
Identification of Outlier Loci Responding to Anthropogenic and Natural Selection Pressure in Stream Insects Based on a Self-Organizing Map

Abstract
:
1. Introduction
2. Materials and Methods
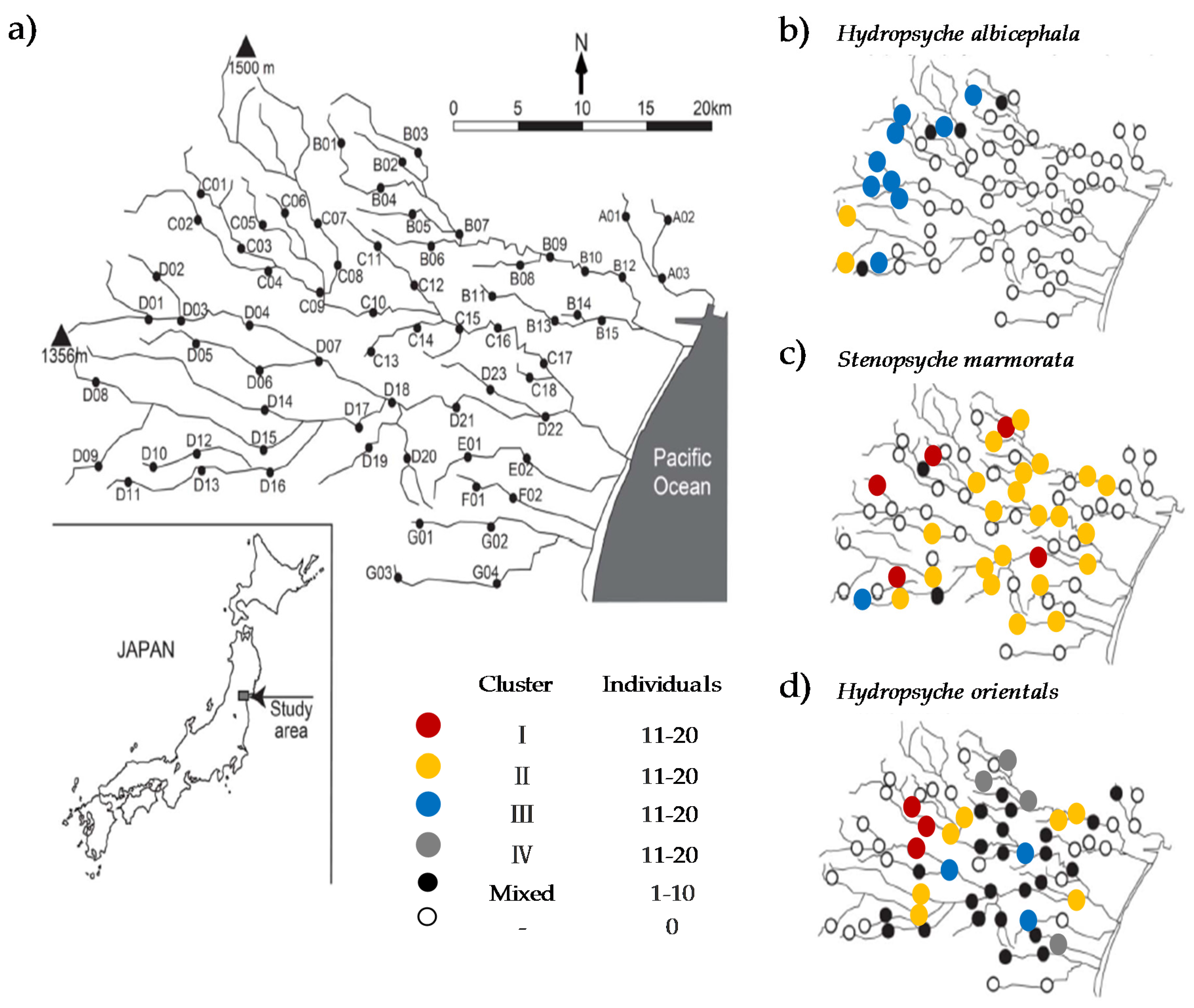
2.1. Ecological and Genetic Data
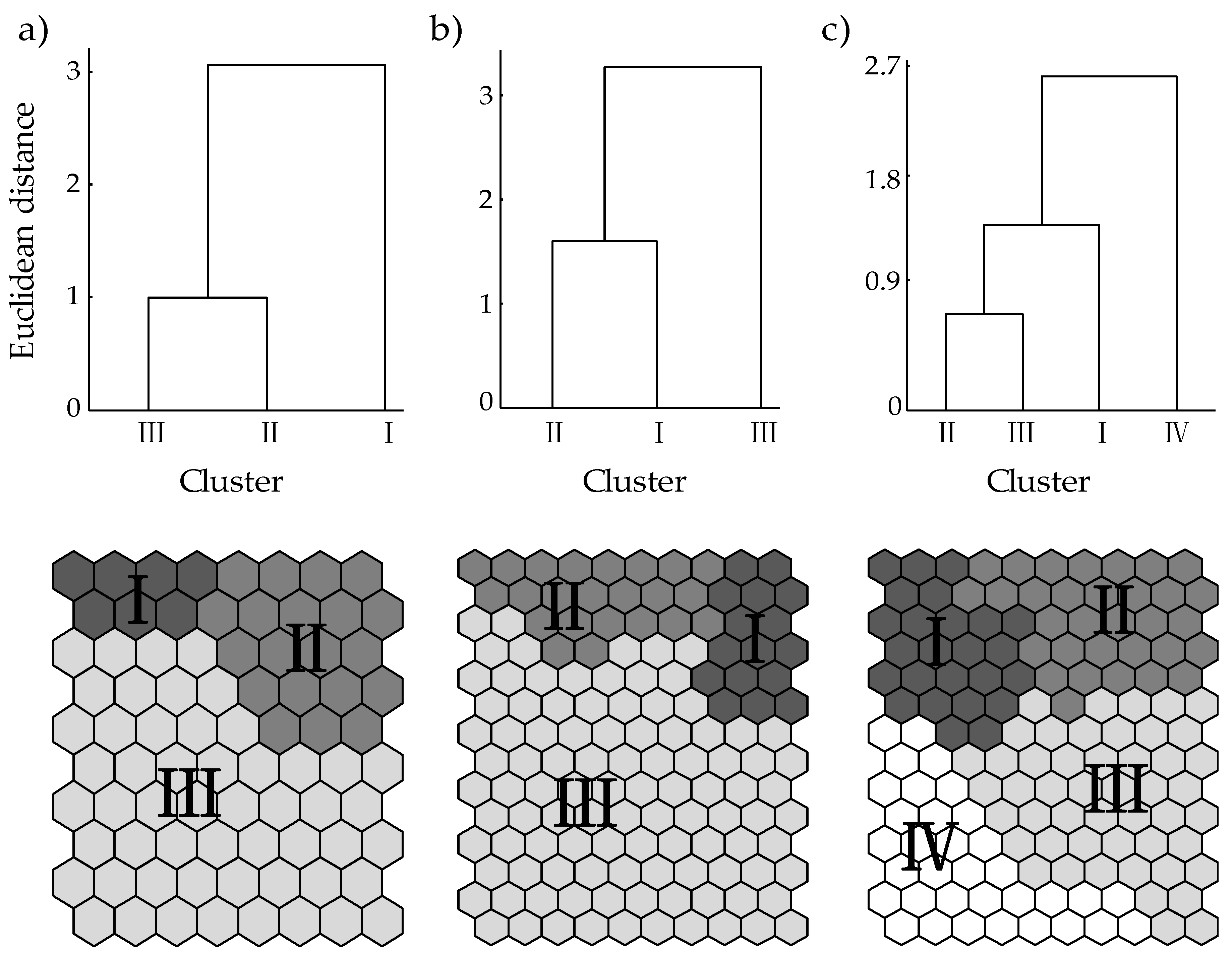
2.2. SOM Applied to Environmental and AFLP Data
2.3. Screening Outlier Loci and Environmental Responsiveness
