
Aquatic Biological Diversity Responses to Flood Disturbance and Forest Management in Small, Forested Watersheds

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
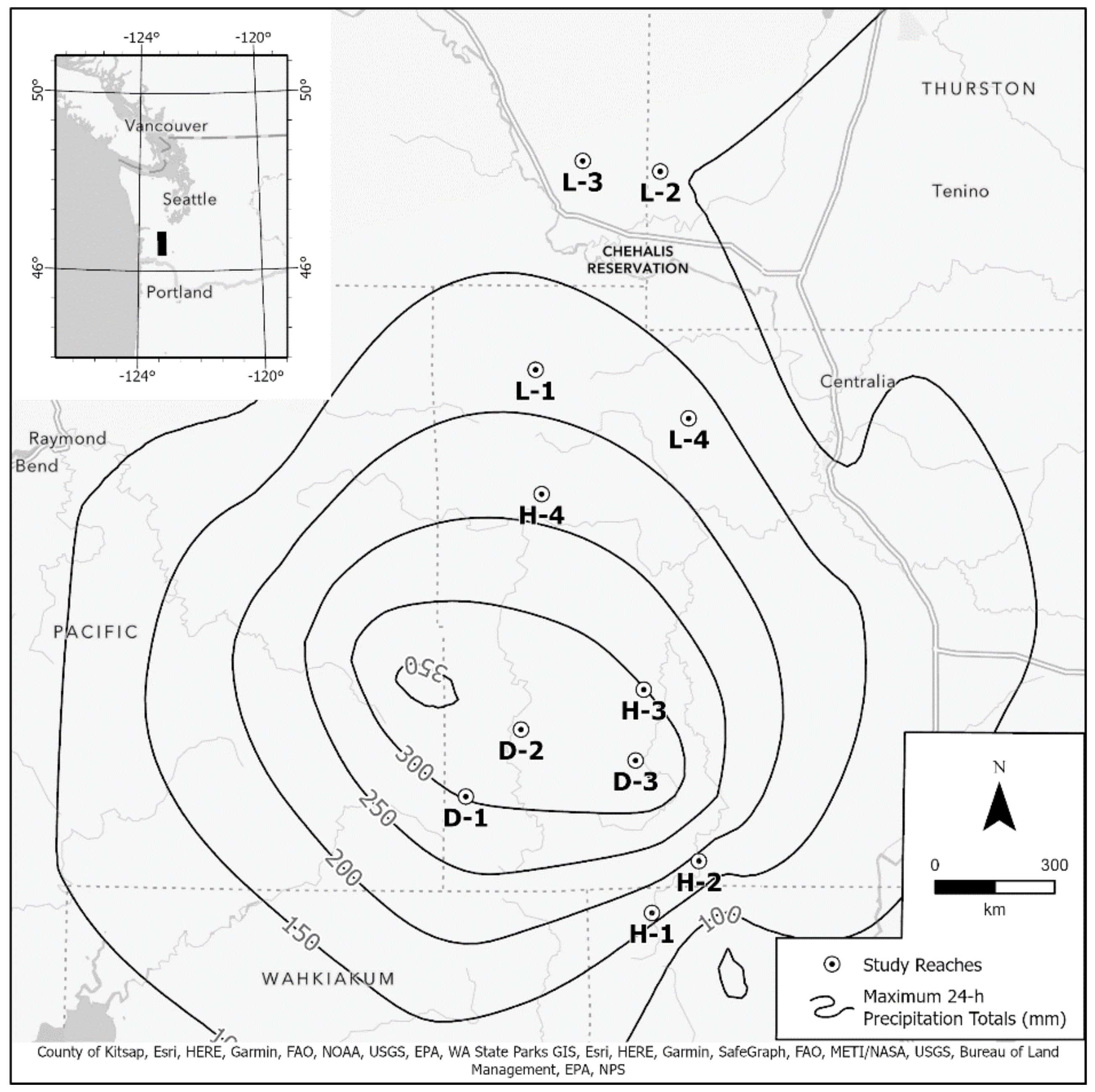
2.1. Study Reaches
2.2. Reach Description
2.3. Physical Habitat Characterization
2.4. Macroinvertebrate Sampling
2.5. Data Analysis
3. Results
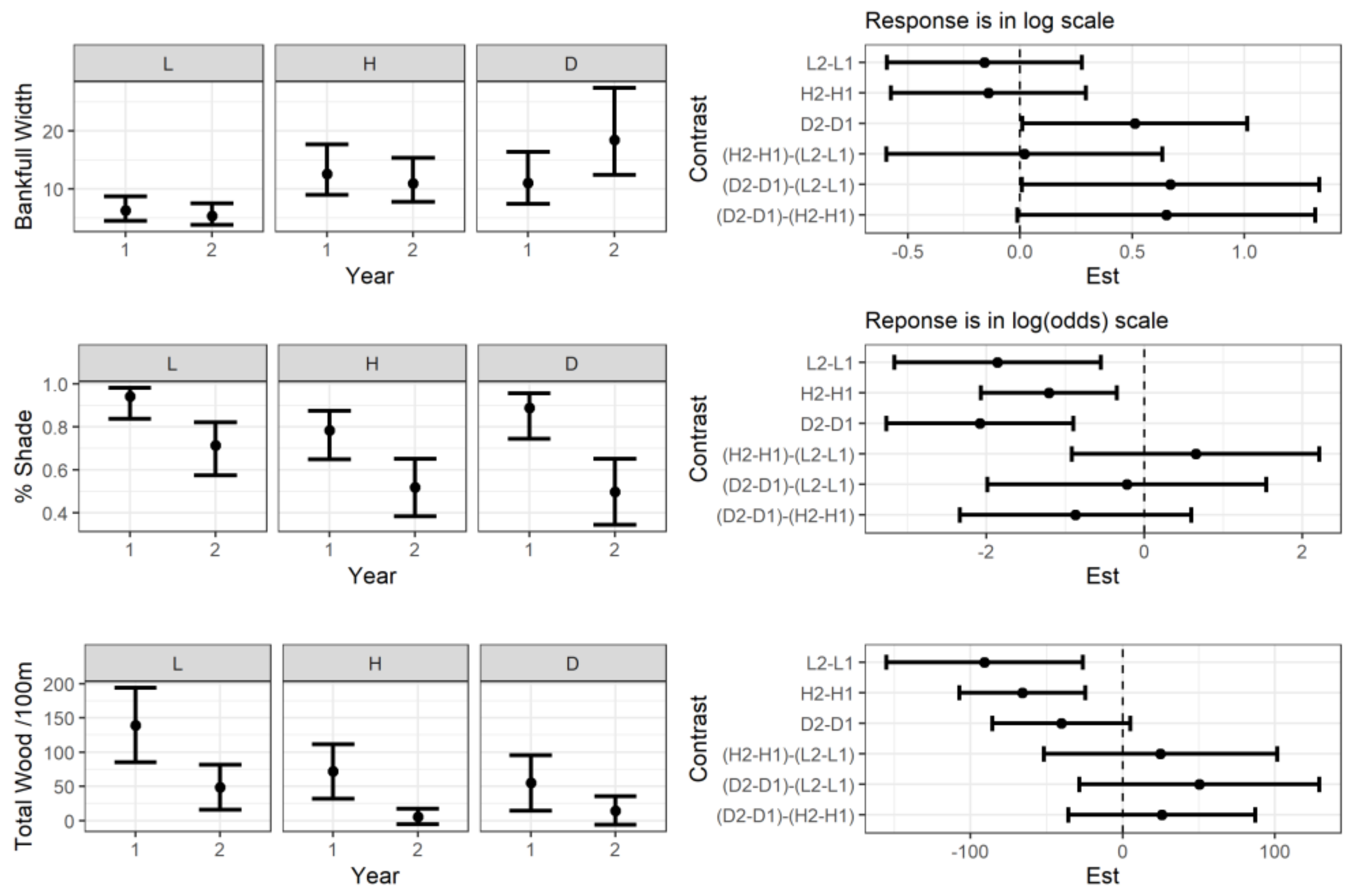
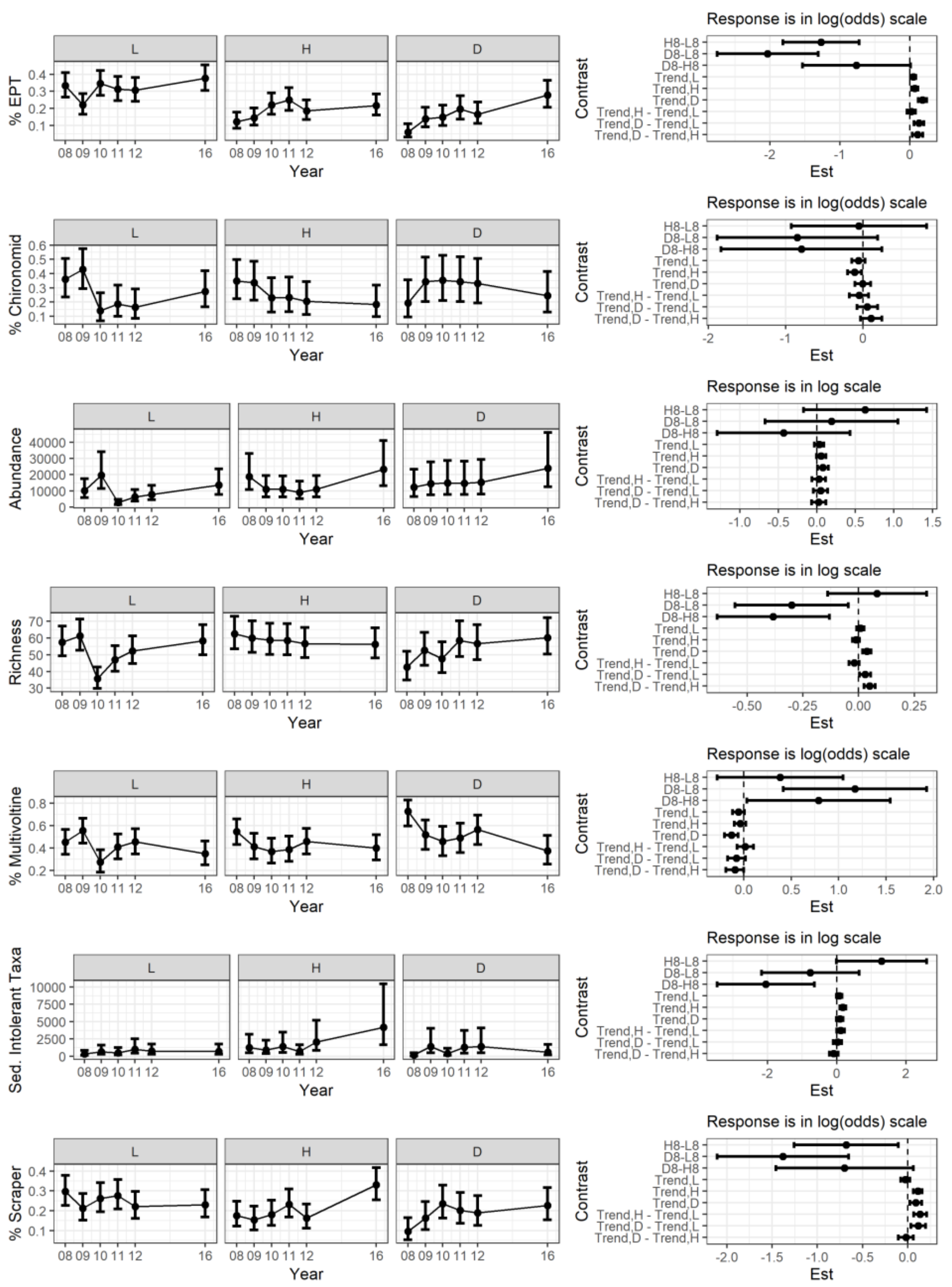
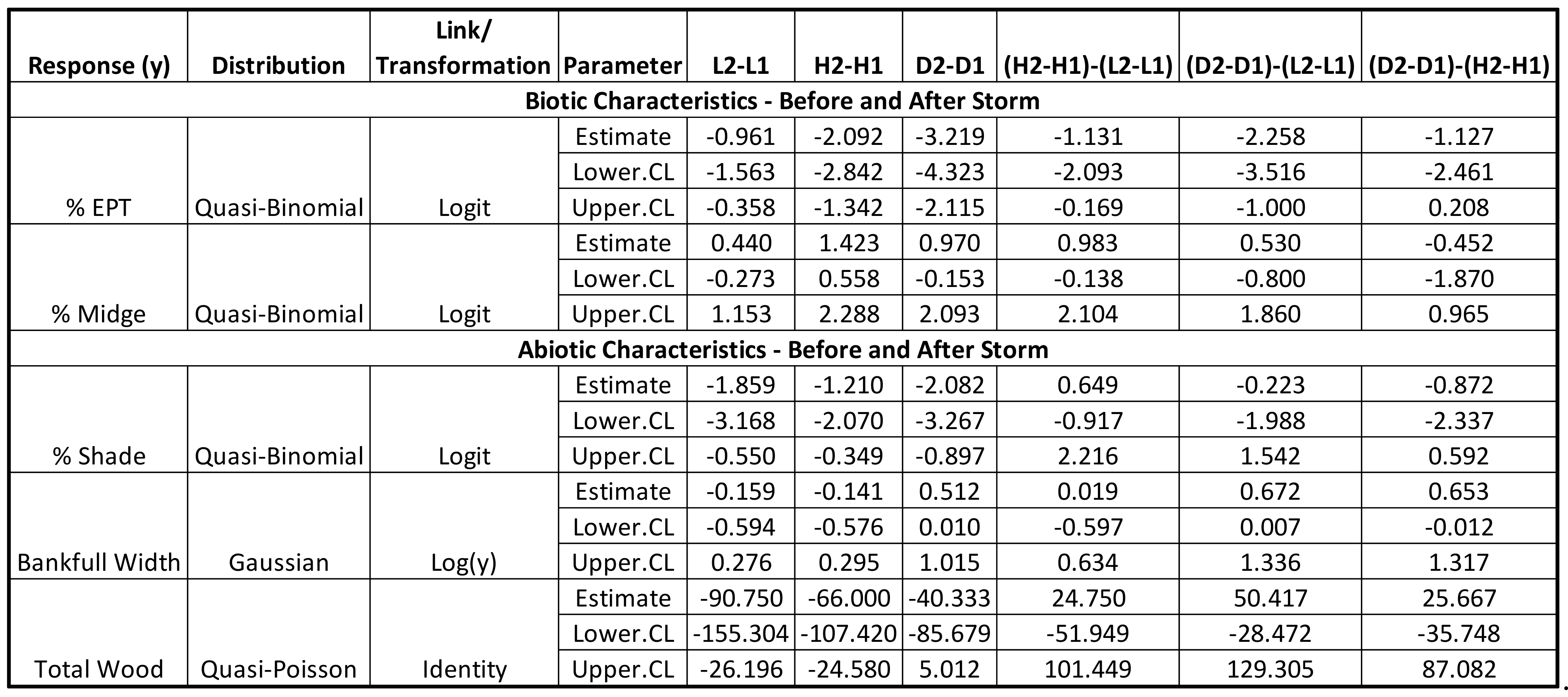
3.1. Abiotic and Biotic Changes before and after Storm
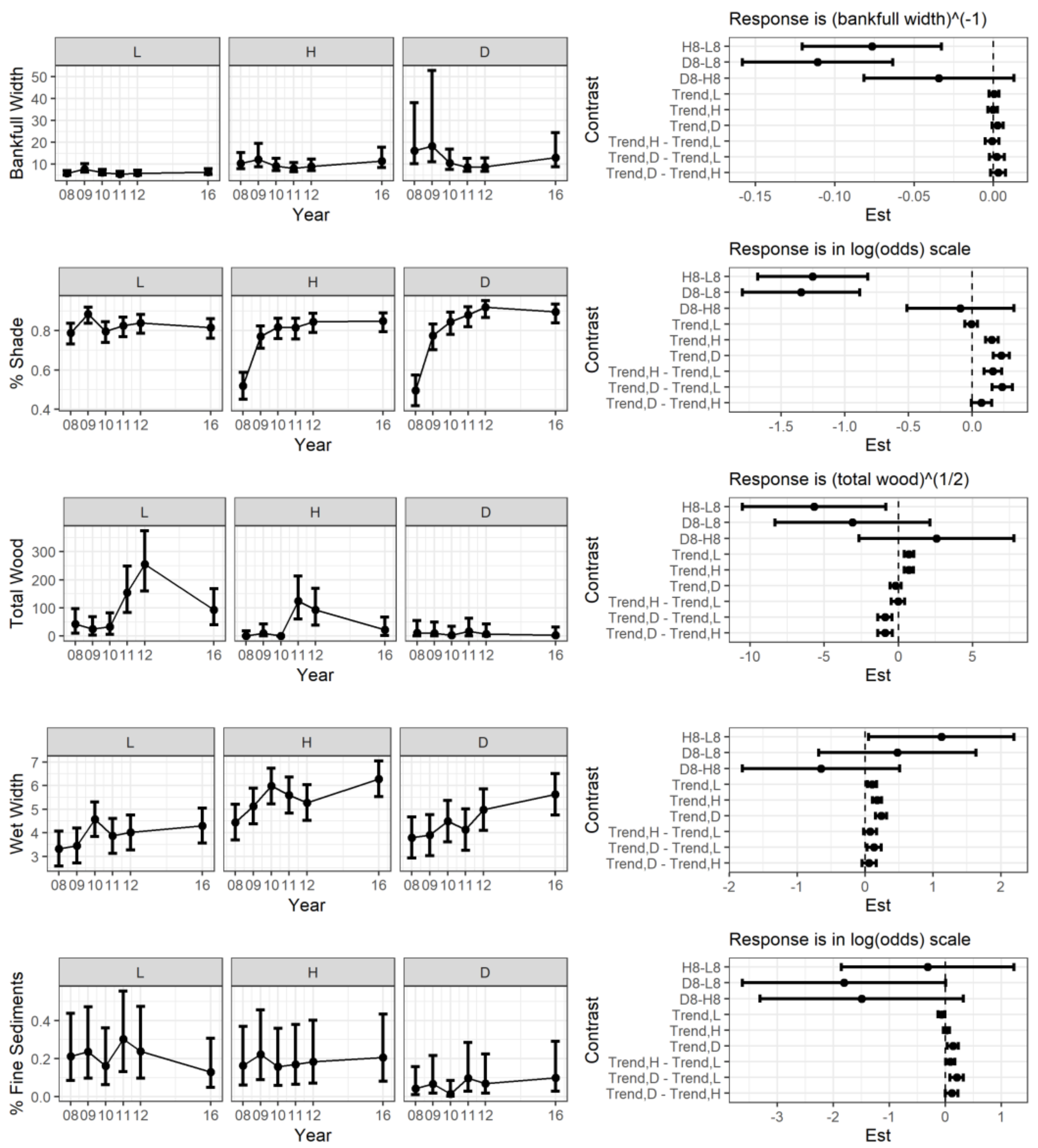
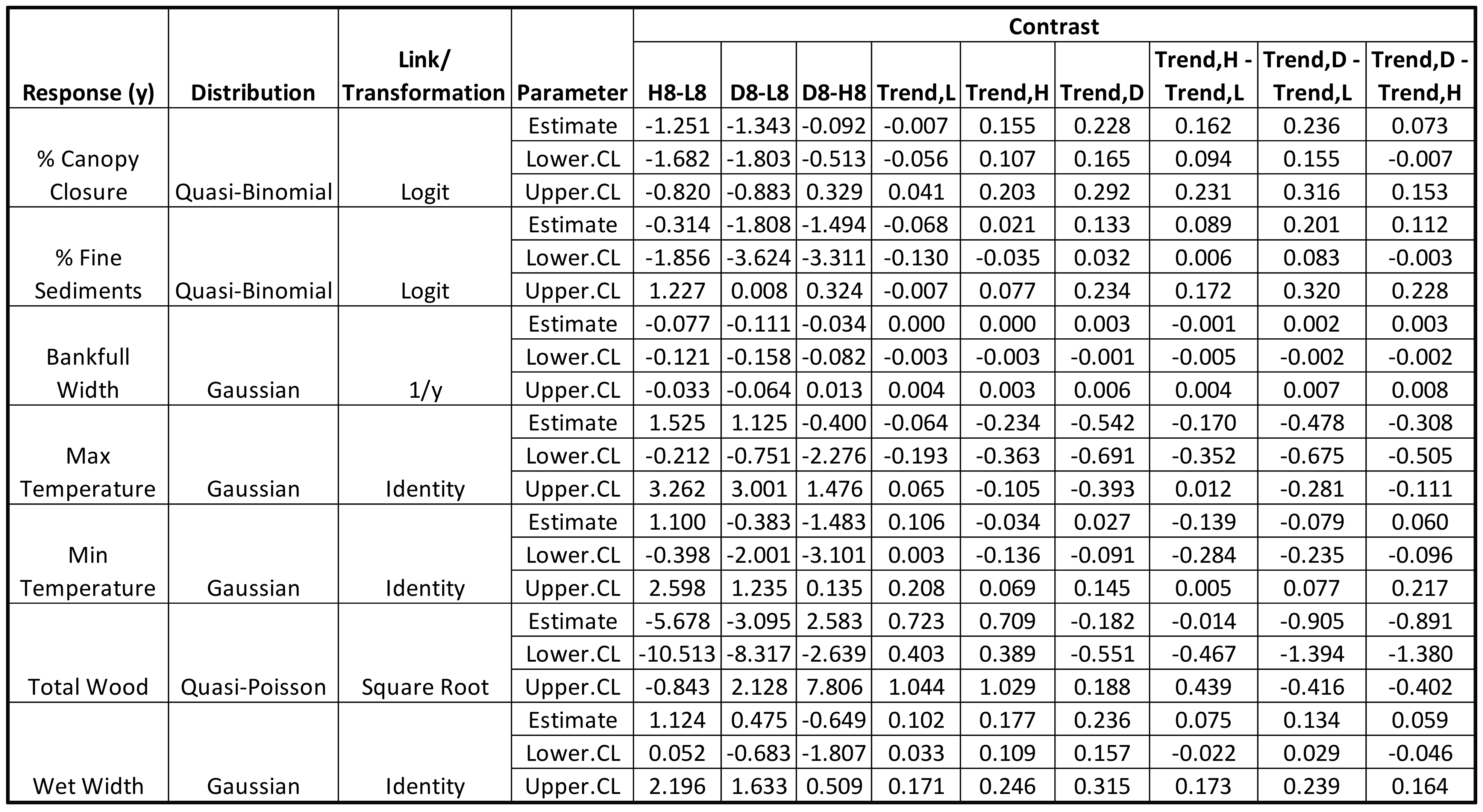
3.2. Stream Temperature and Physical Features Changes after the Storm
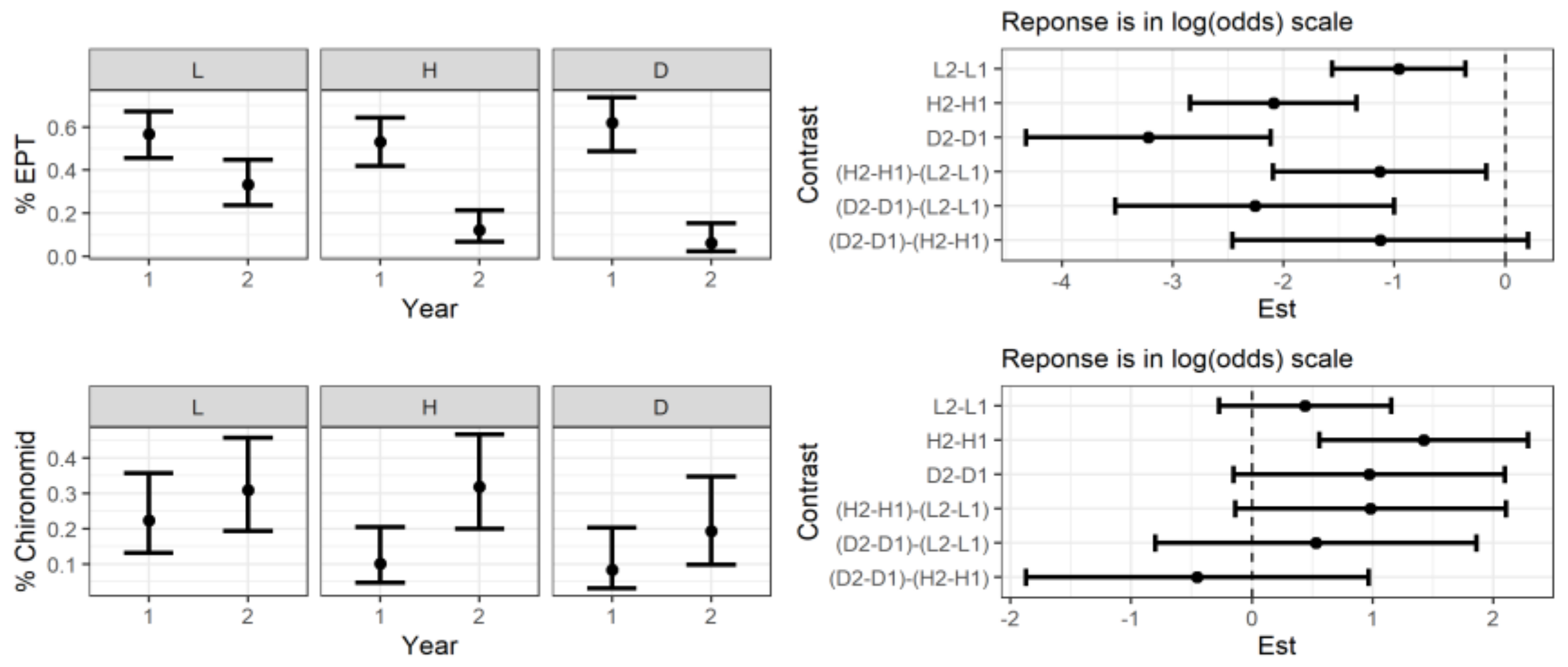
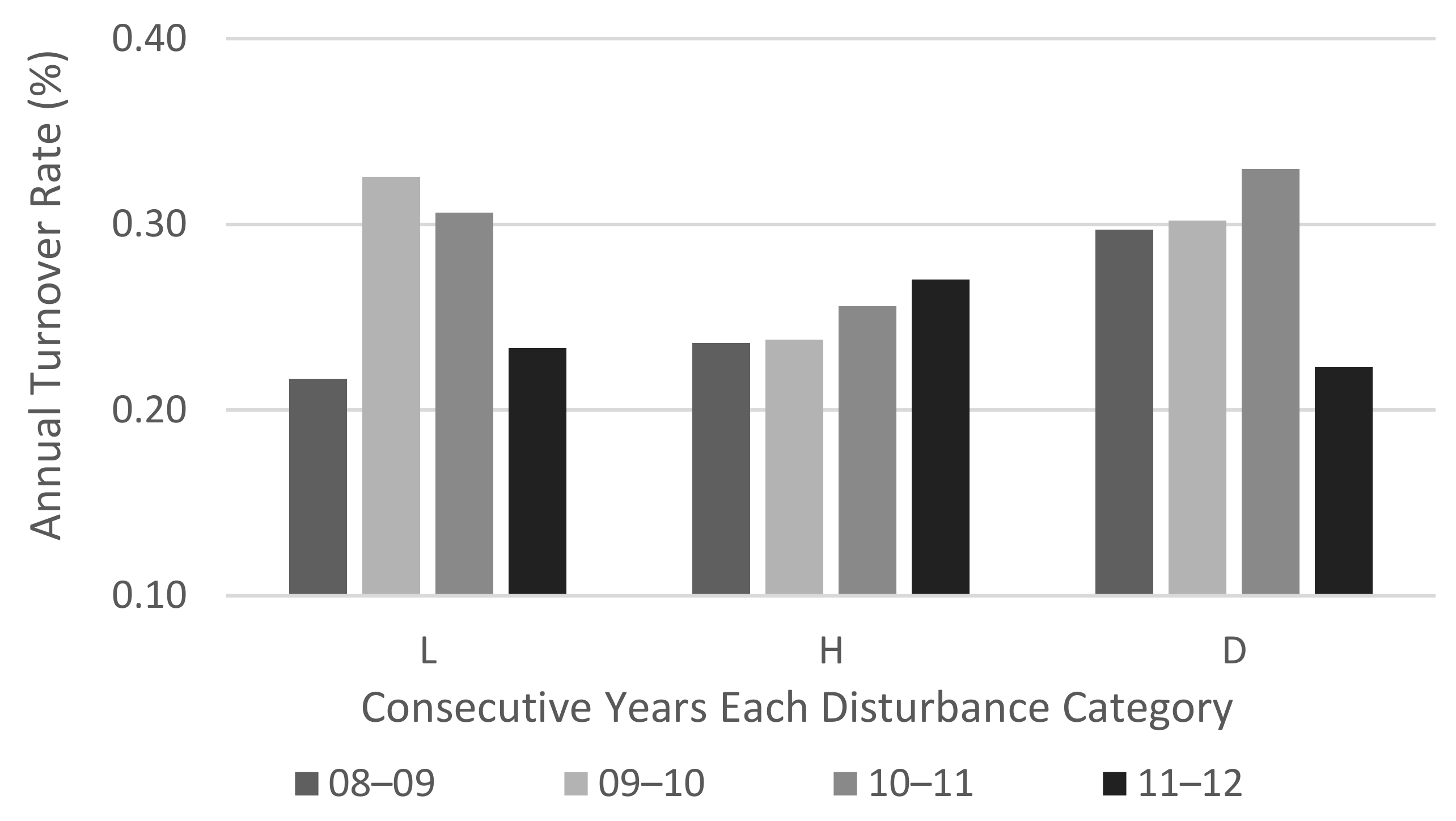
3.3. Macroinvertebrates Changes after the Storm
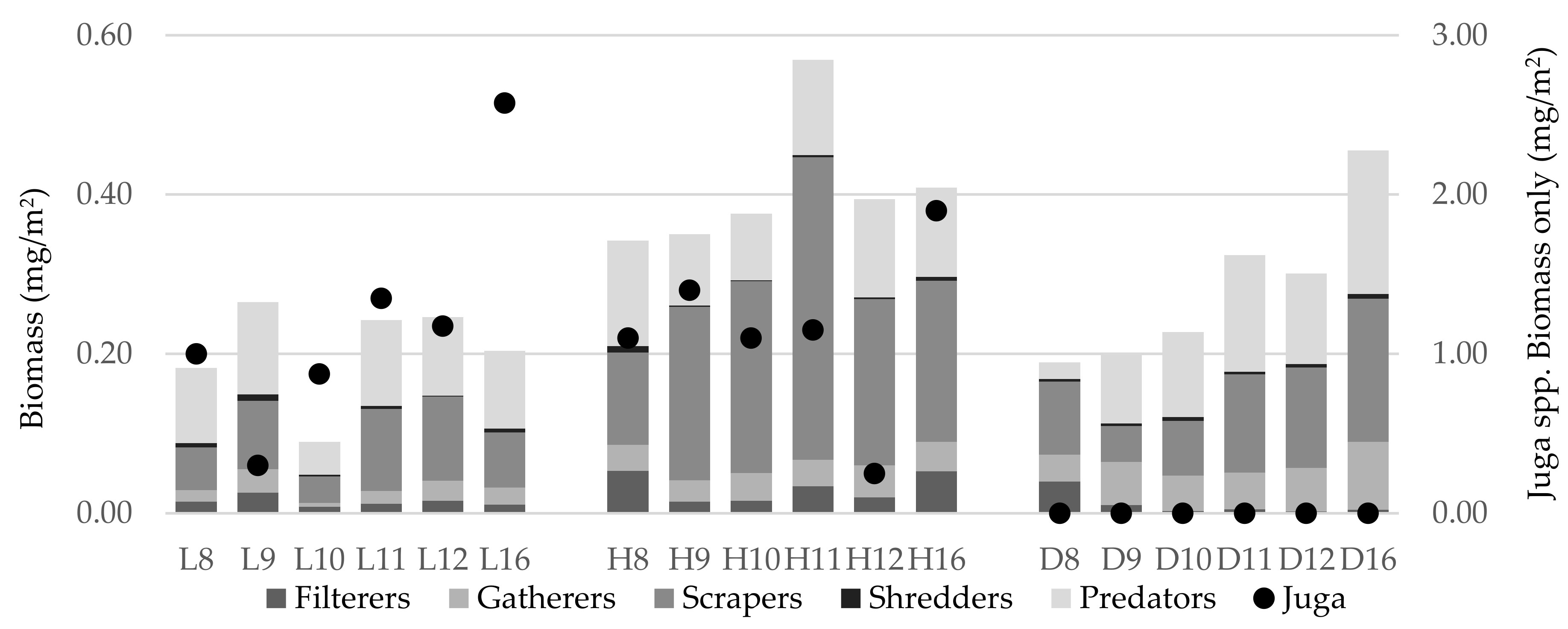
3.4. Biomass
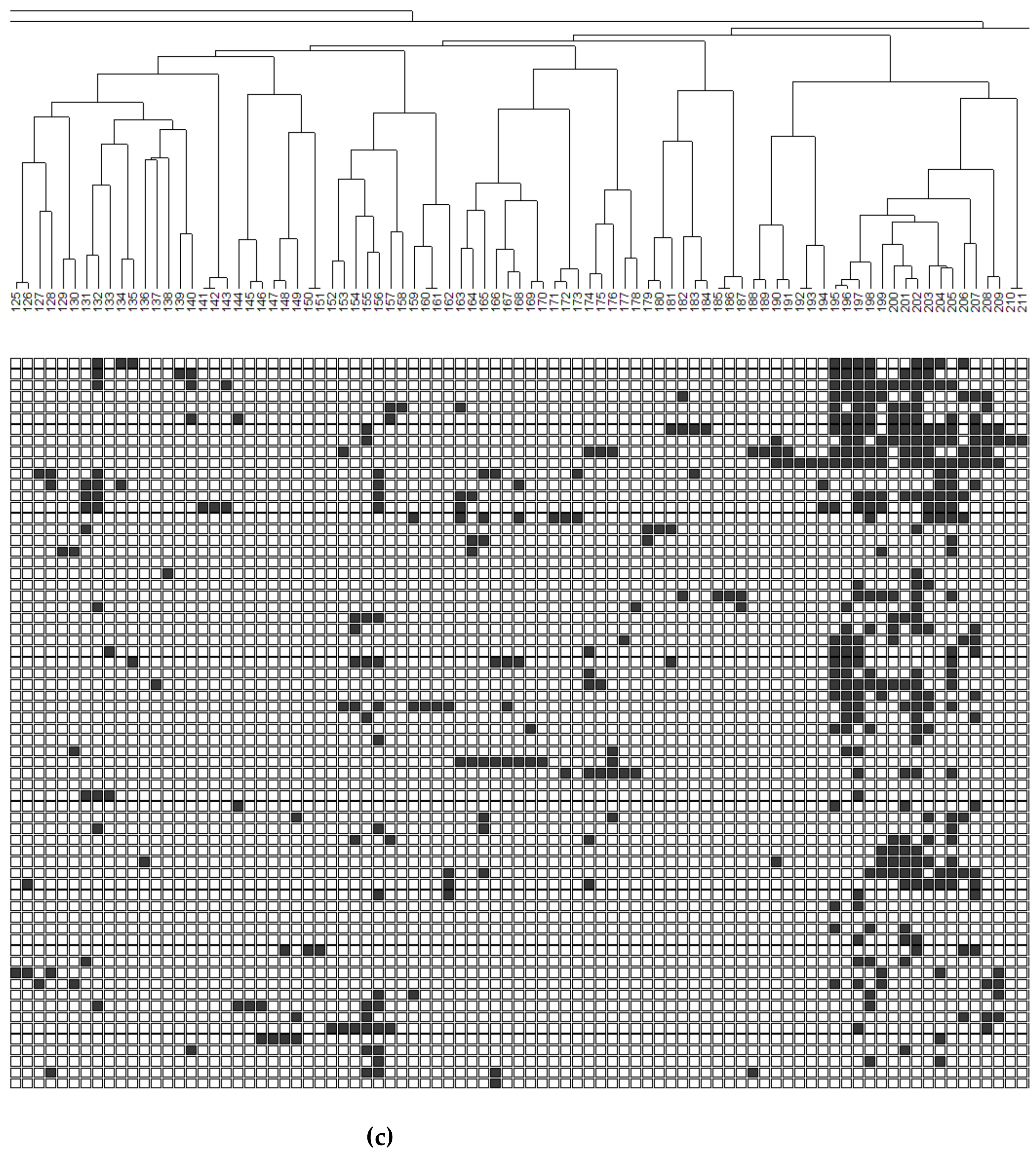
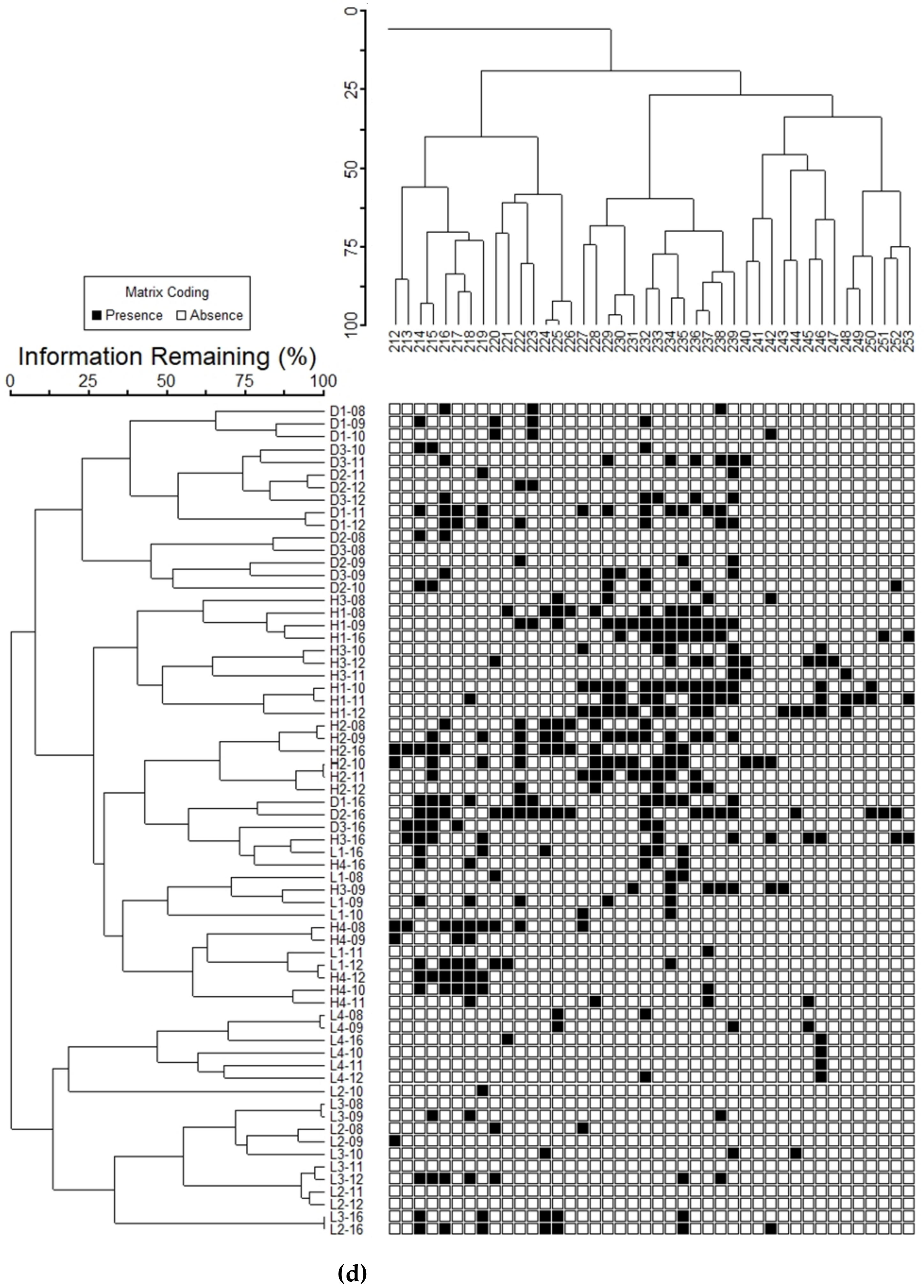
3.5. Community Organization
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

Appendix C

References
- Moore, R.D.; Richardson, J.S. Natural disturbance and forest management in riparian zones: Comparison of effects at reach, catchment, and landscape scales. Freshw. Sci. 2012, 31, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Reeves, G.H.; Benda, L.E.; Burnett, K.M.; Bisson, P.A.; Sedell, J.R. A disturbance-based ecosystem approach to maintaining and restoring freshwater habitats of evolutionarily significant units of anadromous salmonids in the Pacific Northwest. Am. Fish. Soc. Symp. 1995, 17, 334–349. [Google Scholar]
- Bisson, P.A.; Bilby, R.E.; Bryant, M.D.; Dolloff, C.A.; Grette, G.B.; House, R.A.; Sedell, J.R. Large woody debris in forested streams in the Pacific Northwest: Past, present, and future. In Streamside Management: Forestry and Fisheries Interactions; Salo, E.O., Cundy, T.W., Eds.; Contribution No. 57; Institute of Forest Resources, University of Washington: Seattle, WA, USA, 1987. [Google Scholar]
- Imaizumi, F.; Sidle, R.C.; Kamei, R. Effects of forest harvesting on the occurrence of landslides and debris flows in steep terrain in central Japan. Earth Surf. Process. Landf. 2008, 33, 827–840. [Google Scholar] [CrossRef]
