
Biological Compositions of Canine Amniotic Membrane and Its Extracts and the Investigation of Corneal Wound Healing Efficacy In Vitro



Abstract
:1. Introduction
2. Materials and Methods
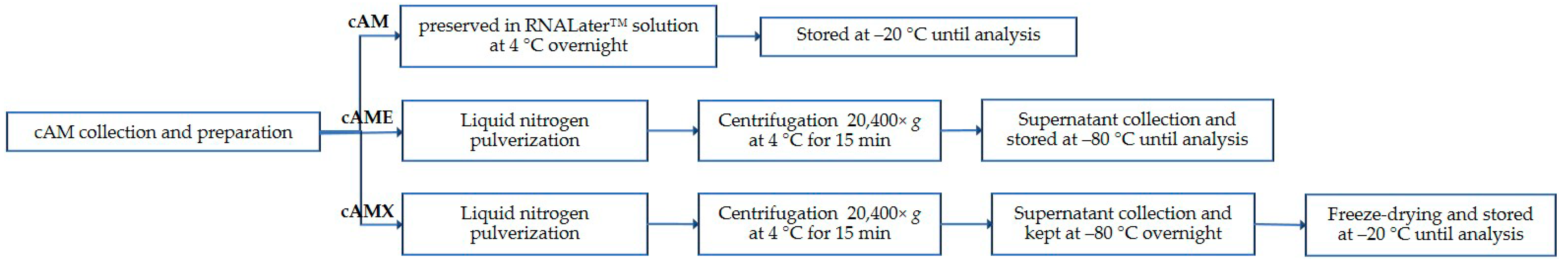
2.1. Canine Amniotic Membrane Collection and Preparation
2.2. Canine Amniotic Membrane Preparation
2.3. Gene Expression Study by Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.3.1. RNA Isolation
2.3.2. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.3.3. Statistical Analysis
2.4. Protein Expression Study by Western Blot Analysis
2.4.1. Protein Extraction
2.4.2. Western Blot Analysis
2.4.3. Statistical Analysis
2.5. Investigation of Corneal Wound Healing Efficacy of cAME and cAMX
2.5.1. Preparation of cAME and cAMX
2.5.2. Human Corneal Epithelial Cell Culture
2.5.3. Cell Viability Assay
2.5.4. Statistical Analysis
2.5.5. Wound Healing Assay
2.5.6. Statistical Analysis
3. Results
3.1. cAME and cAMX Can Be Generated from cAM
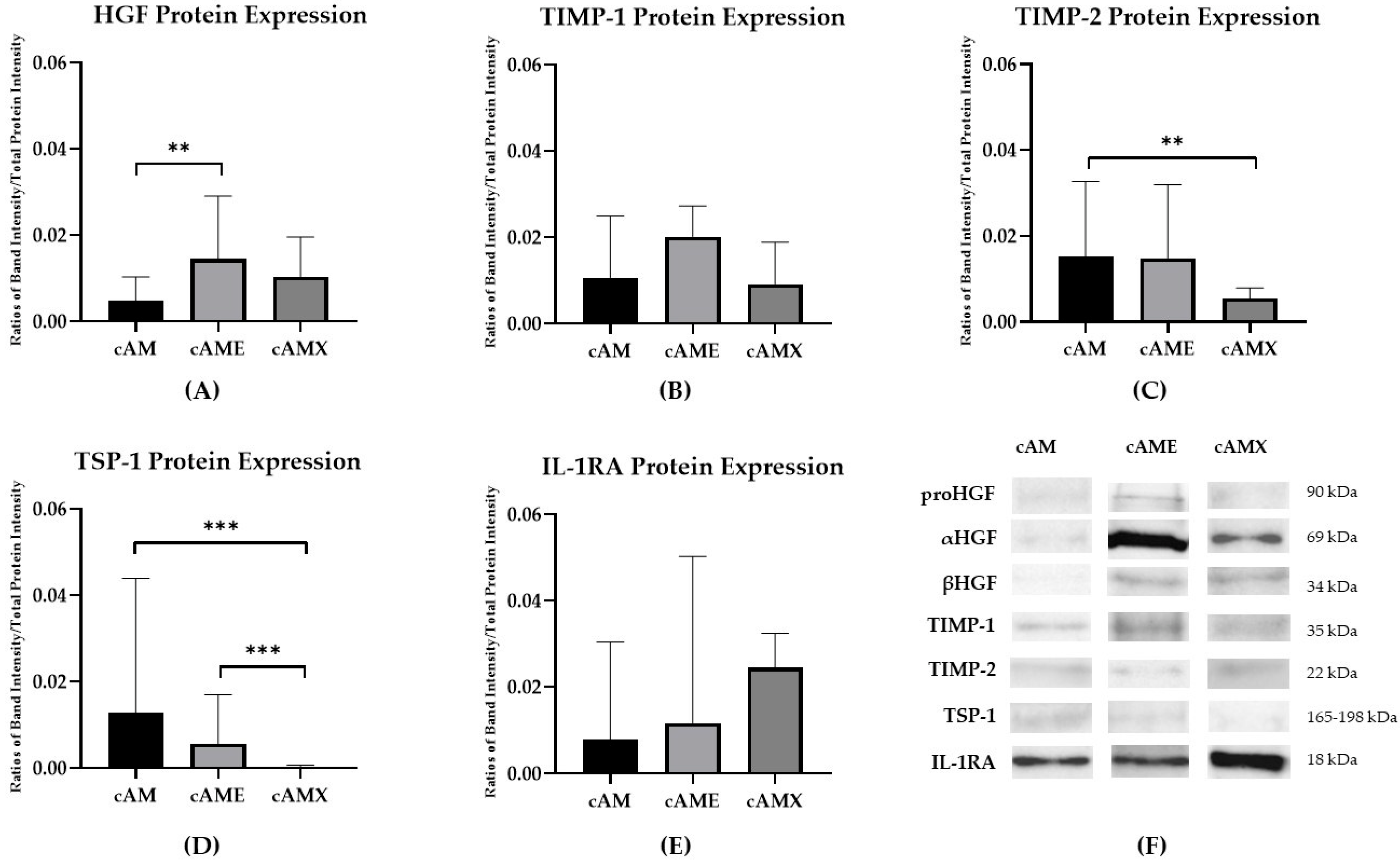
3.2. Biological Compositions Associated with Corneal Wound Healing Were Identified in cAM, cAME and cAMX Both Gene and Protein Expression Levels
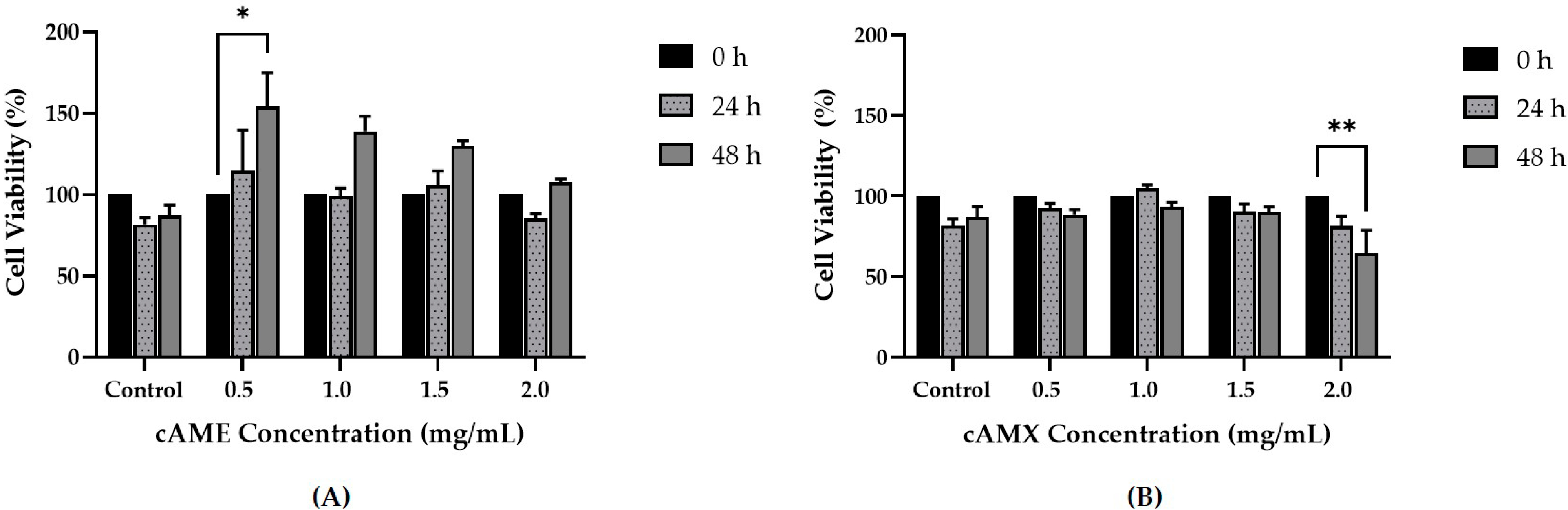
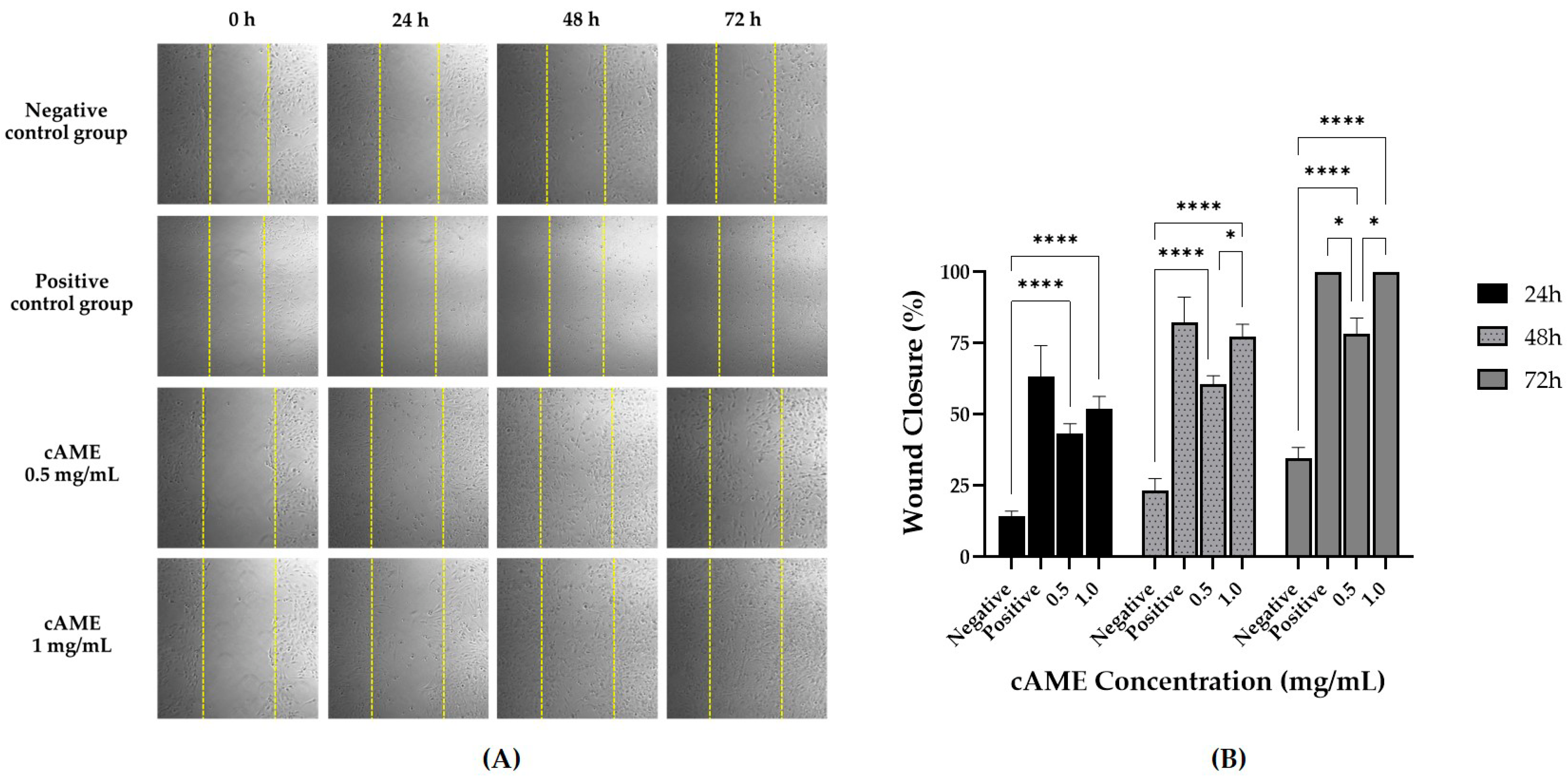
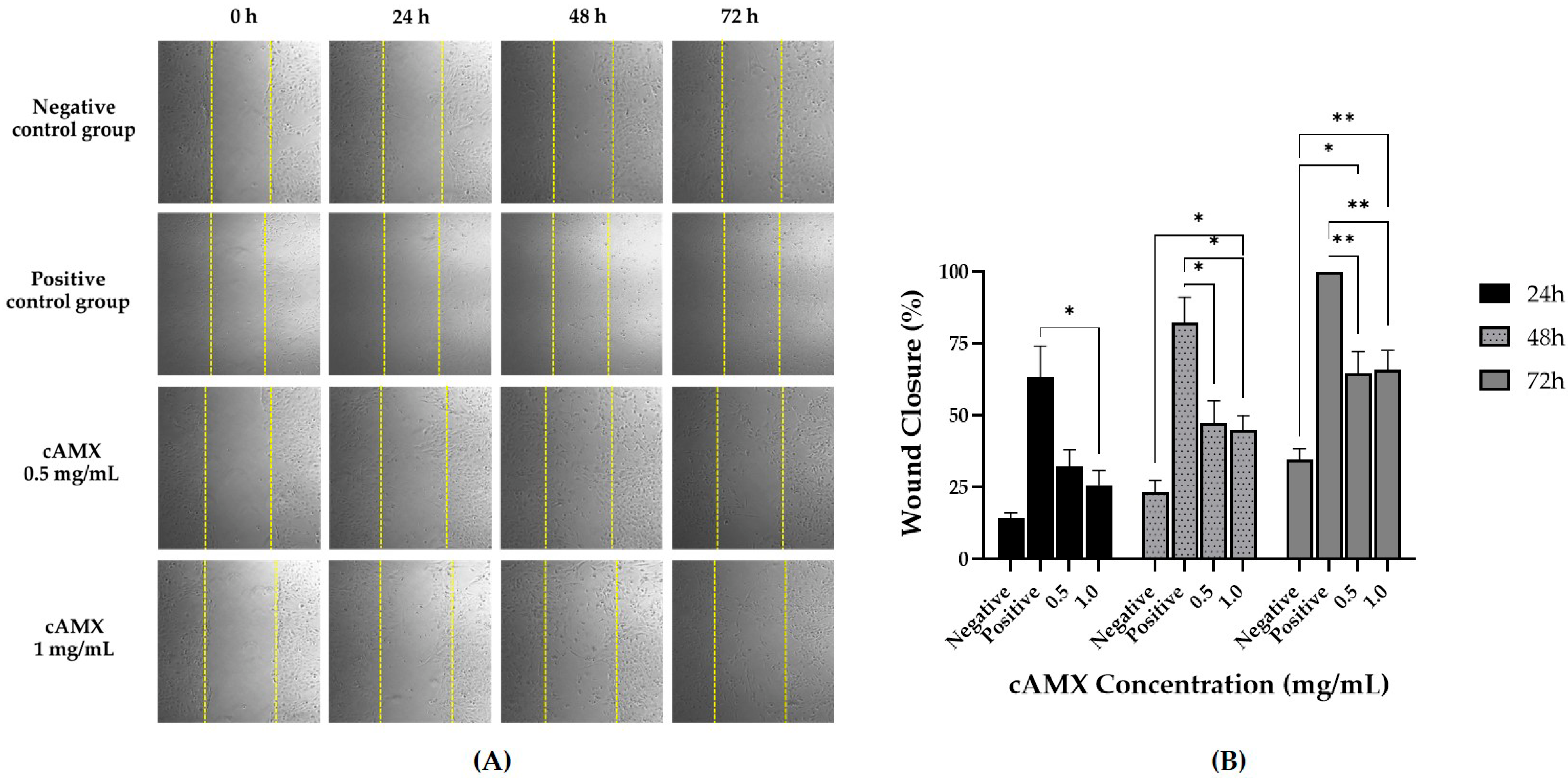
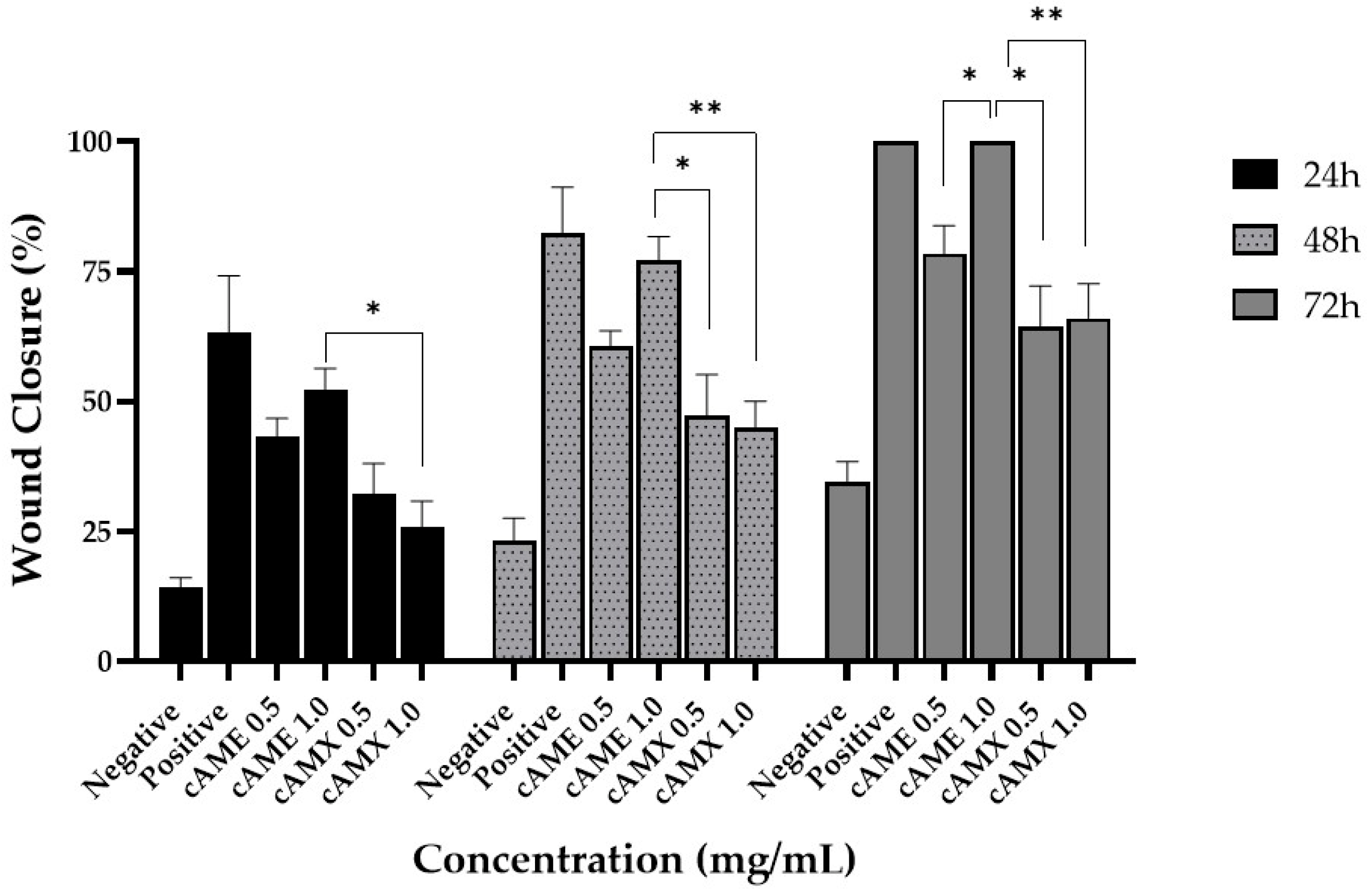
3.3. cAME and cAMX Demonstrated Corneal Wound Healing Efficacy In Vitro
3.3.1. Cell Viability Assay
3.3.2. Wound Healing Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malhotra, C.; Jain, A.K. Human amniotic membrane transplantation: Different modalities of its use in ophthalmology. World J. Transplant. 2014, 4, 111. [Google Scholar] [CrossRef] [PubMed]
- Jirsova, K.; Jones, G.L. Amniotic membrane in ophthalmology: Properties, preparation, storage and indications for grafting—A review. Cell Tissue Bank. 2017, 18, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Dua, H.S.; Gomes, J.A.; King, A.J.; Maharajan, V.S. The amniotic membrane in ophthalmology. Surv. Ophthalmol. 2004, 49, 51–77. [Google Scholar] [CrossRef]
