
Identification and expression analysis of cytokinin metabolic genes IPTs, CYP735A and CKXs in the biofuel plant Jatropha curcas
- Published
- Accepted
- Received
- Academic Editor
- Ivo Feussner
- Subject Areas
- Molecular Biology, Plant Science
- Keywords
- Jatropha, Cytokinins, Expression analysis, IPT, Isopentenyl transferases, Trans-zeatin, CYP735A, CRISPR/Cas9, Cytokinin oxidase/dehydrogenase, CKX
- Copyright
- © 2018 Cai et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2018. Identification and expression analysis of cytokinin metabolic genes IPTs, CYP735A and CKXs in the biofuel plant Jatropha curcas. PeerJ 6:e4812 https://doi.org/10.7717/peerj.4812
Abstract
The seed oil of Jatropha curcas is considered a potential bioenergy source that could replace fossil fuels. However, the seed yield of Jatropha is low and has yet to be improved. We previously reported that exogenous cytokinin treatment increased the seed yield of Jatropha. Cytokinin levels are directly regulated by isopentenyl transferase (IPT), cytochrome P450 monooxygenase, family 735, subfamily A (CYP735A), and cytokinin oxidase/dehydrogenase (CKX). In this study, we cloned six IPT genes, one JcCYP735A gene, and seven JcCKX genes. The expression patterns of these 14 genes in various organs were determined using real-time quantitative PCR. JcIPT1 was primarily expressed in roots and seeds, JcIPT2 was expressed in roots, apical meristems, and mature leaves, JcIPT3 was expressed in stems and mature leaves, JcIPT5 was expressed in roots and mature leaves, JcIPT6 was expressed in seeds at 10 days after pollination, and JcIPT9 was expressed in mature leaves. JcCYP735A was mainly expressed in roots, flower buds, and seeds. The seven JcCKX genes also showed different expression patterns in different organs of Jatropha. In addition, CK levels were detected in flower buds and seeds at different stages of development. The concentration of N6-(Δ2-isopentenyl)-adenine (iP), iP-riboside, and trans-zeatin (tZ) increased with flower development, and the concentration of iP decreased with seed development, while that of tZ increased. We further analyzed the function of JcCYP735A using the CRISPR-Cas9 system, and found that the concentrations of tZ and tZ-riboside decreased significantly in the Jccyp735a mutants, which showed severely retarded growth. These findings will be helpful for further studies of the functions of cytokinin metabolic genes and understanding the roles of cytokinins in Jatropha growth and development.
Introduction
Jatropha curcas is a multipurpose tree that belongs to the Euphorbiaceae family. It can endure drought and adapt to barren land in tropical and subtropical regions. Jatropha is considered a promising biofuel plant due to the high oil content in its seeds (Akashi, 2012; Francis, Edinger & Becker, 2005; Makkar & Becker, 2009). However, the seed yield is very low, potentially because of the relatively low number of total flowers and/or the ratio of female to male flowers in Jatropha (Kumar & Sharma, 2008; Kumar Tiwari, Kumar & Raheman, 2007; Rao et al., 2008). Recently, several studies have reported that exogenous cytokinin (CK) treatment can significantly increase the total number of flowers per inflorescence, the female-to-male flower ratio, and the seed yield (Fröschle, Horn & Spring, 2017; Pan & Xu, 2011).
Cytokinins are important hormones in plants and participate in many biological processes, such as apical dominance (Shimizu-Sato, Tanaka & Mori, 2009; Tanaka et al., 2006), root proliferation (Kudo, Kiba & Sakakibara, 2010; Werner et al., 2003), reproductive development (Ashikari et al., 2005), and senescence (Gan & Amasino, 1995). Endogenous CKs containing N6-(Δ2-isopentenyl)-adenine (iP), trans-zeatin (tZ), cis-zeatin (cZ), dihydrozeatin (DZ), and their conjugates are known as isoprenoid CKs (Mok & Mok, 2001). The major derivatives are generally iP- and tZ-type CKs (Sakakibara, 2006).