- Turner, T.R.; Duke, S.D.; Fransen, B.R.; Reiter, M.L.; Kroll, A.J.; Ward, J.W.; Bach, J.L.; Justice, T.E.; Bilby, R.E. Landslide densities associated with rainfall, stand age, and topography on forested landscapes, southwestern Washington, USA. For. Ecol. Manag. 2010, 259, 2233–2247. [Google Scholar] [CrossRef]
- Bixby, R.J.; Cooper, S.D.; Gresswell, R.E.; Brown, L.E.; Dahm, C.N.; Dwire, K.A. Fire effects on aquatic ecosystems: An assessment of the current state of the science. Freshw. Sci. 2015, 34, 1340–1350. [Google Scholar] [CrossRef]
- Goetz, J.N.; Guthrie, R.H.; Brenning, A. Forest harvesting is associated with increased landslide activity during an extreme rainstorm on Vancouver Island, Canada. Nat. Hazards Earth Syst. Sci. 2014, 15, 1311–1330. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.S. Aquatic arthropods and forestry: Large-scale land-use effects on aquatic systems in nearctic temperate regions. Can. Entomol. 2008, 140, 495–509. [Google Scholar] [CrossRef]
- Grant, G.E.; Swanson, F.J. Morphology and processes of valley floors in mountain streams, Western Cascades, Oregon. In Natural and Anthropogenic Influences in Fluvial Geomorphometry; Costa, J.E., Miller, A.J., Potter, K.W., Wilcock, P., Eds.; Geophysical Monograph Series 89; American Geophysical Union: Washington, DC, USA, 1995; pp. 83–101. [Google Scholar] [CrossRef]
- Wohl, E.; Goode, J.R. Wood dynamics in headwater streams of the Colorado Rocky Mountains. Water Resour. Res. 2008, 44, W09429. [Google Scholar] [CrossRef] [Green Version]
- Le, C.T.; Paul, W.L.; Gawne, B.; Suter, P.J. Insight into the multi-decadal effects of floods on aquatic macroinvertebrate community structure in the Murray River using distributed lag nonlinear models and counterfactual analysis. Sci. Total. Environ. 2021, 757, 143988. [Google Scholar] [CrossRef]
- Anderson, N.H. Influence of disturbance on insect communities in Pacific Northwest streams. Hydrobiologia 1992, 248, 79–92. [Google Scholar] [CrossRef]