- Woo, H.; Kim, M.; Kweon, O.; Kim, D.; Nam, T.; Kim, J. Effects of amniotic membrane on epithelial wound healing and stromal remodelling after excimer laser keratectomy in rabbit cornea. Br. J. Ophthalmol. 2001, 85, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Koizumi, N.; Inatomi, T.; Sotozono, C.; Fullwood, N.J.; Quantock, A.J.; Kinoshita, S. Growth factor mRNA and protein in preserved human amniotic membrane. Curr. Eye Res. 2000, 20, 173–177. [Google Scholar] [CrossRef]
- Murri, M.S.; Moshirfar, M.; Birdsong, O.C.; Ronquillo, Y.C.; Ding, Y.; Hoopes, P.C. Amniotic membrane extract and eye drops: A review of literature and clinical application. Clin. Ophthalmol. 2018, 12, 1105. [Google Scholar] [CrossRef] [Green Version]
- Arcelli, R.; Tibaldini, P.; Angeli, G.; Bellezza, E. Equine amniotic membrane transplantation in some ocular surface diseases in the dog and cat: A preliminary study. Vet. Res. Commun. 2009, 33, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Choi, Y.M.; Jeong, S.W.; Williams, D.L. Effect of bovine freeze-dried amniotic membrane (Amnisite-BA™) on uncomplicated canine corneal erosion. Vet. Ophthalmol. 2009, 12, 36–42. [Google Scholar] [CrossRef]
- Tsuzuki, K.; Yamashita, K.; Izumisawa, Y.; Kotani, T. Microstructure and glycosaminoglycan ratio of canine cornea after reconstructive transplantation with glycerin-preserved porcine amniotic membranes. Vet. Ophthalmol. 2008, 11, 222–227. [Google Scholar] [CrossRef]
- Barros, P.S.; Safatle, A.M.; Godoy, C.A.; Souza, M.S.; Barros, L.F.; Brooks, D.E. Amniotic membrane transplantation for the reconstruction of the ocular surface in three cases. Vet. Ophthalmol. 2005, 8, 189–192. [Google Scholar] [CrossRef]
- Kalpravidh, M.; Tuntivanich, P.; Vongsakul, S.; Sirivaidyapong, S. Canine amniotic membrane transplantation for corneal reconstruction after the excision of dermoids in dogs. Vet. Res. Commun. 2009, 33, 1003. [Google Scholar] [CrossRef] [PubMed]
- Vongsakul, S.; Tuntivanich, P.; Sirivaidyapong, S.; Kalpravidh, M. Canine amniotic membrane transplantation for ocular surface reconstruction of created deep corneal ulcers in dogs. Thai J. Vet. Med. 2009, 39, 135–144. [Google Scholar]