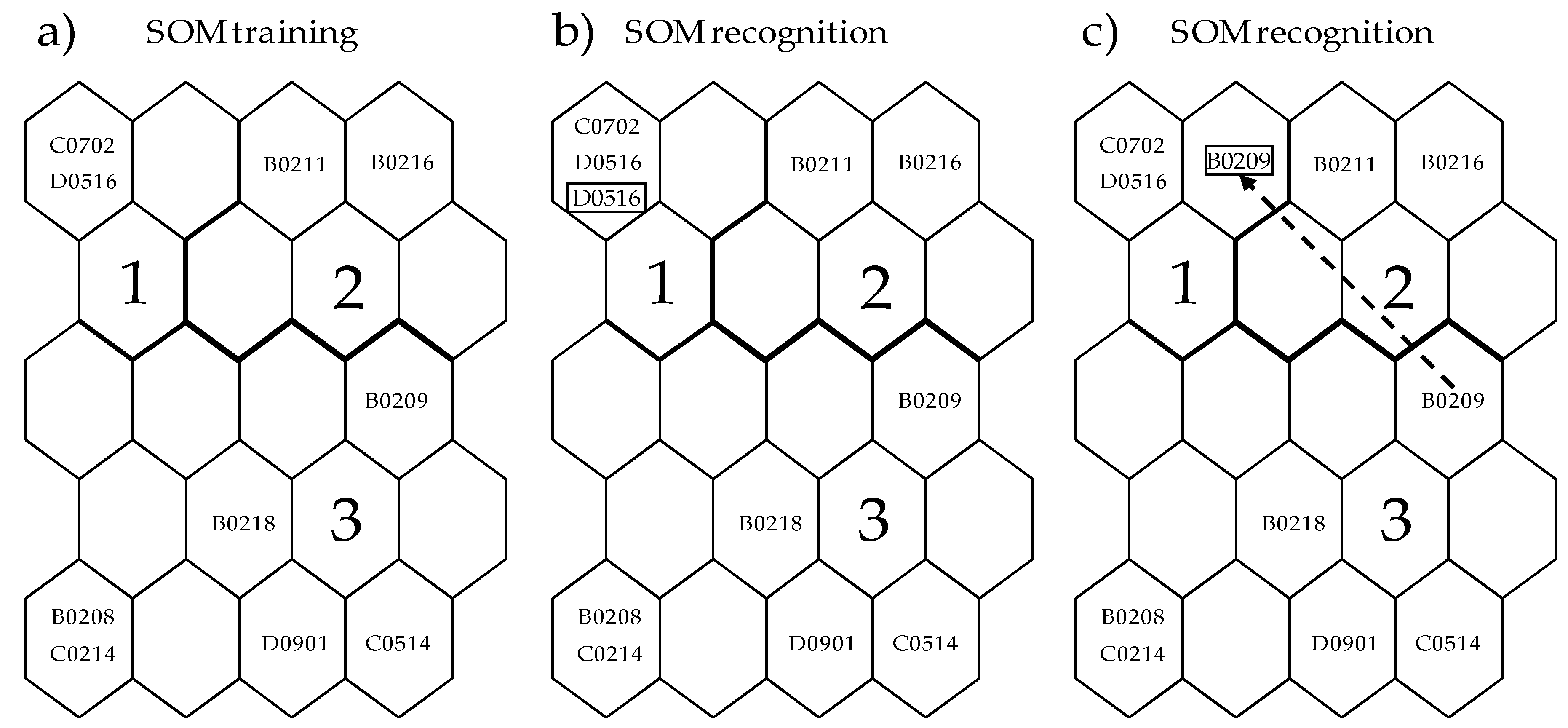
- SOM is trained with the AFLP data and the trained SOM output units are classified to clusters (I, II, …, N) (see Section 2.2).
- A vector, B (list of clusters with training data for each individual) is produced (e.g., B = (I, II, I, III) with the first, second, third, and fourth individual matching cluster I, II, I, and III, respectively). Euclidian distance is calculated between clusters.