- McCabe, D.; Gotelli, N. Effects of disturbance frequency, intensity, and area on assemblages of stream macroinvertebrates. Oecologia 2000, 124, 270–279. [Google Scholar] [CrossRef]
- Steiger, J.; Tabacchi, E.; Dufour, S.; Jean-François Corenblit, D.; Peiry, J. Hydrogeomorphic processes affecting riparian habitat within alluvial channel-floodplain river systems: A review for the temperate zone. River Res. Appl. 2005, 21, 719–737. [Google Scholar] [CrossRef]
- Richardson, J.S.; Danehy, R.J. Synthesis of the ecology of headwater streams and their riparian zones in temperate forests. For. Sci. 2007, 53, 131–147. [Google Scholar] [CrossRef]
- Jakob, M.; Friele, P. Frequency and magnitude of debris flows on Cheekye River, British Columbia. Geomorphology 2010, 114, 382–395. [Google Scholar] [CrossRef]
- Johnson, S.L.; Swanson, F.J.; Grant, G.E.; Wondzell, S.M. Riparian forest disturbances by a mountain flood—the influence of floated wood. Hydrol. Process. 2000, 14, 3031–3050. [Google Scholar] [CrossRef]
- Benda, L.E.; Veldhuisen, V.; Black, J. Debris flows as agents of morphological heterogeneity at low-order confluences, Olympic Mountains, Washington. Geol. Soc. Am. Bull. 2003, 115, 1110–1121. [Google Scholar] [CrossRef]
- D’Souza, L.; Reiter, M.; Six, L.J.; Bilby, R.E. Response of vegetation, shade, and stream temperature to debris torrents in two western Oregon watersheds. For. Ecol. Manag. 2011, 261, 2157–2167. [Google Scholar] [CrossRef]
- Kobayashi, S.; Gomi, T.; Sidle, R.C.; Takemon, T. Disturbances structuring macroinvertebrate communities in steep headwater streams: Relative importance of forest clearcutting and debris flow occurrence. Can. J. Fish. Aquat. Sci. 2010, 67, 427–444. [Google Scholar] [CrossRef]
- Lamberti, G.A.; Gregory, S.V.; Ashkenas, L.R.; Wildman, W.C.; Moore, K.M.S. Stream ecosystem recovery following a catastrophic debris flow. Can. J. Fish. Aquat. Sci. 1991, 48, 196–208. [Google Scholar] [CrossRef]
- Minshall, G.W. Responses of stream benthic macroinvertebrates to fire. For. Ecol. Manag. 2003, 178, 155–161. [Google Scholar] [CrossRef]