- Rozin, A.; Cardoso, K.M.; Guimarães, T.G. Clinical Aspects of Amniotic Membrane Implants in Corneal Repair in Dogs with Complicated Ulcers. Acta Sci. Vet. 2020, 48, 534. [Google Scholar] [CrossRef]
- Costa, D.; Leiva, M.; Sanz, F.; Espejo, V.; Esteban, J.; Vergara, J.; Díaz, C.; Huguet, E.; Cairó, M.; Ríos, J. A multicenter retrospective study on cryopreserved amniotic membrane transplantation for the treatment of complicated corneal ulcers in the dog. Vet. Ophthalmol. 2019, 22, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-B.; Seo, M.-S.; Kim, H.-S.; Kang, K.-S. Isolation and characterization of canine amniotic membrane-derived multipotent stem cells. PLoS ONE 2012, 7, e44693. [Google Scholar] [CrossRef] [PubMed]
- Borghesi, J.; Lima, M.F.; Mario, L.C.; da Anunciação, A.R.d.A.; Rabelo, A.C.S.; da Silva, M.G.K.C.; Fernandes, F.A.; Miglino, M.A.; Carreira, A.C.O.; Favaron, P.O. Canine amniotic membrane mesenchymal stromal/stem cells: Isolation, characterization and differentiation. Tissue Cell 2019, 58, 99–106. [Google Scholar] [CrossRef]
- De Oliveira Pinheiro, A.; Lara, V.M.; Souza, A.F.; Casals, J.B.; Bressan, F.F.; Neto, P.F.; Oliveira, V.C.; Martins, D.S.; Ambrosio, C.E. Characterization and immunomodulation of canine amniotic membrane stem cells. Stem Cells Cloning 2020, 13, 43. [Google Scholar] [CrossRef]
- Omoto, M.; Suri, K.; Amouzegar, A.; Li, M.; Katikireddy, K.R.; Mittal, S.K.; Chauhan, S.K. Hepatocyte growth factor suppresses inflammation and promotes epithelium repair in corneal injury. Mol. Ther. 2017, 25, 1881–1888. [Google Scholar] [CrossRef] [Green Version]
- Ries, C. Cytokine functions of TIMP-1. Cell. Mol. Life Sci. 2014, 71, 659–672. [Google Scholar] [CrossRef]
- Sang, Q.X.A. Complex role of matrix metalloproteinases in angiogenesis. Cell Res. 1998, 8, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Lawler, P.R.; Lawler, J. Molecular basis for the regulation of angiogenesis by thrombospondin-1 and -2. Cold Spring Harb. Perspect. Med. 2012, 2, a006627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lawler, J. Thrombospondin-based