- Each locus is altered by flipping over (switching either “presence to absence” or “absence to presence”) separately for each individual.
- Sensitivity analysis is conducted with altered datasets through recognition (See Figure A1).
- A vector, G (list of clusters with altered data) is produced (e.g., G = (I, II, I, I) with each individual sequentially belonging to I, II, I, and I, similar to the case of B in process 2).
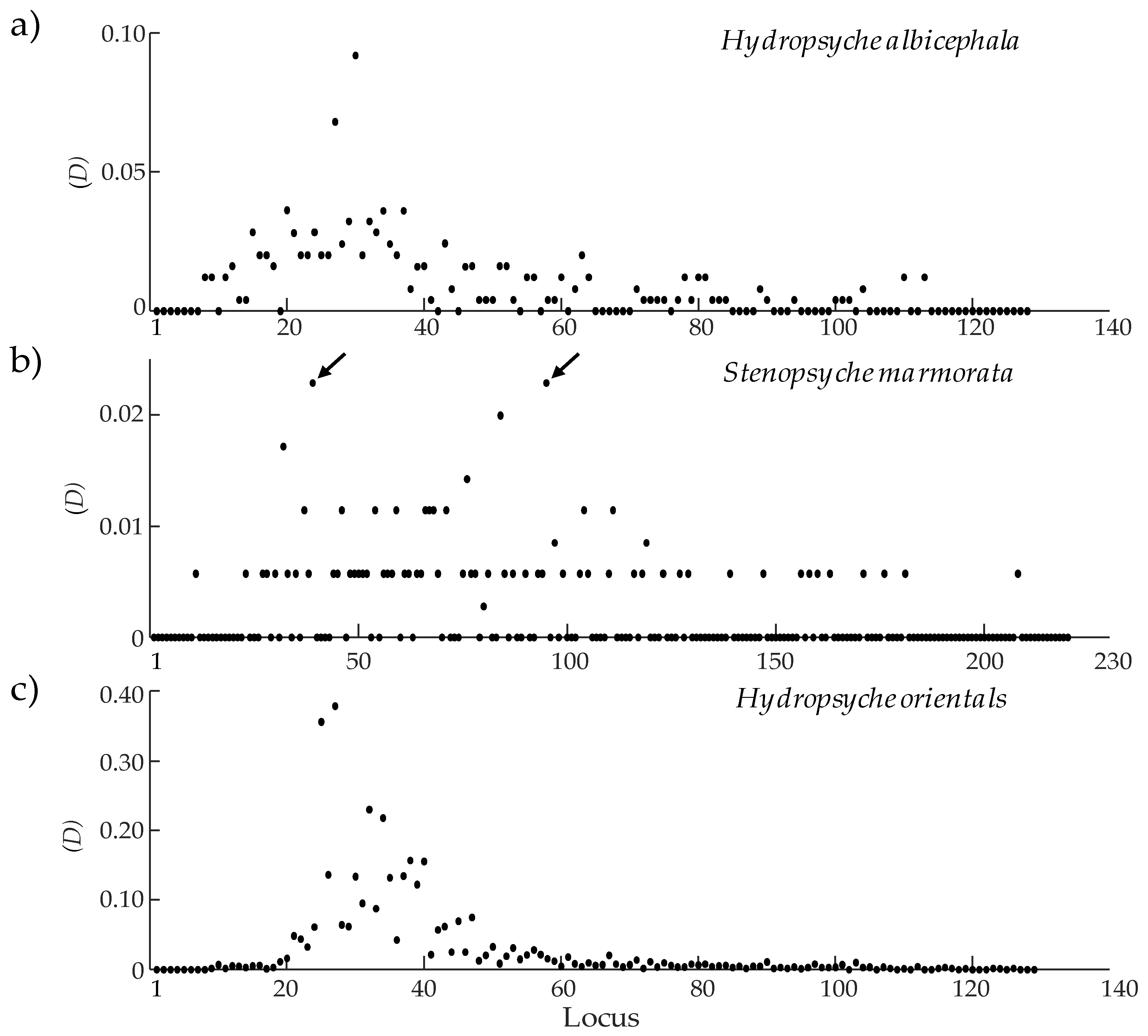
- Mean cluster distance for each locus (D) is defined to determine the overall differences between training and recognition for individuals. D is calculated as average of the summed Euclidian distance according to B compared with G. If the change crossed over clusters with higher distance, higher values of distance would be given to this individual.