Cytokinin biosynthesis and degradation pathways have been well studied in the past decade (Fig. 1) (Galuszka et al., 2007; Kudo, Kiba & Sakakibara, 2010; Sakakibara, 2006). The first step of iP and tZ biosynthesis is catalyzed by adenosine phosphate-isopentenyltransferases (IPTs). IPTs produce iP-ribotides from dimethylallyl diphosphate (DMAPP) and adenosine 5′-diphosphate (ADP) or adenosine 5′-triphosphate (ATP) (Ihara et al., 1984; Taya, Tanaka & Nishimura, 1978). iP-ribotides can then be hydroxylated to tZ-ribotides by cytochrome P450 monooxygenase, family 735, subfamily A (CYP735A) (Takei, Yamaya & Sakakibara, 2004). These cytokinin ribotides are converted to free-base CKs by cytokinin-activating enzymes LONELY GUYs (LOGs) (Kurakawa et al., 2007; Kuroha et al., 2009; Tokunaga et al., 2012). In addition, cZ and tZ can be enzymatically interconverted by zeatin cis–trans isomerase (Bassil, Mok & Mok, 1993; Sakakibara, 2006). In Arabidopsis, IPT1 and IPT3–IPT8 are involved in iP and tZ biosynthesis (Kakimoto, 2001; Sun et al., 2003; Takei, Sakakibara & Sugiyama, 2001), while IPT2 and IPT9 are involved in cZ biosynthesis (Golovko et al., 2002). CYP735A1 is abundant in roots and flowers in Arabidopsis, while CYP735A2 specifically accumulates in roots (Takei, Yamaya & Sakakibara, 2004). CYP735As are required for shoot growth (Kiba et al., 2013). Cytokinin oxidase/dehydrogenase (CKX) catalyzes the irreversible degradation of CKs (Galuszka et al., 2001, 2007; Schmulling et al., 2003). CKXs play important roles in controlling CK levels in plant tissues. In Arabidopsis, CKX3 and CKX5 regulate the activity of reproductive meristems (Bartrina et al., 2011). In rice, OsCKX4 mediates crown root development by integrating cytokinin and auxin signaling (Gao et al., 2014).
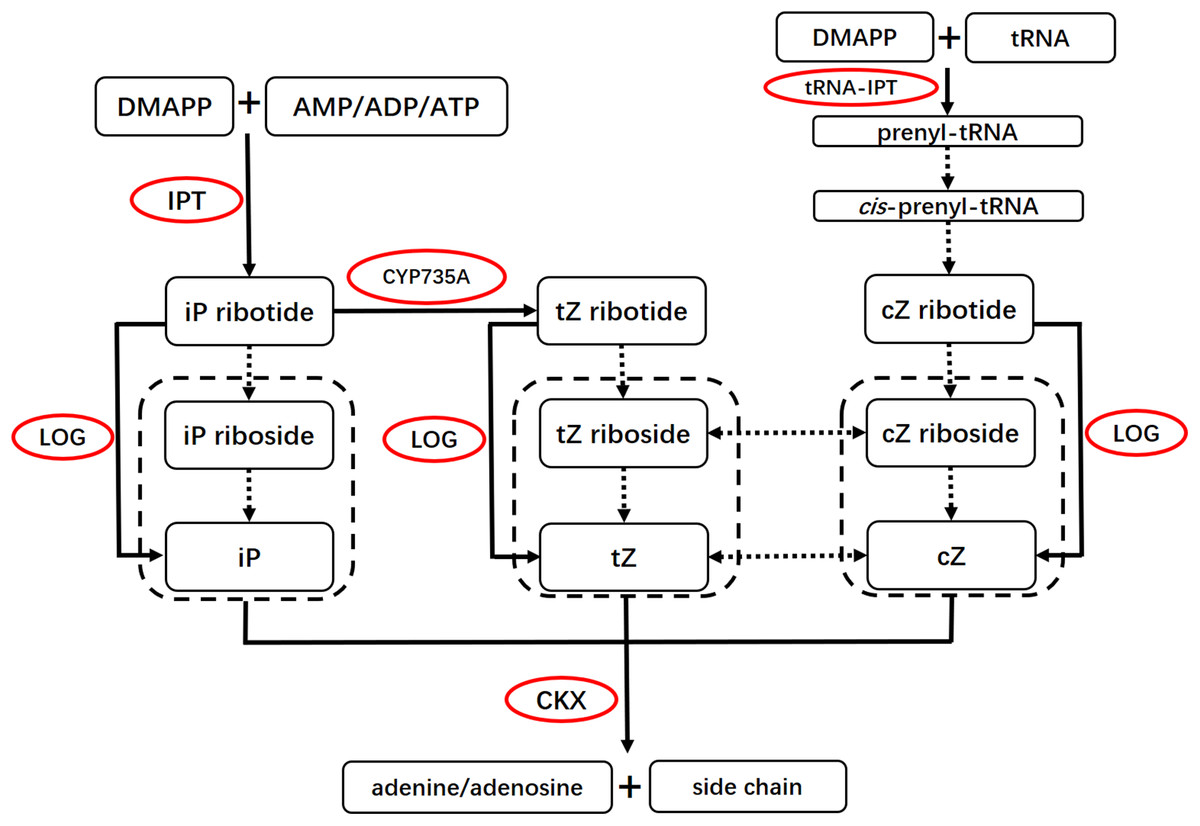
Figure 1: Basic scheme for the cytokinin biosynthesis and degradation pathways.