antiangiogenic therapy. Microvasc. Res. 2007, 74, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stapleton, W.M.; Chaurasia, S.S.; Medeiros, F.W.; Mohan, R.R.; Sinha, S.; Wilson, S.E. Topical interleukin-1 receptor antagonist inhibits inflammatory cell infiltration into the cornea. Exp. Eye Res. 2008, 86, 753–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkinson, A.; McIntosh, R.S.; Tighe, P.J.; James, D.K.; Dua, H.S. Amniotic membrane for ocular surface reconstruction: Donor variations and the effect of handling on TGF-β content. Invest. Ophthalmol. Vis. Sci. 2006, 47, 4316–4322. [Google Scholar] [CrossRef] [Green Version]
- López-Valladares, M.J.; Teresa Rodriguez-Ares, M.; Tourino, R.; Gude, F.; Teresa Silva, M.; Couceiro, J. Donor age and gestational age influence on growth factor levels in human amniotic membrane. Acta Ophthalmol. 2010, 88, e211–e216. [Google Scholar] [CrossRef]
- Litwiniuk, M.; Radowicka, M.; Krejner, A.; Śladowska, A.; Grzela, T. Amount and distribution of selected biologically active factors in amniotic membrane depends on the part of amnion and mode of childbirth. Can we predict properties of amnion dressing? A proof-of-concept study. Cent. Eur. J. Immunol. 2018, 43, 97. [Google Scholar] [CrossRef] [Green Version]
- Riau, A.K.; Beuerman, R.W.; Lim, L.S.; Mehta, J.S. Preservation, sterilization and de-epithelialization of human amniotic membrane for use in ocular surface reconstruction. Biomaterials 2010, 31, 216–225. [Google Scholar] [CrossRef]
- Thomasen, H.; Pauklin, M.; Noelle, B.; Geerling, G.; Vetter, J.; Steven, P.; Steuhl, K.-P.; Meller, D. The effect of long-term storage on the biological and histological properties of cryopreserved amniotic membrane. Curr. Eye Res. 2011, 36, 247–255. [Google Scholar] [CrossRef]
- Phoomvuthisarn, P.; Suriyaphol, G.; Tuntivanich, N. Effect of glycerol concentrations and temperatures on epidermal growth factor protein expression in preserved canine amniotic membrane. Cell Tissue Bank. 2019, 20, 579–583. [Google Scholar] [CrossRef]