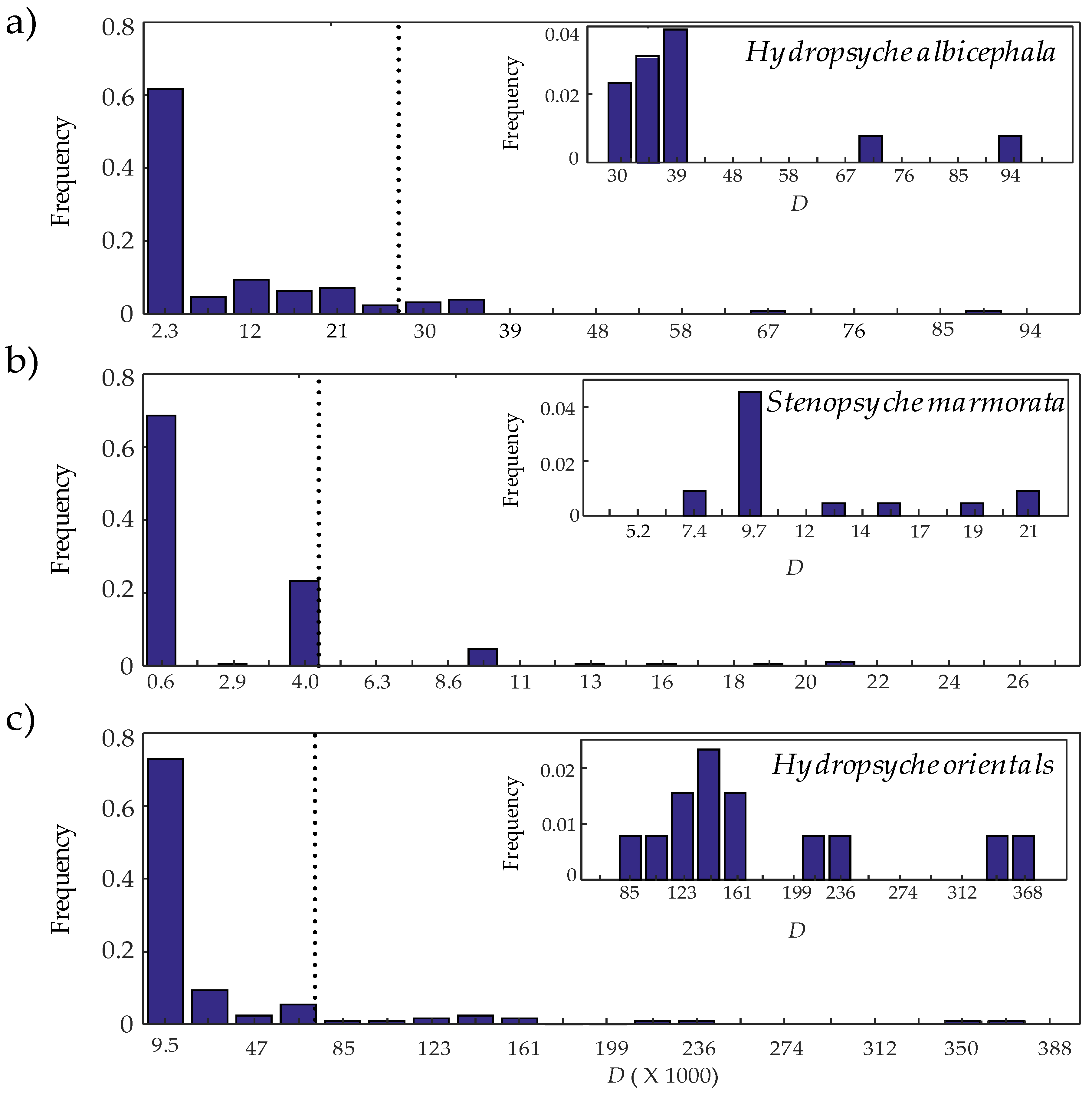
- According to D outlier loci are determined. Loci with D value higher than the 90th percentile [40] were considered as outliers under selection in this study.
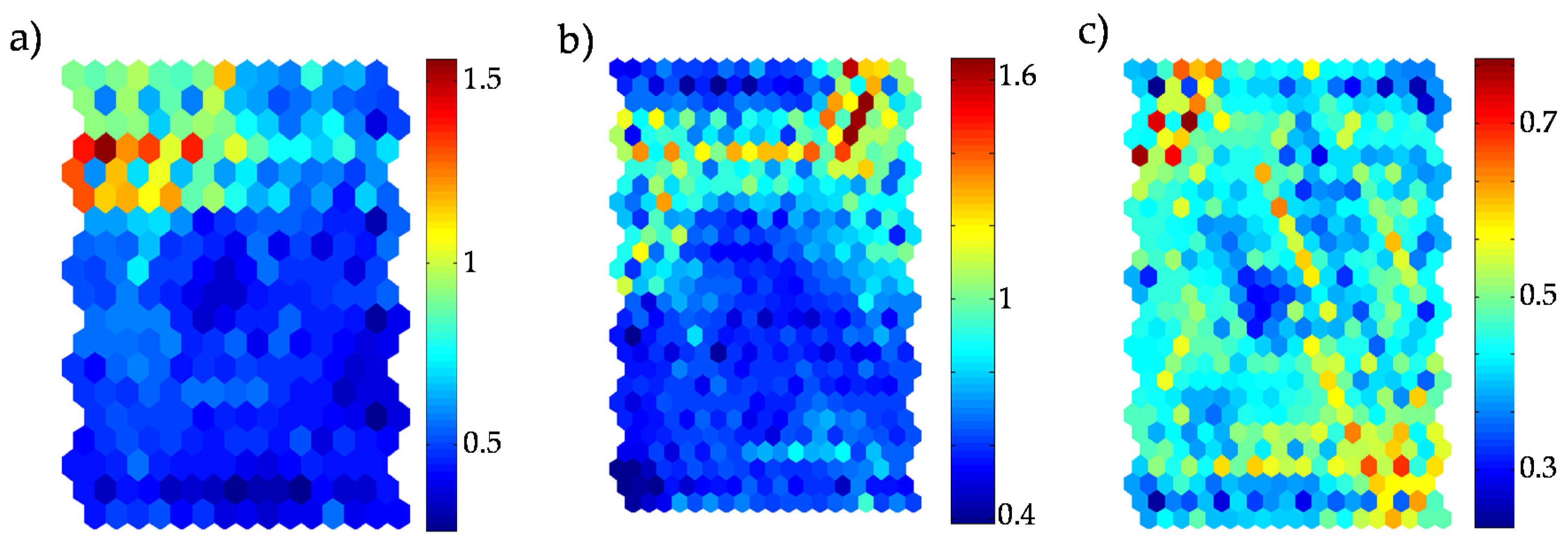
- Once outlier loci were identified, we examined their relationships with each environmental variable. Indices Ek,i and Ek were devised to present responsiveness of each environmental variable (k) in each cluster (i) and overall responsiveness across clusters, respectively, after SOM recognition as follows:where ek,i is the mean of environmental variable k for all individuals belonging to cluster i before data alteration, hk,i is the mean of the same environmental variable k for all individuals belonging to cluster i after recognition, and N is the total number of clusters. Higher Ek,i value indicates a higher potential of variation in specific environmental factor, k, due to loci alteration between training and recognition in cluster i, whereas a higher Ek value represents overall responsiveness across all clusters on the SOM.
- 9.
- Based on outliers according to D (process 7), a locus-specific pattern showing degree of associations between outlier loci and environmental factors was determined according to the level of Ek’ (process 8) (see Figure 9 for details).