- Smith, A.J.; Baldigo, B.P.; Duffy, B.T.; George, S.D.; Dresser, B. Resilience of benthic communities to extreme floods in a Catskill Mountain river, New York USA: Implications for water quality monitoring and assessment. Ecol. Indic. 2019, 104, 107–113. [Google Scholar] [CrossRef]
- Nakamura, F.; Swanson, F.J.; Wondzell, S.M. Disturbance regimes of stream and riparian systems—a disturbance-cascade perspective. Hydrol. Process. 2000, 14, 2849–2860. [Google Scholar] [CrossRef]
- Miller, D.; Luce, C.; Benda, L. Time, space, and episodicity of physical disturbance in streams. For. Ecol. Manag. 2003, 178, 121–140. [Google Scholar] [CrossRef] [Green Version]
- Pool, T.K.; Olden, J.D.; Whittier, J.B.; Paukert, C.P. Environmental drivers of fish functional diversity and composition in the Lower Colorado River Basin. Can. J. Fish. Aquat. Sci. 2010, 67, 1791–1807. [Google Scholar] [CrossRef]
- Sousa, W.P. The Role of Disturbance in Natural Communities. Annu. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- May, C.L. Debris flows through different forest age classes in Central Oregon Coast Range. J. Am. Water Resour. Assoc. 2002, 38, 1097–1113. [Google Scholar] [CrossRef]
- Cederholm, C.J.; Johnson, D.H.; Bilby, R.E.; Dominguez, L.G.; Garrett, A.M.; Graeber, W.H.; Greda, E.L.; Kunze, M.D.; Marcot, B.G.; Palmisano, J.F.; et al. Pacific Salmon and Wildlife—Ecological Contexts, Relationships, and Implications for Management; Special Edition Technical Report; Prepared for D. H. Johnson and T. A. O’Neil (Managing directors), Wildlife-Habitat Relationships in Oregon and Washington; Washington Department of Fish and Wildlife: Olympia, WA, USA, 2000; p. 145.
- Bisson, P.A.; Dunham, J.B.; Reeves, G.H. Freshwater ecosystems and resilience of Pacific salmon: Habitat management based on natural variability. Ecol. Soc. 2009, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.H.; Hansen, B.P. An Annotated Check List of Aquatic Insects Collected at Berry Creek, Benton County, Oregon 1960–1984; Occasional Publication Number 2; Systematic Entomology Laboratory, Department of Entomology, Occasional Publication Number 2, Oregon State University: Corvallis, OR, USA, 1987; p. 13. [Google Scholar]