- Dower, N.M.B.; Ribeiro, A.P.; Gomes, L.G.; de Cássia Martini, A.; Taques, I.I.G.G.; de Almeida, S.L.H.; da Silva, M.I.V.; de Aguiar, D.M. Concentrations of tissue inhibitor of matrix metalloproteinase-1 and hyaluronic acid in canine amniotic membranes cryopreserved for different time points and its effects in dogs with complicated corneal ulcers. Vet. Ophthalmol. 2021, 25, 62–72. [Google Scholar] [CrossRef]
- Stachon, T.; Bischoff, M.; Seitz, B.; Huber, M.; Zawada, M.; Langenbucher, A.; Szentmary, N. Growth factors and interleukins in amniotic membrane tissue homogenate. Klin. Monbl. Augenheilkd. 2015, 232, 858–862. [Google Scholar] [PubMed]
- Russo, A.; Bonci, P.; Bonci, P. The effects of different preservation processes on the total protein and growth factor content in a new biological product developed from human amniotic membrane. Cell Tissue Bank. 2012, 13, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Ma, D.H.; Hwang, D.G.; Kim, W.S.; Zhang, F. Identification of antiangiogenic and antiinflammatory proteins in human amniotic membrane. Cornea 2000, 19, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Capistrano da Silva, E.; Gibson, D.J.; Jeong, S.; Zimmerman, K.L.; Smith-Fleming, K.M.; Martins, B.d.C. Determining MMP-2 and MMP-9 reductive activities of bovine and equine amniotic membranes homogenates using fluorescence resonance energy transfer. Vet. Ophthalmol. 2021, 24, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Hopkinson, A.; McIntosh, R.S.; Shanmuganathan, V.; Tighe, P.J.; Dua, H.S. Proteomic analysis of amniotic membrane prepared for human transplantation: Characterization of proteins and clinical implications. J. Proteome Res. 2006, 5, 2226–2235. [Google Scholar] [CrossRef]
- Da Silva, E.C.; Arrington, J.; Yau, P.M.; Smith-Fleming, K.M.; Canisso, I.F.; da Costa Martins, B. Proteome composition of bovine amniotic membrane and its potential role in corneal healing. Invest. Ophthalmol. Vis. Sci. 2021, 62, 11. [Google Scholar] [CrossRef]
- Galera, P.D.; Ribeiro, C.R.; Sapp, H.L.; Coleman, J.; Fontes, W.; Brooks, D.E. Proteomic analysis of equine amniotic membrane: Characterization of proteins. Vet. Ophthalmol. 2015, 18, 198–209. [Google Scholar] [CrossRef]