Solid arrows indicate pathways with genes that are known, and dotted arrows indicate pathways with genes that remain to be identified. The enzymes are marked by red frames. The iP, Z and their ribosides inside the dotted boxes could be degraded by CKX. cZ, cis-zeatin; DMAPP, dimethylallyldiphosphate; CKX, cytokinin oxidase/dehydrogenase; cZ, cis-zeatin; DMAPP, dimethylallyl diphosphate; iP, N6-(Δ2-isopentenyl)adenine; IPT, adenosine phosphate-isopentenyltransferase; LOG, LONELY GUY; tRNA-IPT, tRNA-isopentenyltransferase; tZ, trans-zeatin; Z, zeatin. This figure was modified and redrawn from reference (Kudo, Kiba & Sakakibara, 2010).{kind=link}
Cytokinins play important roles in flower bud development and floral sex differentiation (Chandler, 2011; Gerashchenkov & Rozhnova, 2013; Yamasaki, Fujii & Takahashi, 2005). However, the roles of CK biosynthesis genes IPTs and CYP735A and catabolism gene CKXs in Jatropha are not clear. In this study, we isolated sequences of cytokinin metabolic genes, including six IPTs, one JcCYP735A, and seven JcCKXs, using the Jatropha Genome Database (Hirakawa et al., 2012; Sato et al., 2010; Wu et al., 2015). The 14 genes showed different expression patterns in different tissues of Jatropha. Some of them exhibited tissue-specific expression. JcIPT6 was only expressed in seeds a few days after pollination. JcCYP735A was highly expressed in roots and seeds. JcCKX4 was expressed mainly in seeds. In addition, CK types and contents were detected in flower buds and seeds. With flower bud development iP-type CKs increased, while tZ-type CKs decreased. With seed development, tZ-type CKs increased, while iP-type CKs decreased. The Jccyp735a mutants were obtained by the clustered regularly interspaced short palindromic repeats (CRISPR)-Cas9 system. Compared with the wild-type (WT) plants, the concentrations of tZ and tZ-riboside (tZR) decreased significantly in the Jccyp735a mutants, which showed severely retarded growth. These results will be helpful for future studies of the functions of these genes and for improving the biological characteristics of Jatropha.
Materials and Methods
Plant materials and growth conditions
Three-year-old Jatropha trees were grown in the field at Xishuangbanna Tropical Botanical Garden of the Chinese Academy of Sciences, Mengla County, Yunnan Province, China (21°54′N, 101°46′E; 580 m in altitude). The seedlings of WT and the T1 plants of Jccyp735a mutants were grown in the greenhouse (28°C, 12 h light/12 h dark, 70% humidity). Flower buds, ovules, and seeds at different developmental stages were collected in May–July 2015 for quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR) analysis of JcIPT6 expression and quantification of cytokinin contents. All other samples used in qRT-PCR expriments were collected at the same time in May 2015. Various plant tissue samples, including lateral roots of 1–2 mm in diameter with fine-roots and root tips, shoot apex of 0.3 cm in length from the top of shoots, stems of 1.5 cm in diameter, young leaf blades of 2 cm in length, mature leaf blades of 15 cm in length, flower buds of 0.3 cm in length, just-opened female and male flowers, fruits of 15 days after pollination, and seeds of 30 days after pollination, were harvested for qRT-PCR analysis. All tissues were immediately frozen in liquid nitrogen and stored at −80 °C until needed.