- Richardson, J.S.; Naiman, R.J.; Swanson, F.J.; Hibbs, D.E. Riparian communities associated with Pacific Northwest headwater streams: Assemblages, processes, and uniqueness. J. Am. Water Resour. Assoc. 2005, 41, 935–947. [Google Scholar] [CrossRef]
- Danehy, R.J.; Chan, S.S.; Lester, G.T.; Langshaw, R.B.; Turner, T. Periphyton and macroinvertebrate assemblage structure in headwaters bordered by mature, thinned, and clearcut Douglas-Fir Stands. For. Sci. 2007, 53, 294–307. [Google Scholar] [CrossRef]
- Smith, W.B.; Miles, P.D.; Perry, C.H.; Pugh, S.A. Forest Resources of the United States, 2007; General Technical Report WO-78; United States Department of Agriculture, Forest Service, Washington Office: Washington, DC, USA, 2009; p. 336. [CrossRef] [Green Version]
- Richardson, J.S.; Béraud, S. Effects of riparian forest harvest on streams: A meta-analysis. J. Appl. Ecol. 2014, 51, 1712–1721. [Google Scholar] [CrossRef]
- Roni, P.; Beechie, T.J.; Bilby, R.E.; Leonetti, F.E.; Pollock, M.M.; Pess, G.R. A review of stream restoration techniques and a hierarchical strategy for prioritizing restoration in Pacific Northwest watersheds. N. Am. J. Fish. Manag. 2001, 22, 1–20. [Google Scholar] [CrossRef]
- Warren, D.R.; Keeton, W.S.; Kiffney, P.M.; Kaylor, M.J.; Bechtold, H.A.; Magee, J. Changing forests—changing streams: Riparian forest stand development and ecosystem function in temperate headwaters. Ecosphere 2016, 7. [Google Scholar] [CrossRef]
- Hayslip, G.A.; Herger, L.G. Ecological Condition of Upper Chehalis Basin Streams; EPA-910-R-01-005; U.S. Environmental Protection Agency, Region 10. Seattle: Washington, DC, USA, 2001; p. 39.
- Herlihy, A.T.; Larsen, D.P.; Paulsen, S.G.; Urquhart, N.S.; Rosenbaum, B.J. Designing a spatially balanced randomized site selection process for regional stream surveys: The EMAP mid-Atlantic pilot study. Environ. Monit. Assess. 2000, 63, 95–113. [Google Scholar] [CrossRef]
- Kaufmann, P.R.; Levine, P.; Robison, E.G.; Seeliger, C.; Peck, D.V. Quantifying Physical Habitat in Wadeable Streams; EPA/620/R-99/003; U.S. Environmental Protection Agency: Washington, DC, USA, 1999; p. 212.