- Heng, Y.J.; Liong, S.; Permezel, M.; Rice, G.E.; Quinzio, M.K.D.; Georgiou, H.M. The interplay of the interleukin 1 system in pregnancy and labor. Reprod. Sci. 2014, 21, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Fidel Jr, P.; Romero, R.; Ramirez, M.; Cutright, J.; Edwin, S.; LaMarche, S.; Cotton, D.; Mitchell, M. Interleukin-1 receptor antagonist (IL-1ra) production by human amnion, chorion, and decidua. Am. J. Reprod. Immunol. 1994, 32, 1–7. [Google Scholar] [CrossRef]
- Asl, N.S.; Nejat, F.; Mohammadi, P.; Nekoukar, A.; Hesam, S.; Ebrahimi, M.; Jadidi, K. Amniotic membrane extract eye drop promotes limbal stem cell proliferation and corneal epithelium healing. Cell J. 2019, 20, 459. [Google Scholar]
- Mahbod, M.; Shahhoseini, S.; Khabazkhoob, M.; Beheshtnejad, A.-H.; Bakhshandeh, H.; Atyabi, F.; Hashemi, H. Amniotic membrane extract preparation: What is the best method? J. Ophthalmic. Vis. Res. 2014, 9, 314. [Google Scholar] [PubMed]
- Rodríguez-Ares, M.T.; López-Valladares, M.J.; Touriño, R.; Vieites, B.; Gude, F.; Silva, M.T.; Couceiro, J. Effects of lyophilization on human amniotic membrane. Acta Ophthalmol. 2009, 87, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [Green Version]
- Lemons, J.M.; Feng, X.-J.; Bennett, B.D.; Legesse-Miller, A.; Johnson, E.L.; Raitman, I.; Pollina, E.A.; Rabitz, H.A.; Rabinowitz, J.D.; Coller, H.A. Quiescent fibroblasts exhibit high metabolic activity. PLoS Biol. 2010, 8, e1000514. [Google Scholar] [CrossRef] [Green Version]
- Stepanenko, A.; Dmitrenko, V. Pitfalls of the MTT assay: Direct and off-target effects of inhibitors can result in over/underestimation of cell viability. Gene 2015, 574, 193–203. [Google Scholar] [CrossRef]
- Zhang, Y.; Alexander, P.B.; Wang, X.-F. TGF-β family signaling in the control of cell proliferation and survival. Cold Spring Harb. Perspect. Biol. 2017, 9, a022145. [Google Scholar] [CrossRef] [Green Version]