3. Results
3.1. Habitat Specialization
3.2. Patterning of Loci Data
3.3. Identifying Outlier Loci and Environmental Responsiveness
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SOM | self-organizing map |
| AFLP | Amplified Fragment Length Polymorphism |
| QTL | quantitative trait loci |
| BCPOM | Benthic Coarse Particulate Organic Matter |
| SFPOM | Suspended Fine Particulate Organic Matter |
| Chl-a | Chlorophyll-a |
| BOD | Biochemical Oxygen Demand |
| SS | Suspended Solid |
| NOX-N | nitrite/nitrate nitrogen |
| NH4-N | ammonia nitrogen |
| PO4-P | orthophosphate phosphorus |
| K-S test | Kolmogorov-Smirnoff test |
| FST | Wright’s fixation index |
Appendix




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
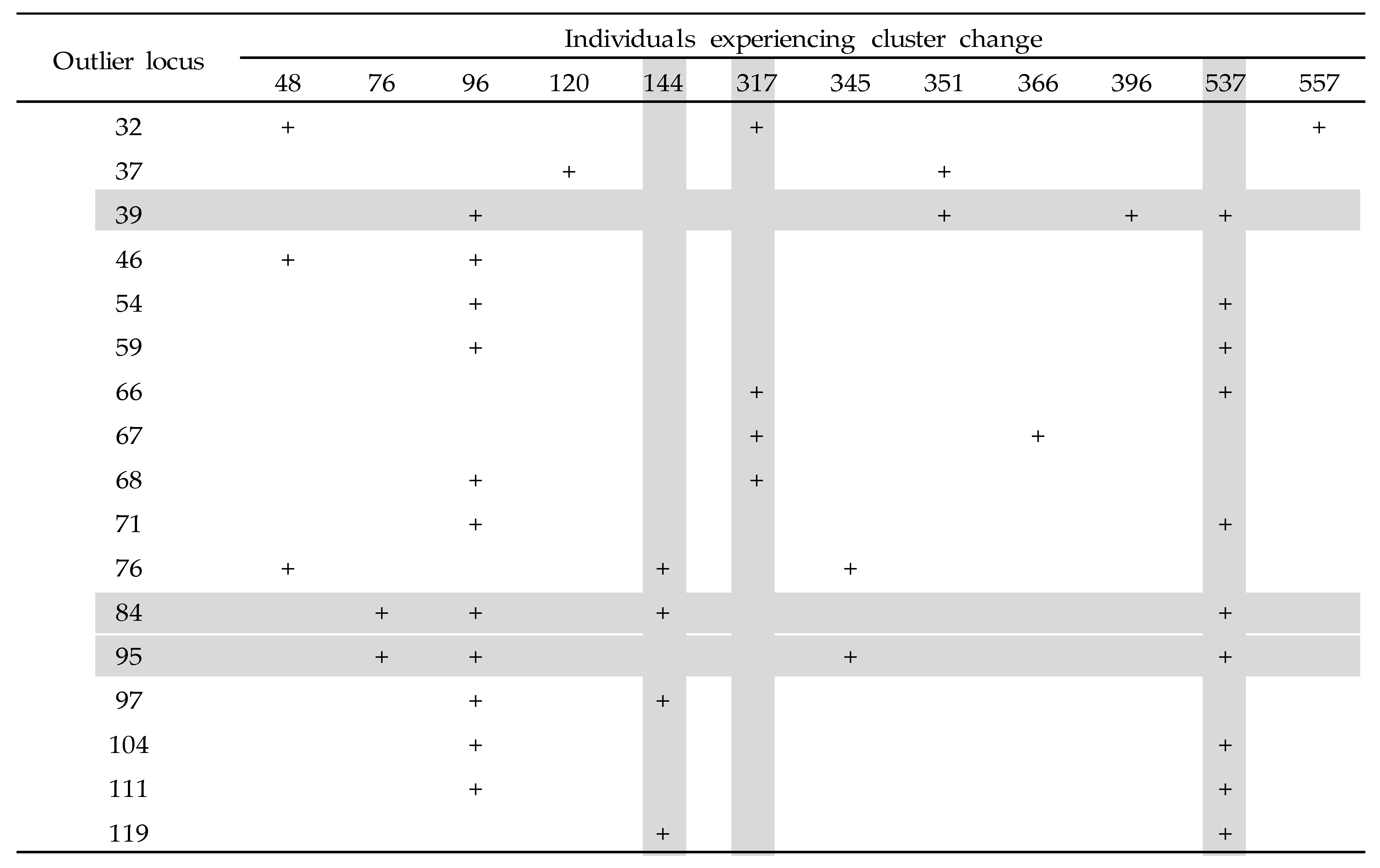
| Species | Dfdist vs. BayeScan | Dfdist vs. SOM | BayeScan vs. SOM | Common Outliers |
|---|---|---|---|---|
| Hydropsyche albicephala | 4, 5, 11, 15, 29, 43 | 15, 29, 43 | 15, 29, 33, 35, 43 | 15, 29, 43 |
| Stenopsyche marmorata | 32, 33, 34, 43, 45, 56, 66, 67, 68, 70, 76, 78, 80, 84, 85, 95, 97, 104, 106, 111, 116 | 32, 66, 67,68, 76,84, 95, 97, 104, 111 | 32, 37, 39, 46, 54, 59, 66, 67, 68, 71, 76, 84, 95, 97, 104, 111 | 32, 66, 67, 68, 76, 84, 95, 97, 104, 111 |
| Hydropsyche orientals | 20, 25, 27, 39, 40, 47, 49, 53, 67 | 25, 27, 39, 40 | 25, 26, 27, 32, 39, 40 | 25, 27, 39, 40 |

References
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P. The urban stream syndrome: Current knowledge and the search for a cure. J. North Am. Benthol. Soc. 2005, 3, 706–723. [Google Scholar] [CrossRef]
- Hellawell, J.M. Biological indicators. In Biological Indicators of Freshwater Pollution and Environmental Management, 1st ed.; Hellawell, J.M., Ed.; Elsevier Science Publishing Co., Inc: New York, NY, USA, 1986; pp. 45–63. [Google Scholar]
- Rosenberg, D.M.; Resh, V.H. Introduction to Freshwater Biomonitoring and Benthic Macroinvertebrates. In FreshwaterBiomonitoring and Benthic Macroinvertebrates, 1st ed.; Rosenberg, D.M., Resh, V.H., Eds.; Chapman & Hall: New York, NY, USA, 1993; pp. 1–30. [Google Scholar]
- Wright, J.F. An introduction to RIVPACS. In Assessing the Biological Quality of Freshwaters, RIVPACS and Other Techniques; Wright, J.F., David, W.S., Mike, T.F., Eds.; Freshwater Biological Association: Ambleside, UK, 2000; pp. 5–26. [Google Scholar]
- Park, Y.S.; Song, M.Y.; Park, Y.C.; Oh, K.H.; Cho, E.C.; Chon, T.S. Community patterns of benthic macroinvertebrates collected on the national scale in Korea. Ecol. Model. 2007, 203, 26–33. [Google Scholar] [CrossRef]
- Storz, J.F. Invited Review: Using genome scans of DNA polymorphism to infer adaptive population divergence. Mol. Ecol. 2005, 14, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Kazama, S.; Omura, T.; Michael, T.M. Adaptive Genetic Divergence along Narrow Environmental Gradients in Four Stream Insects. PLoS ONE 2014, 9, e93055. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, T. An integrated approach of comparative genomics and heritability analysis of pig and human on obesity trait: Evidence for candidate genes on human chromosome 2. BMC Genom. 2012, 13, 711. [Google Scholar] [CrossRef] [PubMed]
- Tabor, H.K.; Risch, N.J.; Myers, R.M. Candidate-gene approaches for studying complex genetic traits: Practical considerations. Nat. Rev. Genet. 2002, 3, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.; Afonso, S.; Geraldes, A.; Garreau, H.; Bolet, G.; Boucher, S.; Tircazes, A.; Queney, G.; Nachman, M.W.; Ferrand, N. The genetic structure of domestic rabbits. Mol. Biol. Evol. 2011, 28, 1801–1816. [Google Scholar]