- Lemmon, P.E. A new instrument for measuring forest overstory density. J. For. 1957, 55, 667–669. [Google Scholar]
- Merritt, R.W.; Cummins, W.W.; Berg, M.B. An Introduction to the Aquatic Insects of North. America, 4th ed.; Kendall/Hunt Publishing Company: Dubuque, IA, USA, 2008. [Google Scholar] [CrossRef]
- Relyea, C.D.; Minshall, G.W.; Danehy, R.J. Development and validation of an aquatic fine sediment biotic index. Environ. Manag. 2012, 49, 242–252. [Google Scholar] [CrossRef]
- Breslow, N.E.; Clayton, D.G. Approximate inference in generalized linear mixed models. J. Am. Stat. Assoc. 1993, 88, 9–25. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 April 2021).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Linear and Nonlinear Mixed Effects Models. R package version 3.1-144. 2020. Available online: https://CRAN.R-project.org/package=nlme. (accessed on 1 April 2021).
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MJM Software Design: Gleneden Beach, OR, USA, 2002; p. 304. [Google Scholar]
- Lance, G.N.; Williams, W.T. A general theory of classification sorting strategies. I. Hierarchical systems. Comput. J. 1967, 9, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Madeira, S.C.; Oliveira, A.L. Biclustering algorithms for biological data analysis: A survey. IEEE Trans. Comput. Biol. Bioinform. 2004, 1, 24–46. [Google Scholar] [CrossRef]
- Nichols, J.D.; Boulinier, T.; Hines, J.E.; Pollock, K.H.; Sauer, J.R. Estimating rates of species extinction, colonization and turnover in animal communities. Ecol. Appl. 1998, 8, 1213–1225. [Google Scholar] [CrossRef]
- Stone, P.H.; Carlson, J.H. Atmospheric Lapse Rate Regimes and Their Parameterization. J. Atmos. Sci. 1979, 36, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Poole, G.C.; Berman, C.H. An ecological perspective on in-stream temperature: Natural heat dynamics and mechanisms of human-caused thermal degradation. Environ. Manag. 2001, 6, 787–802. [Google Scholar] [CrossRef]
- Klaar, M.J.; Shelley, F.S.; Hannah, D.M.; Krause, S. Instream wood increases riverbed temperature variability in a lowland sandy stream. River Res. Appl. 2020, 36, 1529–1542. [Google Scholar] [CrossRef]
- Pabst, R.J.; Spies, T.A. Ten years of vegetation succession on a debris-flow deposit in Oregon. J. Am. Water Resour. Assoc. 2001, 37, 1693–1708. [Google Scholar] [CrossRef]
- Bilby, R.E.; Bisson, P.A. Function and Distribution of Large Woody Debris. In River Ecology and Management: Lessons from the Pacific Coastal Ecoregion; Naiman, R.J., Bilby, R.E., Eds.; Springer: New York, NY, USA, 1998; pp. 324–346. [Google Scholar] [CrossRef]
- Welty, J.; Beechie, B.; Sullivan, K.; Hyink, D.; Bilby, R.; Andrus, C.; Pess, G. Riparian Aquatic Interaction Simulator (RAIS): A model of riparian forest dynamics for the generation of large woody debris and shade. For. Ecol. Manag. 2002, 162, s0378–s1127. [Google Scholar] [CrossRef]
- Meleason, M.A.; Gregory, S.V.; Bolte, J.P. Implications of riparian management strategies on wood in streams of the Pacific Northwest. Ecol. Appl. 2003, 13, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Cover, M.R.; de la Fuente, J.A.; Resh, V.H. Catastrophic disturbances in headwater streams: The long-term ecological effects of debris flows and debris floods in the Klamath Mountains, northern California. Can. J. Fish. Aquat. Sci. 2010, 67, 1596–1610. [Google Scholar] [CrossRef]
- Danehy, R.J.; Bilby, R.E.; Langshaw, R.; Evans, D.M.; Turner, T.R.; Floyd, W.C.; Schoenholtz, S.; Duke, S.D. Biological and water quality responses to hydrologic disturbances in third-order forested streams. Ecohydrology 2011, 5, 90–98. [Google Scholar] [CrossRef]
- Snyder, C.D.; Johnson, Z.B. Macroinvertebrate assemblage recovery following a catastrophic flood and debris flows in an Appalachian mountain stream. J. North. Am. Benthol. Soc. 2006, 25, 825–840. [Google Scholar] [CrossRef]
- Baillie, B.R.; Evanson, A.W.; Kimberley, M.O. Combined effects of an anthropogenic (forest harvest) and natural (extreme rainfall event) disturbance on headwater streams in New Zealand. Freshw. Biol. 2020, 65, 1806–1823. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.H. Species Richness: Estimation and Comparison; Wiley StatsRef: Statistics Reference Online, 2016; pp. 1–26. [Google Scholar] [CrossRef]
- Resh, V.R.; Bêche, L.A.; McElravy, E.P. How common are rare taxa in long-term benthic macroinvertebrate surveys? J. N. Am. Benthol. Soc. 2005, 24, 976–989. [Google Scholar] [CrossRef]
- Foster, A.D.; Claeson, S.M.; Bisson, P.A.; Heimburg, J. Aquatic and riparian system recovery from debris flows in two western Washington streams, USA. Ecol. Evol. 2020, 10, 2749–2777. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, S.T.; Hayes, S.K.; Grant, G.E. Effects of wood on debris flow runout in small mountain watersheds. Water Resour. Res. 2003, 39, 1168. [Google Scholar] [CrossRef] [Green Version]
- Gomi, T.; Moore, R.D.; Hassan, M.A. Suspended sediment dynamics in small, forested streams of the pacific Northwest. J. Am. Water Resour. Assoc. 2005, 41, 877–898. [Google Scholar] [CrossRef]
- Biggs, B.J.F. Contribution of disturbance, catchment geology, and landuse to the habitat template of periphyton in stream Ecosystems. Freshw. Biol. 1995, 33, 419–438. [Google Scholar] [CrossRef]
- Naman, S.M.; Rosenfeld, J.S.; Richardson, J.S. Causes and consequences of invertebrate drift in running waters: From individuals to populations and trophic fluxes. Can. J. Fish. Aquat. Sci. 2016, 73, 1292–1305. [Google Scholar] [CrossRef] [Green Version]
- Ferrington, L.C., Jr.; Berg, M.B.; Coffman, W.P. Chironomidae. In An Introduction to the Aquatic Insects of North America, 4th Ed.; Merritt, R.W., Cummins, K.W., Berg, M.B., Eds.; Kendall/Hunt Publishing Co.: Dubuque, IA, USA, 2008; pp. 847–989. [Google Scholar] [CrossRef]
- Johnson, J.H. Comparative diets of subyearling Chinook salmon (Oncorhynchus tshawytscha) and steelhead (O. mykiss) in the Salmon River, New York. J. Great Lakes Res. 2007, 33, 906–911. [Google Scholar] [CrossRef]
- Tilley, L.J. Diel drift of Chironomidae larvae in a pristine Idaho mountain stream. Hydrobiologia 1989, 174, 133–149. [Google Scholar] [CrossRef]
- Grzybkowska, M.; Dukowska, M.; Figiel, K.; Szczerkowska, E.; Tszydel, M. Dynamics of macroinvertebrate drift in a lowland river. Zool. Polpniae 2004, 49, 1–4. [Google Scholar]
- Danehy, R.J.; Bilby, R.E.; Owen, S.; Duke, S.D.; Farrand, A. Interactions of baseflow habitat constraints: Macroinvertebrate drift, stream temperature, and physical habitat for anadromous salmon in the Calapooia River, Oregon. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 653–662. [Google Scholar] [CrossRef]
- Weber, N.; Bouwes, N.; Jordan, C.E.; Jonsson, B. Estimation of salmonid habitat growth potential through measurements of invertebrate food abundance and temperature. Can. J. Fish. Aquat. Sci. 2014, 71, 1158–1170. [Google Scholar] [CrossRef]
- Naman, S.M.; Rosenfeld, J.S.; Kiffney, P.M.; Richardson, J.S. The energetic consequences of habitat structure for forest stream salmonids. J. Anim. Ecol. 2018, 87, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Agee, J.K. Fire Ecology of Pacific Northwest. Forests; Island Press: Washington, DC, USA, 1993; p. 505. [Google Scholar] [CrossRef]
- May, C.L.; Gresswell, R.E. Processes and rates of sediment and wood accumulation in headwater streams of the Oregon Coast Range, USA. Earth Surf. Process. Landf. 2003, 28, 409–424. [Google Scholar] [CrossRef]
- Adams, P. Policy and management for headwater streams in the Pacific Northwest: Synthesis and reflection. For. Sci. 2007, 53, 104–119. [Google Scholar] [CrossRef]
- Mcintosh, B.A.; Sedell, J.R.; Thurow, R.F.; Clarke, S.E.; Chandler, G.L. Historical changes in pool habitats in the Columbia River basin. Ecol. Appl. 2000, 10, 1478–1496. [Google Scholar] [CrossRef]