- Haber, M.; Cao, Z.; Panjwani, N.; Bedenice, D.; Li, W.W.; Provost, P.J. Effects of growth factors (EGF, PDGF-BB and TGF-β1) on cultured equine epithelial cells and keratocytes: Implications for wound healing. Vet. Ophthalmol. 2003, 6, 211–217. [Google Scholar] [CrossRef]
- Corrales, R.M.; Luo, L.; Chang, E.Y.; Pflugfelder, S.C. Effects of osmoprotectants on hyperosmolar stress in cultured human corneal epithelial cells. Cornea 2008, 27, 574–579. [Google Scholar] [CrossRef]
- Deng, R.; Hua, X.; Li, J.; Chi, W.; Zhang, Z.; Lu, F.; Zhang, L.; Pflugfelder, S.C.; Li, D.-Q. Oxidative stress markers induced by hyperosmolarity in primary human corneal epithelial cells. PLoS ONE 2015, 10, e0126561. [Google Scholar] [CrossRef] [Green Version]
- Masterton, S.; Ahearne, M. The effect of calcium and glucose concentration on corneal epithelial cell lines differentiation, proliferation, and focal adhesion expression. Biores. Open Access 2019, 8, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Gstraunthaler, G.; Seppi, T.; Pfaller, W. Impact of culture conditions, culture media volumes, and glucose content on metabolic properties of renal epithelial cell cultures. Cell. Physiol. Biochem. 1999, 9, 150–172. [Google Scholar] [CrossRef] [PubMed]
- Dudok, D.V.; Nagdee, I.; Cheung, K.; Liu, H.; Vedovelli, L.; Ghinelli, E.; Kenyon, K.; Parapuram, S.; Hutnik, C.M. Effects of amniotic membrane extract on primary human corneal epithelial and limbal cells. Clin. Exp. Ophthalmol. 2015, 43, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, H.; Chen, J.; Zhong, X.; Wang, Y.; Wang, Z.; Wang, Y. The impact of different preservation conditions and freezing-thawing cycles on quality of RNA, DNA, and proteins in cancer tissue. Biopreserv. Biobank. 2015, 13, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.C.; Yañez-Soto, B.; Raghunathan, V.K.; Abbott, N.L.; Murphy, C.J. Species variation and spatial differences in mucin expression from corneal epithelial cells. Exp. Eye Res. 2016, 152, 43–48. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences | Accession Number | Amplicon Size (bp) |
|---|---|---|---|
| HGF | Fw: 5′-AAAGGAGATGAGAAACGCAAACAG-3′ Rv: 5′-GGCCTAGCAAGCTTCAGTAATACC-3′ | AB090353.1 | 92 |