Gene identification and isolation
Sequences of orthologous IPT, CYP735A, and CKX genes from Arabidopsis that were available in the GenBank database were used as query sequences for basic local alignment search tool (BLAST) analysis using GenBank, the Jatropha Genome Database (http://www.kazusa.or.jp/jatropha/index.html) and our Jatropha transcriptome data (Chen et al., 2014; Pan et al., 2014). The full length of complementary DNA (cDNA) and genomic DNA sequences of JcIPT, JcCYP735A, and JcCKX were obtained by PCR amplification. The PCR products were subsequently cloned into the pGEM-T vector (Promega Corporation, Madison, WI, USA) and sequenced. The GenBank accession numbers for the nucleotide sequences of these genes are listed in Table S1. Primers used in PCR are listed in Table S2.
Sequence comparison and phylogenetic analysis
Sequence chromatograms were examined and edited using Chromas Version 2.23 (http://technelysium.com.au/). Related sequences were identified with BLAST (http://www.ncbi.nlm.nih.gov/BLAST/). Genomic organization of all genes was analyzed by using the Gene Structure Display Server (GSDS) with default settings (Hu et al., 2015). A phylogenetic tree was generated with MEGA 7.0 (http://www.megasoftware.net/) using the Poisson model with gamma-distributed rates and 1,000 bootstrap replicates.
Expression pattern analysis by qRT-PCR
Total RNA was extracted from each tissue, and first-strand cDNA was synthesized with a PrimeScript® RT Reagent Kit with gDNA Eraser (Takara, Dalian, China) according to the manufacturer’s instructions. qRT-PCR was performed with LightCycler® 480 SYBR Green I Master (Roche, Indianapolis, IN, USA) on the Roche 480 Real-Time PCR Detection System (Roche Diagnostics, Mannheim, Germany). qRT-PCR was performed with two independent biological replicates (tissue samples were harvested from different plants) and three technical replicates for each sample. Data were analyzed using the 2−ΔΔCT method as described by Livak & Schmittgen (2001). Expression levels of specific genes were normalized to that of the actin gene in Jatropha (Zhang et al., 2013). Primers used in qRT-PCR are listed in Table S3.
Quantification of cytokinin
Cytokinin contents were determined by the Wuhan Greensword Creation Technology Co. Ltd., using a polymer monolith microextraction coupled with hydrophilic interaction chromatography-tandem mass spectrometry method as described previously (Liu, Wei & Feng, 2010).
The leaves used to quantify the CKs were the third and fourth new leaves from four-month-old WT and the T1 plants of Jccyp735a mutants. Three independent biological replicates and three technical replicates were measured for each sample. The data were analyzed using the Statistical Product and Service Solutions software (SPSS Inc., Chicago, IL, USA, version 16.0). Differences among the means were determined using a one-way ANOVA with Tukey’s or Tamhane’s post hoc tests (p < 0.05).
Construction of CRISPR/Cas9 vectors and transformation of Jatropha
The sequence of JcCYP735A (GenBank accession no. XM_012222581.2) was analyzed with the online tool CRISPR-P (http://cbi.hzau.edu.cn/crispr/) to find the target sites of CRISPR/Cas9. pYLsgRNA-AtU3d/LacZ (GenBank accession no. KR029100) as the single-guide RNA (sgRNA) intermediate plasmid, and pYLCRISPR/Cas9P35S-N (GenBank accession no. KR029112) as the binary vector were used for the CRISPR-Cas9 construction of JcCYP735A following the instruction of the CRISPR-Cas9 system (Ma et al., 2015). Transformation of Jatropha with Agrobacterium strain EHA105 carrying the JcCYP735A CRISPR/Cas9 construction was performed according to the protocol described by Fu et al. (2015). The Jccyp735a mutants in transgenic Jatropha plants were identified by PCR amplification and DNA sequencing using a pair of primers, XB619 (5′-ATGGCCATGATATTAACAACTCTATTAG-3′) and XB620 (5′-GCGGTTCTATCCCATTCCAGTATAT-3′).