- Daniels, S.E.; Bhattacharrya, S.; James, A.; Leaves, N.I.; Young, A.; Hill, M.R.; Faux, J.A.; Ryan, G.F.; le Souef, P.N.; Lathrop, G.M.; et al. A genome-wide search for quantitative trait loci underlying asthma. Nature 1996, 383, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, K.J.F.; Vanhala, T.K.; Biere, A.; Nevo, E.; van Damme, J.M. The genetic basis of adaptive population differentiation: A quantitative trait locus analysis of fitness traits in two wild barley populations from contrasting habitats. Evolution 2004, 58, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.M.; Bernatchez, L. Integrating QTL mapping and genome scans towards the characterization of candidate loci under parallel selection in the lake whitefish (Coregonus clupeaformis). Mol Ecol. 2005, 14, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.A.; Nichols, R.A. Evaluating loci for use in the genetic analysis of population structure. Proc. Roy. Soc. B Biol. Sci. 1996, 263, 1619–1626. [Google Scholar] [CrossRef]
- Beaumont, M.A.; Balding, D.J. Identifying adaptive genetic divergence among populations from genome scans. Mol Ecol. 2004, 13, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Foll, M.; Gaggiotti, O.E. Identifying the environmental factors that determine the genetic structure of populations. Genetics 2006, 74, 875–891. [Google Scholar] [CrossRef] [PubMed]
- Foll, M.; Gaggiotti, O.E. Estimating selection with different markers and varying demographic scenarios: A Bayesian perspective. Genetics 2008, 180, 977–993. [Google Scholar] [CrossRef] [PubMed]
- Strachan, T.; Read, A. Genes in Pedigrees and Populations. In Human Molecular Genetics; Garland Science: New York, NY, USA, 2009. [Google Scholar]
- Lotterhos, K.E.; Whitlock, M.C. Evaluation of demographic history and neutral parameterization on the performance of FST outlier tests. Mol. Ecol. 2014, 23, 2178–2192. [Google Scholar] [CrossRef] [PubMed]
- Straškraba, M. Ecotechnological models for reservoir water quality management. Ecol. Model. 1994, 74, 1–38. [Google Scholar] [CrossRef]
- Hamilton, G.; McVinish, R.; Mengersen, K. Bayesian model averaging for harmful algal bloom prediction. Ecol. Appl. 2009, 19, 1805–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornarelli, R.; Galelli, S.; Castelletti, A.; Antenucci, J.P.; Marti, C.L. An empirical modeling approach to predict and understand phytoplankton dynamics in a reservoir affected by interbasin water transfers. Water Resour. Res. 2013, 49, 3626–3641. [Google Scholar] [CrossRef]
- Kohonen, T. The self-organizing map. Proc. IEEE 1990, 78, 1464–1480. [Google Scholar] [CrossRef]
- Park, Y.-S.; Tison, J.; Lek, S.; Coste, M.; Giraudel, J.; Delmas, F. Application of a self-organizing map in ecological informatics: Selection of representative species from large community dataset. Ecol. Inform. 2006, 1, 247–257. [Google Scholar] [CrossRef]
- Chon, T.-S. Self-Organizing Maps applied to ecological sciences. Ecol. Inform. 2011, 6, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Ferrán, E.A.; Ferrara, P. Clustering proteins into families using artificial neural networks. Bioinformatics 1992, 8, 39–44. [Google Scholar]
