmet. Sci. 2008, 30, 233–257. [Google Scholar] [CrossRef] [PubMed]
- Prota, G. Recent advances in the chemistry of melanogenesis in mammals. J. Investig. Dermatol. 1980, 75, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Barsh, G.S. Regulation of Pigment Type Switching by Agouti, Melanocortin Signaling, Attractin, and Mahoganoid. In The Pigmentary System; Blackwell Publishing Ltd: Oxford, UK, 2006; pp. 395–409. [Google Scholar]
- Chandramohan, B.; Renieri, C.; La Manna, V.; La Terza, A. The alpaca agouti gene: Genomic locus, transcripts and causative mutations of eumelanic and pheomelanic coat color. Gene 2013, 521, 303–310. [Google Scholar] [CrossRef]
- Han, J.L.; Yang, M.; Yue, Y.J.; Guo, T.T.; Liu, J.B.; Niu, C.E.; Yang, B.H. Analysis of agouti signaling protein (ASIP) gene polymorphisms and association with coat color in Tibetan sheep (Ovis aries). Genet. Mol. Res. 2015, 14, 1200–1209. [Google Scholar] [CrossRef]
- Millar, S.E.; Miller, M.W.; Stevens, M.E.; Barsh, G.S. Expression and transgenic studies of the mouse agouti gene provide insight into the mechanisms by which mammalian coat color patterns are generated. Development 1995, 121, 3223–3232. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Lamoreux, M.L.; Wilson, B.D.; Barsh, G.S. Interaction of Agouti protein with the melanocortin 1 receptor in vitro and in vivo. Genes Dev. 1998, 12, 316–330. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Willard, D.; Patel, I.R.; Kadwell, S.; Overton, L.; Kost, T.; Luther, M.; Chen, W.; Woychik, R.P.; Wilkison, W.O.; et al. Agouti protein is an antagonist of the melanocyte-stimulating-hormone receptor. Nature 1994, 371, 799–802. [Google Scholar] [CrossRef]
- Vrieling, H.; Duhl, D.M.; Millar, S.E.; Miller, K.A.; Barsh, G.S. Differences in dorsal and ventral pigmentation result from regional expression of the mouse agouti gene. Proc. Natl. Acad. Sci. USA 1994, 91, 5667–5671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, M.B. Genetics of the Dog; Howell Book House: New York, NY, USA, 1989. [Google Scholar]
- Bannasch, D.L.; Kaelin, C.B.; Letko, A.; Loechel, R.; Hug, P.; Jagannathan, V.; Henkel, J.; Roosje, P.; Hytonen, M.K.; Lohi, H.; et al. Dog colour patterns explained by modular promoters of ancient canid origin. Nat. Ecol. Evol. 2021, 5, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Berryere, T.G.; Kerns, J.A.; Barsh, G.S.; Schmutz, S.M. Association of an Agouti allele with fawn or sable coat color in domestic dogs. Mamm. Genome 2005, 16, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Dreger, D.L.; Schmutz, S.M. A SINE insertion causes the black-and-tan and saddle tan phenotypes in domestic dogs. J. Hered. 2011, 102 (Suppl. 1), S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Dreger, D.L.; Parker, H.G.; Ostrander, E.A.; Schmutz, S.M. Identification of a mutation that is associated with the saddle tan and black-and-tan phenotypes in Basset Hounds and Pembroke Welsh Corgis. J. Hered. 2013, 104, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., 3rd; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Dreger, D.L.; Anderson, H.; Donner, J.; Clark, J.A.; Dykstra, A.; Hughes, A.M.; Ekenstedt, K.J. Atypical Genotypes for Canine Agouti Signaling Protein Suggest Novel Chromosomal Rearrangement. Genes 2020, 11, 739. [Google Scholar] [CrossRef]
- Kerns, J.A.; Newton, J.; Berryere, T.G.; Rubin, E.M.; Cheng, J.F.; Schmutz, S.M.; Barsh, G.S. Characterization of the dog Agouti gene and a nonagoutimutation in German Shepherd Dogs. Mamm. Genome 2004, 15, 798–808. [Google Scholar] [CrossRef]
- Letko, A.; Ammann, B.; Jagannathan, V.; Henkel, J.; Leuthard, F.; Schelling, C.; Carneiro, M.; Drogemuller, C.; Leeb, T. A deletion spanning the promoter and first exon of the hair cycle-specific ASIP transcript isoform in black and tan rabbits. Anim. Genet. 2020, 51, 137–140. [Google Scholar] [CrossRef]
- Dreger, D.L.; Hooser, B.N.; Hughes, A.M.; Ganesan, B.; Donner, J.; Anderson, H.; Holtvoigt, L.; Ekenstedt, K.J. True Colors: Commercially-acquired morphological genotypes reveal hidden allele variation among dog breeds, informing both trait ancestry and breed potential. PLoS ONE 2019, 14, e0223995. [Google Scholar] [CrossRef] [Green Version]
- Shiba Inu Pedigree DataBase. Available online: http://www.shiba-pedigree.ru (accessed on 21 October 2021).