- Parson, G.L.; Cassis, G.; Moldenke, A.R.; Lattin, J.D.; Anderson, N.H.; Miller, J.C.; Hammond, P.; Schowalter, T.D. Invertebrates of the H.J. Andrews Experimental Forest, Western Cascade Range, Oregon. V: An. Annotated List of Insects and Other Arthropods; Gen. Tech. Rep. PNW-GTR-290; US Department of Agriculture Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1991; p. 180. [CrossRef]
- Meyer, J.L.; Strayer, D.L.; Wallace, J.B.; Eggert, S.L.; Helfman, G.S.; Leonard, N.E. The contribution of headwater streams to biodiversity in river networks. J. Am. Water Resour. Assoc. 2007, 43, 86–103. [Google Scholar] [CrossRef] [Green Version]
- Olson, D.H.; Andersen, P.D.; Frissell, C.A.; Welsh, H.H.; Bradford, D.F. Biodiversity management approaches for stream riparian areas: Perspectives from Pacific Northwest headwater forests. Microclimate and amphibians. For. Ecol. Manag. 2007, 246, 81–107. [Google Scholar] [CrossRef]
- Danehy, R.J.; Johnson, S.L. Applying four principles of headwater system aquatic biology to forest management. In Density Management in the 21st Century: West Side Story; Anderson, P.D., Ronnenberg, K.L., Eds.; USDA Forest Service: Portland, OR, USA, 2013; pp. 189–202. [Google Scholar] [CrossRef]
- Richardson, J.S. Biological Diversity in Headwater Streams. Water 2019, 11, 366. [Google Scholar] [CrossRef] [Green Version]
- Bilby, R.E.; Danehy, R.J.; Jones, K.K. Woodless rivers in the middle of forests. In Wood in World Rivers, Proceedings of the Third International Conference; Picco, L., Lenzi, M.A., Bertoldi, W., Comiti, F., Rigon, E., Tonon, A., García-Rama, A., Ravazzolo, D., Rainato, R., Eds.; Cooperativa Libraria Editrice Università di Padova Via Belzoni 118/3: Padova, Italy, 2015; pp. 84–86. [Google Scholar]
- Colvin, S.; Sullivan, P.; Shirey, P.D.; Colvin, R.W.; Winemiller, K.O.; Hughes, R.M.; Fausch, K.D.; Infante, D.M.; Olden, J.D.; Bestgen, K.R.; et al. Headwater streams and wetlands are critical for sustaining fish, fisheries, and ecosystem services. Fisheries. Headwater Streams and Wetlands are Critical for Sustaining Fish, Fisheries, and Ecosystem Services. Fisheries 2019, 44, 73–91. [Google Scholar] [CrossRef]
- Bilby, R.E.; Ward, J.W. Changes in characteristics and function of large woody debris in streams draining old-growth, clear-cut, and second-growth forests in southwestern Washington. Can. J. Fish. Aquat. Sci. 2019, 48, 2499–2508. [Google Scholar] [CrossRef]
- Sabo, J.L.; Sponseller, R.; Dixon, M.; Gade, K.; Harms, T.; Heffernan, J.; Jani, A.; Katz, G.; Soykan, C.; Watts, J.; et al. Riparian zones increase regional species richness by harboring different, not more, species. Ecology 2005, 86, 56–62. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H.; Pollock, M. The Role of Riparian Corridors in Maintaining Regional Biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L1 | L2 | L3 | L4 | H1 | H2 | H3 | H4 | D1 | D2 | D3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean Basin Slope (%) | 25 | 19 | 16 | 20 | 31 | 34 | 25 | 30 | 50 | 39 | 47 |
| Basin % Canopy | 80 | 78 | 77 | 80 | 53 | 50 | 73 | 72 | 92 | 74 | 90 |
| Watershed Area (km2) | 7.1 | 9.1 | 7.9 | 12.9 | 45.1 | 14.3 | 28.5 | 11.2 | 5.3 | 18 | 9.7 |
| Mean Basin Elevation(m) | 372 | 293 | 254 | 165 | 387 | 393 | 326 | 390 | 552 | 567 | 524 |
| Maximum Elevation (m) | 686 | 421 | 418 | 272 | 811 | 756 | 945 | 737 | 799 | 988 | 925 |
| Baseflow (m3/s) | 0.02 | 0.02 | 0.05 | 0.01 | 0.13 | 0.04 | 0.05 | 0.04 | 0.01 | 0.11 | 0.02 |
| Reach Slope (%) | 4.08 | 2.7 | 6.1 | 0.49 | 0.57 | 2 | 0.19 | 1.1 | 4.05 | 3.4 | 1.82 |
| Response (y) | Distribution | Link Function/Transformation |
|---|---|---|
| Biotic Characteristics—Before and After Storm | ||
| % EPT | Quasi-Binomial | Logit |
| % Midge | Quasi-Binomial | Logit |
| Abiotic Characteristics—Before and After Storm | ||
| Bankfull Width | Gaussian | Log(y) |
| % Shade | Quasi-Binomial | Logit |
| Total Wood | Quasi-Poisson | None |
| Biotic Characteristics—After Storm | ||
| Abundance | Gaussian | Log(y) |
| Richness | Quasi-Poisson | Log |
| % EPT | Quasi-Binomial | Logit |
| % Midge | Quasi-Binomial | Logit |
| % Multi-voltine | Quasi-Binomial | Logit |
| % Scraper | Quasi-Binomial | Logit |
| Sediment Intolerant Species | Gaussian | Log(y) |
| Abiotic Characteristics—After Storm | ||
| Max Temperature | Gaussian | None |
| Min Temperature | Gaussian | None |
| Total Wood | Quasi-Poisson | Square Root |
| Bankfull Width | Gaussian | 1/y |
| Wet Width | Gaussian | None |
| % Fine Sediments | Quasi-Binomial | Logit |
| % Canopy Closure | Quasi-Binomial | Logit |
| Cluster 1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxa lowest level | Major group | RA | FFG | FSBI | Taxa lowest level | Major group | RA | FFG | FSBI | ||
| Group 1A | 45 | Dicosmoecus gilvipes | T | C | S | ||||||
| 1 | Atractides sp. | A | C | P | 46 | Synorthocladius sp. | C | C | G | ||
| 2 | Lebertia sp. | A | C | P | 47 | Antocha sp. | D | C | G | ES | |
| 3 | Micropsectra sp. | C | A | G | 48 | Cricotopus sp. | C | A | SH | ||
| 4 | Polypedilum sp. | C | A | SH | 49 | Eukiefferiella brehmi gr. | C | C | G | ||
| 5 | Sperchon sp. | A | C | P | 50 | Eukiefferiella pseudomontana gr. | C | C | G | ||
| 6 | Zapada cinctipes | P | A | SH | 51 | Paraperla sp. | P | R | P | ||
| 7 | Torrenticola sp. | A | C | P | 52 | Rhyacophila brunnea gr. | T | C | P | ||
| 8 | Oligochaeta | Oth | A | G | 53 | Hexatoma sp. | D | C | P | ||
| 9 | Dicranota sp. | D | C | P | 54 | Paraleptophlebia sp. | E | C | G | S | |
| 10 | Brillia sp. | C | C | SH | 55 | Lepidostoma sp. | T | R | SH | ||
| 11 | Tvetenia bavarica gr. | C | A | G | 56 | Krenosmittia sp. | C | C | G | ||
| 12 | Baetis tricaudatus | E | A | G | 57 | Rheocricotopus sp. | C | C | G | ||
| 13 | Cinygmula sp. | E | A | S | S | 58 | Yoraperla sp. | P | C | SH | S |
| 14 | Calineuria californica | P | A | P | S | 59 | Clinocera sp. | D | C | P | |
| 15 | Glossosoma sp. | T | A | S | S | 60 | Parapsyche almota | T | C | G | |
| 16 | Simulium sp. | D | A | F | 61 | Eukiefferiella gracei gr. | D | C | G | ||
| 17 | Sweltsa sp. | P | A | P | 62 | Rhyacophila hyalinata gr. | T | C | P | ES | |
| 18 | Nematoda | Oth | C | P | 63 | Eukiefferiella tirolensis | C | R | G | ||
| 19 | Rhyacophila betteni gr. | T | C | P | S | 64 | O. (Symp.) lignicola | C | R | G | |
| 20 | Thienemannimyia gr. sp. | C | C | G | 65 | Ironodes sp. | E | R | S | S | |
| 21 | Parametriocnemus sp. | C | C | G | Group 1B | ||||||
| 22 | Malenka sp. | P | C | SH | 66 | Dixa sp. | D | C | G | ||
| 23 | Optioservus sp. | Co | C | S | 67 | Rheotanytarsus sp. | C | C | F | ||
| 24 | Bezzia/Palpomyia sp. | D | C | P | 68 | Skwala sp. | P | C | P | ||
| 25 | Wormaldia sp. | T | A | F | 69 | Neoplasta sp. | D | R | S | ||
| 26 | Zaitzevia sp. | Co | C | g | 70 | Attenella delantala | E | C | G | ||
| 27 | Narpus sp. | Co | C | G | 71 | Hydropsyche sp. | T | C | F | ||
| 28 | Heterlimnius sp. | Co | A | G | S | 72 | Heleniella sp. | C | C | G | |
| 29 | Neophylax rickeri | T | C | S | S | 73 | Glutops sp. | D | C | P | |
| 30 | Drunella doddsii | E | C | S | VS | 74 | Arctopsyche grandis | T | C | F | |
| 31 | Epeorus sp. | E | A | S | VS | 75 | Pericoma/Telmatoscopus sp. | D | C | G | |
| 32 | Hesperoperla pacifica | P | C | P | VS | 76 | Zavrelimyia sp. | C | R | P | |
| 33 | Rhyacophila angelita gr. | T | C | P | VS | 77 | Protzia sp. | A | R | P | |
| 34 | Eukiefferiella claripennis gr. | C | C | G | 78 | Paraphaenocladius “n. sp.” | C | C | G | ||
| 35 | Thienemanniella sp. | C | C | G | 79 | Nilotanypus fimbriatus | C | R | P | ||
| 36 | Diphetor hageni | E | C | G | 80 | Sperchonopsis sp. | A | R | P | ||
| 37 | Rhithrogena sp. | E | C | S | ES | 81 | Pacifastacus leniusculus | Oth | R | SH | |
| 38 | Orthocladius Complex | C | A | G | 82 | Juga spp. | G | C | S | ||
| 39 | Stempellinella sp. | C | C | G | 83 | Pisiduim | Oth | C | F | ||
| 40 | Chelifera/Metachela sp. | D | C | U | 84 | Rhyacophila blarina | T | C | P | ||
| 41 | Serratella tibialis | E | C | G | S | 85 | Despaxia augusta | P | C | SH | |
| 42 | Corynoneura sp. | C | R | G | ES | 86 | Rhyacophila narvae | T | C | P | |
| 43 | Pagastia sp. | C | C | G | 87 | Rhyacophila vagrita gr. | T | C | P | ||