- Nikkilä, J.; Törönen, P.; Kaski, S.; Venna, J.; Castrén, E.; Wong, G. Analysis and visualization of gene expression data using Self-Organizing Maps. Neural Netw. 2002, 15, 953–966. [Google Scholar] [CrossRef]
- Chavez-Alvarez, R.; Chavoya, A.; Mendez-Vazquez, A. Discovery of Possible Gene Relationships through the Application of Self-Organizing Maps to DNA Microarray Databases. PLoS ONE 2014, 9, e93233. [Google Scholar]
- Giraudel, J.L.; Aurelle, D.; Berrebi, P.; Lek, S. Application of the self-organizing mapping and fuzzy clustering to microsatellite data: How to detect genetic structure in brown trout (Salmo trutta) populations. In Artificial Neuronal Networks; Lek, S., Guégan, J.F., Eds.; Springer: New York, NY, USA, 2000; pp. 187–202. [Google Scholar]
- Kontunen-Soppela, S.; Parviainen, J.; Ruhanen, H.; Brosché, M.; Keinäen, M.; Thakur, R.C.; Kolehmainen, M.; Kangasjävi, J.; Oksanen, E.; Karnosky, D.F.; et al. Gene expression responses of paper birch (Betula papyrifera) to elevated CO2 and O3 during leaf maturation and senescence. Environ. Pollut. 2010, 158, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Z.; Jang, M.; Reggia, J.A. Effects of Varying Parameters on Properties of Self-Organizing Feature Maps. Neural Process. Lett. 1996, 4, 53–59. [Google Scholar] [CrossRef]
- Gonzalez, A.I.; Grana, M.; Anjou, A.D.; Albizuri, F.X.; Cottrell, M. A sensitivity analysis of the self-organizing maps as an adaptive one-pass non-stationary clustering algorithm: The case of color quantization of image sequences. Neural Processing Lett. 1997, 6, 77–89. [Google Scholar] [CrossRef]
- Paini, D.R.; Worner, S.P.; Cook, D.C.; De Barro, P.J.; Thomas, M.B. Using a self-organizing map to predict invasive species: Sensitivity to data errors and a comparison with expert opinion. J. Appl. Ecol. 2010, 47, 290–298. [Google Scholar] [CrossRef]
- Lek, S.; Guégan, J.F. Artificial Neuronal Networks: Application to Ecology and Evolution; Springer: Berlin, Germany, 2000. [Google Scholar]
- Park, Y.-S.; Céréghino, R.; Compin, A.; Lek, S. Applications of artificial neural networks for patterning and predicting aquatic insect species richness in running waters. Ecol. Model. 2003, 160, 265–280. [Google Scholar]
- Vesanto, J.; Alhoniemi, E. Clustering of the self-organizing map. Neural Netw. 2000, 11, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Wishart, D. An algorithm for hierarchical classifications. Biometrics 1969, 25, 165–170. [Google Scholar] [CrossRef]
- Kolmogorov, A. Sulla determinazione empirica di una legge di distribuzione. Giornale dell’Istituto Italiano degli Attuari 1933, 4, 83–91. [Google Scholar]
- Marsaglia, G.; Tsang, W.W.; Wang, J. Evaluating Kolmogorov’s Distribution. J. Stat. Softw. 2003, 8, 1–4. [Google Scholar] [CrossRef]
- Parkhomenko, E.; Tritchler, D.; Lemire, M.; Pingzhao, H.; Beyene, J. Using a higher criticism statistic to detect modest effects in a genome-wide study of rheumatoid arthritis. BMC Proceed. 2009, S40, 1–4. [Google Scholar] [CrossRef]
- Jaccard, P. The distribution of the flora in the alpine zone. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Chapman, D.; Kimstach, V. Selection of water quality variables. In Water Quality Assessments-A Guide to Use of Biota, Sediments and Water in Environmental Monitoring, 2nd ed.; Chapman, D., Ed.; E&FN Spon: London, UK, 1996; pp. 90–100. [Google Scholar]