- Lis, J.T. Fractionation of DNA fragments by polyethylene glycol induced precipitation. Methods Enzymol. 1980, 65, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.; Honkanen, L.; Ruotanen, P.; Mathlin, J.; Donner, J. Comprehensive genetic testing combined with citizen science reveals a recently characterized ancient MC1R mutation associated with partial recessive red phenotypes in dog. Canine Med. Genet. 2020, 7, 16. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Examples of red (A), red sesame (B), and sesame (C) coat colors in Shiba Inu. Compared to sesame, red sesame dogs have considerably more red pigmentation that is specifically distributed along the body, as indicated with white arrows. A characteristic “widow’s peak”-shaped dark pigmentation between the eyes is shown on the red sesame dog (B). Sesame dogs (C) typically demonstrate different facial marks (shown with arrows). The illustrations below schematically show the differences in the facial markings of red sesame and sesame dogs.
Figure 1.
Examples of red (A), red sesame (B), and sesame (C) coat colors in Shiba Inu. Compared to sesame, red sesame dogs have considerably more red pigmentation that is specifically distributed along the body, as indicated with white arrows. A characteristic “widow’s peak”-shaped dark pigmentation between the eyes is shown on the red sesame dog (B). Sesame dogs (C) typically demonstrate different facial marks (shown with arrows). The illustrations below schematically show the differences in the facial markings of red sesame and sesame dogs.

Figure 2.
Two dogs that were tested identical Ay/at with the old tests using the associated markers for Ay (chr24:23393351T, chr24:23393514A) and at (SINE insertion at chr24:23365297). While the two dogs have identical marker genotypes, they show a striking difference in phenotype: the dog in (A) is purely red, while the dog in (B) is red sesame.
Figure 2.
Two dogs that were tested identical Ay/at with the old tests using the associated markers for Ay (chr24:23393351T, chr24:23393514A) and at (SINE insertion at chr24:23365297). While the two dogs have identical marker genotypes, they show a striking difference in phenotype: the dog in (A) is purely red, while the dog in (B) is red sesame.

Figure 3.
Genotyping of the red sesame dog and her red parents. Red parents and their red sesame daughter were PCR-genotyped for ASIP gene promoter haplotypes as described in [14]. The genotyping of the sire revealed the presence of VP1, VP2, HCP1, and HCP3 promoter haplotypes, which uniquely corresponds to the Ay/at genotype as indicated under the gel photograph [14]. The dam possessed VP1, VP2, and HCP1 promoters, which correspond to the Ay/Ays genotype. Their daughter inherited Ays from dam and at from sire, as shown by the presence of VP2, HCP1, and HCP3 variants. The DNA marker sizes are shown in the first photograph.
Figure 3.
Genotyping of the red sesame dog and her red parents. Red parents and their red sesame daughter were PCR-genotyped for ASIP gene promoter haplotypes as described in [14]. The genotyping of the sire revealed the presence of VP1, VP2, HCP1, and HCP3 promoter haplotypes, which uniquely corresponds to the Ay/at genotype as indicated under the gel photograph [14]. The dam possessed VP1, VP2, and HCP1 promoters, which correspond to the Ay/Ays genotype. Their daughter inherited Ays from dam and at from sire, as shown by the presence of VP2, HCP1, and HCP3 variants. The DNA marker sizes are shown in the first photograph.

Figure 4.
Incomplete dominance of the Ays allele over the at allele in red sesame Shiba Inu. Ays/at dog has a typical red sesame coat. The Ays/Ays dog discovered in this study demonstrates significantly more red in their coat color, although with obvious dark shading, clearly distinguishing it from red Shiba Inu dogs. The illustrations below schematically show the differences in the facial markings of Ays/at and Ays/Ays dogs.