| TIMP-1 | Fw: 5′-CTCACCAGAGAACCCACCAT-3′ Rv: 5′-CCTGATGACGATTTGGGAGT-3′ | AB016817.1 | 147 |
| TIMP-2 | Fw: 5′-AGCAGCACCCAGAAGAAGAG-3′ Rv: 5′-GTCCATCCAGAGGCACTCAT-3′ | NM_001003082.1 | 120 |
| TSP-1 | Fw: 5′-CGCGATGGCAGCTGGAAATGTG-3′ Rv: 5′-GGGCAGGGCAGGCAGTTGTAGC-3 | XM_544610.6 | 167 |
| IL-1RA | Fw:5′-GAAGAGACCTTGCAGGATGC-3′ Rv: 5′-GACGGGCACCACATCTAACT-3′ | AY026462.2 | 142 |
| GAPDH | Fw: 5′-TGTCCCCACCCCCAATGTATC-3′ Rv: 5′-CTCCGATGCCTGCTTCACTACCTT-3′ | AB038240.1 | 100 |
| Genes | cAME (cAM) | cAMX (cAM) | cAMX (cAME) | |||
|---|---|---|---|---|---|---|
| Fold Change | p Value | Fold Change | p Value | Fold Change | p Value | |
| HGF | 1.561 (0.291–12.123) | 0.455 | 0.478 (0.146–1.613) | 0.048 *,b | 0.306 (0.031–2.071) | 0.045 *,c |
| TIMP-1 | 2.162 (0.737–8.165) | 0.024 *,a | 1.669 (0.824–3.116) | 0.008 **,b | 0.772 (0.195–2.143) | 0.404 |
| TIMP-2 | 0.670 (0.233–2.143) | 0.218 | 0.730 (0.274–1.705) | 0.237 | 1.089 (0.330–3.031) | 0.788 |
| TSP-1 | 0.941 (0.198–8.224) | 0.900 | 0.422 (0.174–1.042) | 0.001 **,b | 0.449 (0.058–2.411) | 0.129 |
| IL-1RA | 10.061 (1.133–125.353) | 0.000 ***,a | 2.152 (0.914–5.314) | 0.009 **,b | 0.214 (0.017–1.777) | 0.012 *,c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Permkam, C.; Suriyaphol, G.; Sirisawadi, S.; Tuntivanich, N. Biological Compositions of Canine Amniotic Membrane and Its Extracts and the Investigation of Corneal Wound Healing Efficacy In Vitro. Vet. Sci. 2022, 9, 227. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050227
Permkam C, Suriyaphol G, Sirisawadi S, Tuntivanich N. Biological Compositions of Canine Amniotic Membrane and Its Extracts and the Investigation of Corneal Wound Healing Efficacy In Vitro. Veterinary Sciences. 2022; 9(5):227. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050227
Chicago/Turabian StylePermkam, Chompunut, Gunnaporn Suriyaphol, Sujin Sirisawadi, and Nalinee Tuntivanich. 2022. "Biological Compositions of Canine Amniotic Membrane and Its Extracts and the Investigation of Corneal Wound Healing Efficacy In Vitro" Veterinary Sciences 9, no. 5: 227. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050227