| 44 | Acentrella turbida | E | A | G | 88 | Atylotus/Tabanus sp. | D | R | G | ||
| Cluster 2 | |||||||||||
| Taxa lowest level | Major group | RA | FFG | FSBI | # | Taxa lowest level | Major group | RA | FFG | FSBI | |
| Group 2A | 171 | Allocosmoecus partitus | T | R | P | ||||||
| 89 | Tanytarsus sp. | C | C | F | 172 | Psychoglypha sp. | T | R | G | ||
| 90 | Agraylea sp. | T | R | SH | 173 | Polycelis sp. | Oth | R | P | ||
| 91 | Mideopsis sp. | A | R | P | 174 | Saetheria tylus | C | R | U | ||
| 92 | Micrasema sp. | T | R | SH | 175 | Cultus sp. | P | R | P | VS | |
| 93 | Centroptilum sp. | E | R | G | 176 | Baetis alius | E | R | G | ||
| 94 | Oreogeton sp. | D | R | P | 177 | Crenitis sp. | Co | C | P | ||
| 95 | Amiocentrus aspilus | T | R | SH | 178 | Nixe sp. | E | R | S | S | |
| 96 | Hydrobiidae | G | C | S | 179 | Brundiniella sp. | C | R | U | ||
| 97 | Boreochlus sp. | C | R | G | 180 | Attenella margarita | E | R | G | VS | |
| 98 | Rickera sorpta | P | R | P | 181 | Ochthebius sp. | Co | R | P | ||
| 99 | Eukiefferiella brevicalcar gr. | C | R | G | 182 | Sanfillipodytes sp. | Co | R | P | ||
| 100 | Potthastia gaedii gr. | C | C | G | 183 | Goera sp. | T | R | S | ||
| 101 | Paratanytarsus sp. | D | R | G | 184 | Tipula sp. | D | R | SH | ||
| 102 | Potthastia longimana gr. | C | R | G | 185 | Euryhapsis sp. | C | R | U | ||
| 103 | Oribatei | A | R | P | 186 | Parorthocladius sp. | C | R | G | ||
| 104 | Sublettea sp. | C | R | F | 187 | Psilometriocnemus sp. | C | R | G | ||
| 105 | Pteronarcys californica | P | R | SH | S | 188 | Testudacarus sp. | A | R | P | |
| 106 | Parakiefferiella sp. | C | R | G | 189 | Ulomorpha sp. | D | R | U | ||
| 107 | P. umbilicatellus | G | R | G | 190 | Stygothrombium sp. | A | R | P | ||
| 108 | Bryelmis rivularis | Co | R | G | 191 | Limnophila sp. | D | R | P | ||
| 109 | Pteronarcella sp. | P | R | SH | 192 | Frisonia picticeps | P | R | P | ||
| 110 | Stenochironomus sp. | C | R | G | 193 | Oreodytes sp. | Co | R | P | ||
| 111 | Ephemerella aurivillii | E | R | G | 194 | Perlinodes aurea | P | C | P | ES | |
| 112 | Ampumixis dispar | Co | R | F | ES | 195 | Rhyacophila arnaudi | T | R | P | |
| 113 | Cryptolabis sp. | D | R | SH | 196 | Rhyacophila pellisa/valuma | T | R | P | ||
| 114 | Labiobaetis sp. | E | R | S | 197 | Atherix sp. | D | C | P | ||
| 115 | Meringodixa sp. | D | R | G | 198 | Turbellaria | Oth | R | P | ||
| 116 | Ptychoptera sp. | D | R | G | 199 | Baetis flavistriga | E | C | G | ||
| 117 | Hydrobaenus sp. | C | R | S | 200 | Ecdyonurus sp. | E | R | S | ||
| 118 | Sialis sp. | M | R | P | 201 | Hesperoconopa sp. | D | R | G | VS | |
| 119 | Maruina sp. | D | R | S | 202 | Lopescladius sp. | C | C | G | ||
| 120 | Psychomyia sp. | T | R | CF | 203 | Demicryptochironomus sp. | C | R | G | ||
| 121 | Cleptelmis addenda | Co | R | B | 204 | Drunella grandis | E | C | S | S | |
| 122 | Cinygma sp. | E | R | S | 205 | Onocosmoecus sp. | T | R | SH | ||
| 123 | Atrichopogon sp. | D | R | B | VS | 206 | Chrysops sp. | D | R | SH | |
| 124 | Larsia sp. | C | R | P | 207 | Pentaneura sp. | C | R | P | ||
| Group 2B–1 | 208 | Hydraena sp. | Co | R | P | ||||||
| 125 | Eukiefferiella devonica gr. | C | R | G | 209 | Fluminicola sp. | G | C | S | ||
| 126 | O. (Euortho.) rivicola gr. | C | C | G | 210 | Brachycentrus occidentalis | T | R | G | ||
| 127 | Oligophlebodes sp. | T | R | S | 211 | Krenopelopia sp. | C | R | P | ||
| 128 | Ephemerella sp. | E | R | S | Group 2B–2 | ||||||
| 129 | Doroneuria sp. | P | R | P | VS | 212 | Pedicia sp. | D | R | P | |
| 130 | Diamesa sp. | C | R | G | 213 | Psephenus sp. | Co | R | S | ||
| 131 | Agabus sp. | Co | R | P | 214 | Lara sp. | C | R | SH | ||
| 132 | Timpanoga hecuba | E | R | G | 215 | Moselia infuscata | P | R | SH | ||
| 133 | Limnesia | A | R | P | 216 | Cladopelma sp. | C | R | G | ||
| 134 | Microtendipes rydalensis gr. | C | R | F | 217 | Pseudocloeon sp. | E | R | SH | ||
| 135 | Nilobezzia sp. | D | R | P | 218 | Natarsia sp. | C | R | P | ||
| 136 | Tokunagaia sp. | C | C | G | 219 | Pseudosmittia sp. | C | R | G | ||
| 137 | Caudatella sp. | E | R | G | VS | 220 | Wandesia sp. | A | R | P | |
| 138 | Gonomyia sp. | D | R | G | 221 | O. (Euortho.) rivulorum gr. | C | R | G | ||
| 139 | Zapada frigida | P | R | SH | S | 222 | Dixella sp. | D | R | G | |
| 140 | Drunella spinifera | E | R | S | S | 223 | Taeniopterygidae | P | R | SH | |
| 141 | Suwallia sp. | P | R | P | S | 224 | Matriella teresa | E | R | SH | |
| 142 | Megarcys sp. | P | R | P | ES | 225 | Ablabesmyia sp. | C | R | P | |
| 143 | Zapada columbiana | P | R | SH | 226 | Tribelos jucundum | C | R | G | ||
| 144 | Chaetocladius sp. | C | R | G | 227 | Valvata sp. | G | R | S | ||
| 145 | Parorthocladius sp. | C | R | G | 228 | Paracladopelma sp. | C | R | G | ||
| 146 | Reomyia sp. | C | R | G | 229 | Cladotanytarsus sp. | C | R | G | ||
| 147 | Diplocladius sp. | C | C | G | 230 | Muscidae | D | R | P | ||
| 148 | Parochlus sp. | C | C | G | 231 | Boreoheptagyia sp. | C | R | G | ||
| 149 | Cardiocladius sp. | C | R | G | 232 | Baetis notos | E | R | G | ||
| 150 | Isoperla sp. | P | R | P | 233 | Serratella micheneri | E | R | G | ||
| 151 | Ametor sp. | Co | R | U | 234 | Parachaetocladius sp. | C | R | G | S | |
| 152 | Serratella teresa | E | R | G | S | 235 | Gomphidae | M | R | G | |
| 153 | Aturus sp. | A | R | P | 236 | Mesobates sp. | A | R | P | ||
| 154 | Setvena sp. | P | R | P | 237 | Stilobezzia sp. | D | R | P | ||
| 155 | Neophylax splendens | T | R | S | S | 238 | Goera archaon | T | R | S | |
| 156 | Zapada oregonensis gr. | P | R | SH | S | 239 | Ptychoptera sp. | D | R | u | |
| 157 | Pteronarcys princeps | P | R | SH | S | 240 | Metriocnemus sp. | C | R | G | |
| 158 | Hygrobates sp. | A | R | P | 241 | Labrundinia sp. | C | R | P | ||
| 159 | Heterotrissocladius marcidus gr. | C | R | G | 242 | Forcipomyia sp. | D | R | G | ||
| 160 | Constempellina sp. | C | R | G | 243 | Phaenopsectra sp. | C | R | S | ||
| 161 | Hydatophylax hesperus | T | R | SH | 244 | Anafroptilum sp. | E | R | G | ||
| 162 | Capniidae | P | C | SH | 245 | Laccobius sp. | Co | R | SH | ||
| 163 | Rhabdomastix fascigera gr. | D | R | G | S | 246 | Paratendipes sp. | C | R | G | |
| 164 | Apatania sp. | T | C | S | S | 247 | Pseudodiamesa sp. | C | R | G | |
| 165 | Cryptochia sp. | T | R | SH | 248 | Agapetus sp. | T | R | S | ||
| 166 | Ameletus sp. | E | C | G | 249 | Limnophyes sp. | C | R | B | S | |
| 167 | Drunella coloradensis/flavilinea | E | C | S | S | 250 | Limonia sp. | D | R | SH | |
| 168 | Trichoclinocera sp. | D | R | P | 251 | Nanocladius sp. | C | R | G | ||
| 169 | Rhyacophila vofixa gr. | T | C | P | VS | 252 | Eukiefferiella coerulescens gr. | C | R | B | |
| 170 | Kogotus sp. | P | R | P | VS | 253 | Paraboreochlus sp. | C | R | G |
| Cluster | # Species | % Total Abundance | % Total Biomass | % Rare Species in Cluster |
|---|---|---|---|---|
| 1a | 65 | 87 | 97.4 | 9 |
| 1b | 22 | 10 | 1.2 | 32 |
| 2a | 40 | 2 | 1.4 | 82 |
| 2b | 141 | 0.03 | <0.1 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danehy, R.J.; Bilby, R.E.; Justice, T.E.; Lester, G.T.; Jones, J.E.; Haddadi, S.S.; Merritt, G.D. Aquatic Biological Diversity Responses to Flood Disturbance and Forest Management in Small, Forested Watersheds. Water 2021, 13, 2793. https://0-doi-org.brum.beds.ac.uk/10.3390/w13192793
Danehy RJ, Bilby RE, Justice TE, Lester GT, Jones JE, Haddadi SS, Merritt GD. Aquatic Biological Diversity Responses to Flood Disturbance and Forest Management in Small, Forested Watersheds. Water. 2021; 13(19):2793. https://0-doi-org.brum.beds.ac.uk/10.3390/w13192793
Chicago/Turabian StyleDanehy, Robert J., Robert E. Bilby, Tiffany E. Justice, Gary T. Lester, Jay E. Jones, Sogal S. Haddadi, and Glenn D. Merritt. 2021. "Aquatic Biological Diversity Responses to Flood Disturbance and Forest Management in Small, Forested Watersheds" Water 13, no. 19: 2793. https://0-doi-org.brum.beds.ac.uk/10.3390/w13192793