Figure 4.
Incomplete dominance of the Ays allele over the at allele in red sesame Shiba Inu. Ays/at dog has a typical red sesame coat. The Ays/Ays dog discovered in this study demonstrates significantly more red in their coat color, although with obvious dark shading, clearly distinguishing it from red Shiba Inu dogs. The illustrations below schematically show the differences in the facial markings of Ays/at and Ays/Ays dogs.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Genetic characteristics of ASIP gene alleles.
| Allele Name | Causative Promoter Haplotype & | Associated Genetic Variant # | Reference § |
|---|---|---|---|
| Dominant yellow (allele Ay) | DY: VP1-HCP1 | chr24:23393510T and chr24:23393514A | [15] |
| Shaded yellow (allele Ays) £ | SY: VP2-HCP1 | chr24:23393510T and chr24:23393514A | [15] |
| Wild type (allele aw) | AG: VP2-HCP2 | NA | |
| Saddle tan (allele asa) | BS: VP1-HCP4 | SINE insertion at chr24:23365297 | [16] |
| Black and tan (allele at) | BB: VP2-HCP3-5 | SINE insertion at chr24:23365297 and chr24:23252755-70dup | [16] [17] |
& Causative promoter haplotypes discovered [14]. Alternative allele abbreviations suggested by [14] state for: DY—Dominant Yellow; SY: Shaded Yellow; AG: Agouti; BS: Black Saddle; BB: Black Back. # Genomic coordinates according to CanFam3.1 assembly [18]. § Literature references for the associations indicated in the column “Associated genetic variant”. £ The Ays (Ay, shaded) name was introduced in this study to distinguish it from the canonical Dominant yellow Ay allele. Both of these alleles are associated with the same genetic variant indicated in the column “Associated genetic variant” but they correspond to different causative promoter haplotypes.
Table 2.
Primers used for VP and HCP variant detection and the resulting fragment lengths.
| Primer Pair * | Product Size (bp) in the ASIP Promoter Haplotypes | ||||||
|---|---|---|---|---|---|---|---|
| VP1 | VP2 | HCP1 | HCP2 | HCP3 | HCP4 | HCP5 | |
| 5′-AGCATGCTTATGTGGCATGT-3′ 5′-CGCTCTTTCAATGTGATTGG-3′ | 475 | 300 | - | - | - | - | - |
| 5′-TTAAAAAGTGAAAGTGAAAAGATAACCC-3′ 5′-CAAAATTCTGGGTGGGCTAA-3′ | - | - | 600 | 400 | 780 | - | - |
| 5′-TTAAAAAGTGAAAGTGAAAAGATAACCC-3′ 5′-TCAATGGAAATGGCAGAACA-3′ | - | - | - | - | - | 860 | - |
| 5′-GATTGAAAGCCAAAGGGTGA-3′ 5′-AGAGCAGGCCAGGTTTTACA-3′ | - | - | 300 | 300 | 300 | 300 | 400 |
* The primer sequences were taken from [14].
Table 3.
Genotyping ASIP gene promoter haplotypes * in 56 red and sesame Shiba Inu dogs.
| Coat Color | Ay/Ay | Ay/Ays | Ay/at | Ay/aw | Ays/at | aw/at |
|---|---|---|---|---|---|---|
| red | 26 | 7 | 6 | 3 | - | - |
| red sesame | - | - | - | - | 11 | - |
| sesame | - | - | - | - | - | 3 |
* The combinations of promoter haplotypes indicated in Table 1 were used as the determinants of ASIP alleles: Ay, Ays, aw, and at.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Belyakin, S.N.; Maksimov, D.A.; Pobedintseva, M.A.; Laktionov, P.P.; Voronova, D. ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs. Vet. Sci. 2022, 9, 222. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050222
AMA Style
Belyakin SN, Maksimov DA, Pobedintseva MA, Laktionov PP, Voronova D. ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs. Veterinary Sciences. 2022; 9(5):222. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050222
Chicago/Turabian StyleBelyakin, Stepan N., Daniil A. Maksimov, Maria A. Pobedintseva, Petr P. Laktionov, and Dinara Voronova. 2022. "ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs" Veterinary Sciences 9, no. 5: 222. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050222
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.