- Bonin, A.; Miaud, C.; Taberlet, P.; Pompanon, F. Explorative genome scan to detect candidate loci for adaptation along a gradient of altitude in the common frog (Ranatemporaria). Mol. Biol. Evol. 2006, 23, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.C.; Foll, M.; Excoffier, L.; Heckel, G. Enhanced AFLP genome scans detect local adaptation in high-altitude populations of a small rodent (Microtusarvalis). Mol. Ecol. 2011, 20, 1450–1462. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Monaghan, M.T.; Takemon, Y.; Omura, T. Dispersal ability determines the genetic effects of habitat fragmentation caused by reservoirs in three species of aquatic insect. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 574–579. [Google Scholar] [CrossRef]









| Variables | Mean ± SD | Minimum | Maximum |
|---|---|---|---|
| Altitude (m) | 180.33 ± 142.36 | 2 | 590 |
| Stream order | 2.44 ± 1.17 | 1 | 5 |
| Width (m) | 7.83 ± 7.87 | 1.24 | 38.33 |
| Velocity (m·s−1) | 0.55 ± 0.23 | 0.03 | 1.28 |
| Mean gravel size (cm) | 12.71 ± 4.21 | 4.81 | 23.03 |
| Sediment (mm) | 10.43 ± 4.45 | 0.50 | 19 |
| Epilithon (mg Chl-a·cm−2) | 0.0016 ± 0.0024 a | 0.0001 a | 0.01 |
| BCPOM (mg AFDM·m−2) | 10.20 ± 9.58 | 0.89 | 54.07 |
| SFPOM (mg AFDM·L−1) | 0.0069 ± 0.0052 a | 0.0009 a | 0.02 |
| Species | No. of Outlier Loci | Dfdist vs. BayeScan | Dfdist vs. SOM | BayeScan vs. SOM | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BayeScan | Dfdist | SOM | Common Outliers | Similarity Index | No. of Common Outliers | Similarity Index | No. of Common Outliers | Similarity Index | No. of Common Outliers | |
| Hydropsyche albicephala | 16 | 7 | 14 | 3 | 0.35 | 6 | 0.17 | 3 | 0.2 | 5 |
| Stenopsyche marmorata | 56 | 23 | 17 | 10 | 0.36 | 21 | 0.34 | 10 | 0.28 | 16 |
| Hydropsyche orientals | 31 | 9 | 12 | 4 | 0.29 | 9 | 0.22 | 4 | 0.16 | 6 |
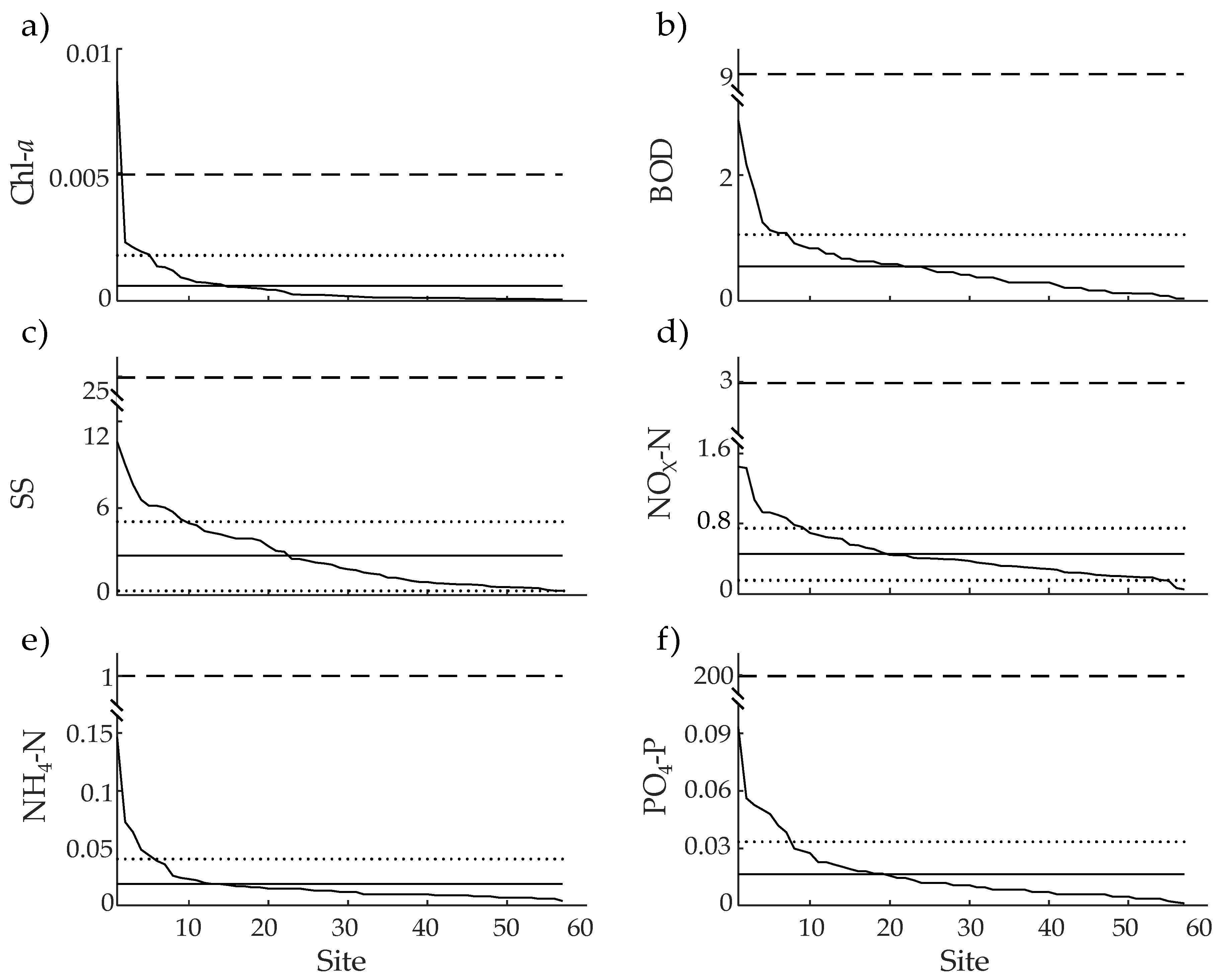
| Variables | Unit | Mean ± SD | Observed Values | WHO Standards a | Comparison (%) b | ||
|---|---|---|---|---|---|---|---|
| Minimum | Maximum | Minimum | Maximum | ||||
| Chl-a | mg·L−1 | 0.000,6 ± 0.001,2 | 0.0001 | 0.0087 | <0.0025 | 0.005,0–0.140,0 c | 12.000,0 |
| BOD | mg·L−1 | 0.455 ± 0.430 | 0.035 | 2.394 | 2 or less than 2 | 9 | 5.057 |
| SS | mg·L−1 | 3.004 ± 2.653 | 0.318 | 11.756 | – | 25 | 12.016 |
| NOX–N | mg·L−1 | 0.475 ± 0.313 | 0.055 | 1.513 | <0.1 | 3 | 15.817 |
| NH4–N | mg·L−1 | 0.019 ± 0.022 | 0.004 | 0.147 | 0.04 | 1 | 1.880 |
| PO4–P | mg·L−1 | 0.013,4 ± 0.014 | 0.001 | 0.078 | 0.001 | 200 | 0.007 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Watanabe, K.; Kim, D.-H.; Lee, S.-B.; Heo, M.; Kim, H.-S.; Chon, T.-S. Identification of Outlier Loci Responding to Anthropogenic and Natural Selection Pressure in Stream Insects Based on a Self-Organizing Map. Water 2016, 8, 188. https://0-doi-org.brum.beds.ac.uk/10.3390/w8050188
Li B, Watanabe K, Kim D-H, Lee S-B, Heo M, Kim H-S, Chon T-S. Identification of Outlier Loci Responding to Anthropogenic and Natural Selection Pressure in Stream Insects Based on a Self-Organizing Map. Water. 2016; 8(5):188. https://0-doi-org.brum.beds.ac.uk/10.3390/w8050188
Chicago/Turabian StyleLi, Bin, Kozo Watanabe, Dong-Hwan Kim, Sang-Bin Lee, Muyoung Heo, Heui-Soo Kim, and Tae-Soo Chon. 2016. "Identification of Outlier Loci Responding to Anthropogenic and Natural Selection Pressure in Stream Insects Based on a Self-Organizing Map" Water 8, no. 5: 188. https://0-doi-org.brum.beds.ac.uk/10.3390/w8050188