Results
Cloning and identification of JcIPTs, JcCYP735A, and JcCKXs
Using all annotated Arabidopsis IPT, CYP735A, and CKX family members in the TAIR as query sequences to perform a BLAST analysis in GenBank and with our Jatropha transcriptome data (Chen et al., 2014; Pan et al., 2014), we identified and cloned IPT, CYP735A, and CKX orthologous sequences in Jatropha. The Jatropha IPT family included only six members, while there are nine members in Arabidopsis. These genes were named JcIPT1, JcIPT2, JcIPT3, JcIPT5, JcIPT6, and JcIPT9. The BLAST analysis identified only one member of the CYP735A family, JcCYP735A. The CKX gene family, encoding degradation enzymes, included the same seven members in Jatropha as in Arabidopsis. These genes were named JcCKX1, JcCKX2, JcCKX3, JcCKX4, JcCKX5, JcCKX6, and JcCKX7.
Sequence structure analysis showed that IPT, CYP735A, and CKX family members shared almost the same numbers of exons and introns between Jatropha and Arabidopsis and had similar exon lengths (Figs. 2 and 3). JcIPT6 has two more exons than IPT6 from Arabidopsis. However, the extra two exon sequences are short and are not part of the P-loop NTPase domain (Fig. 2A).
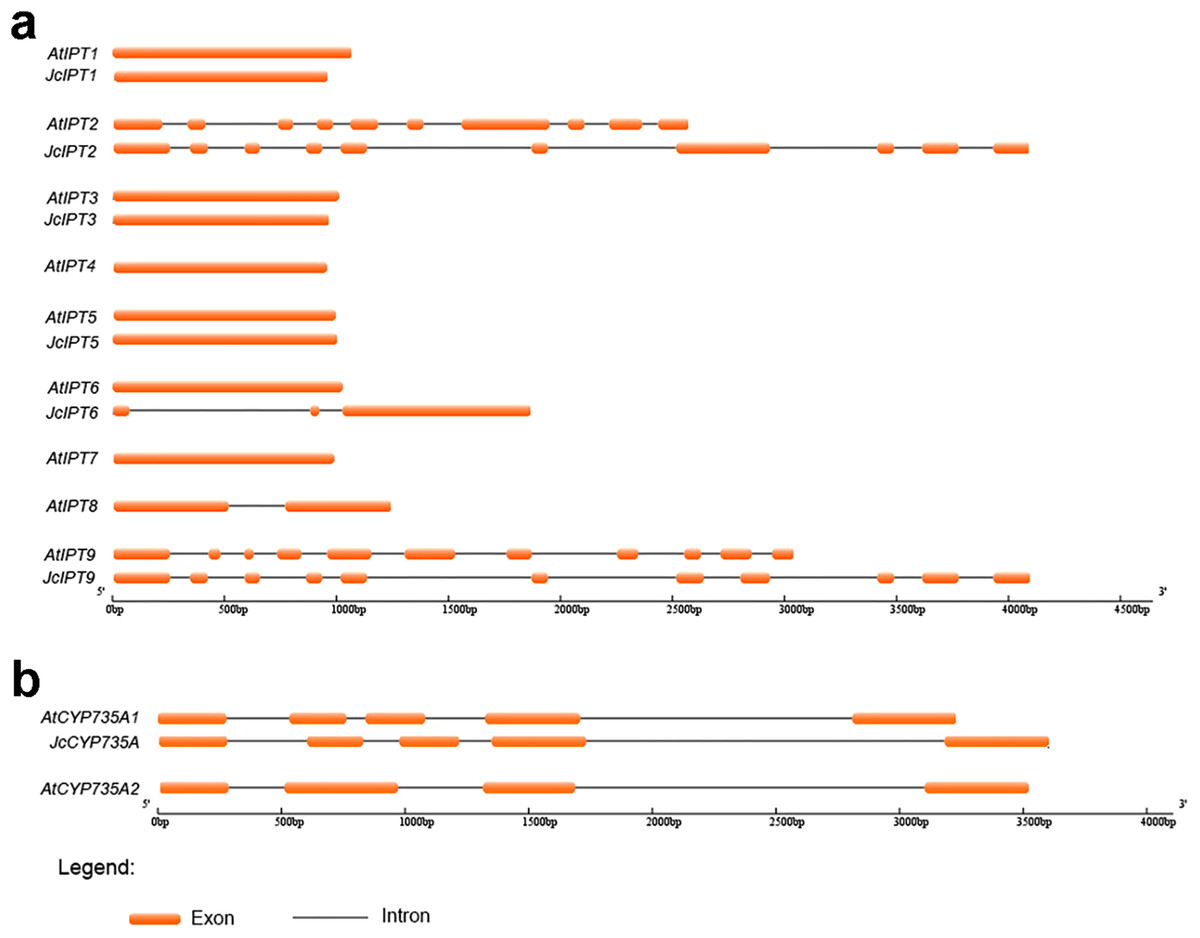
Figure 2: Genomic organization of IPT and CYP735A family members in Jatropha and Arabidopsis.
(A) IPT family members; (B) CYP735A family members. At, Arabidopsis thaliana; Jc, Jatropha curcas.{kind=link}
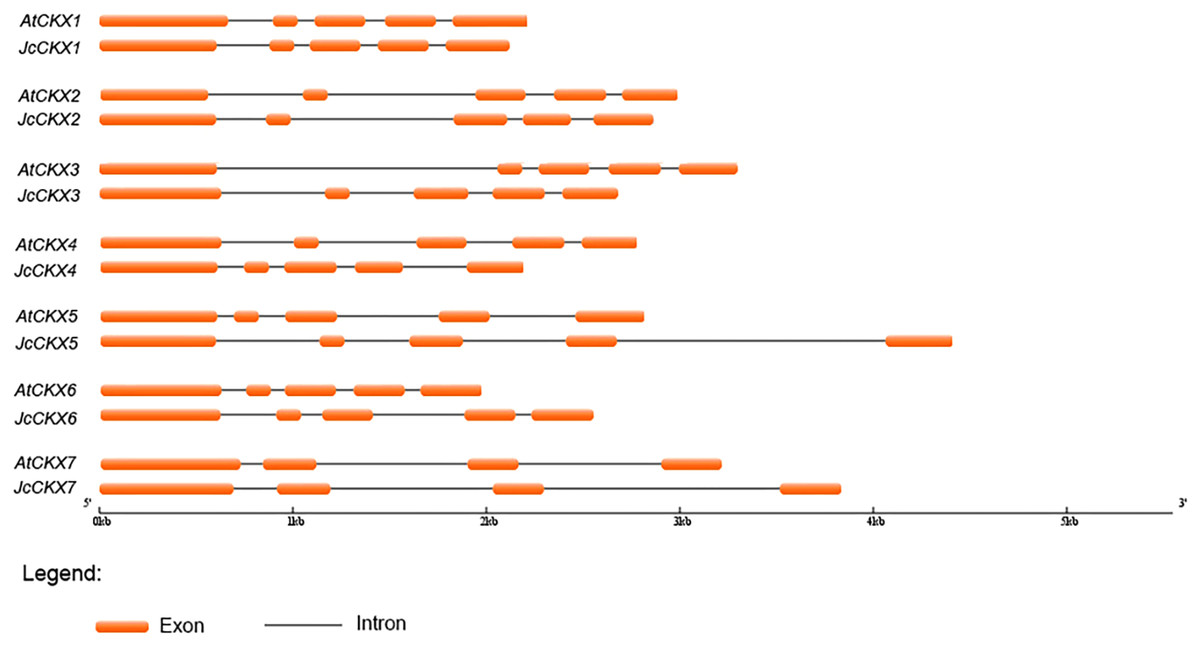
Figure 3: Genomic organization of CKX family members in Jatropha and Arabidopsis.
At, Arabidopsis thaliana; Jc, Jatropha curcas.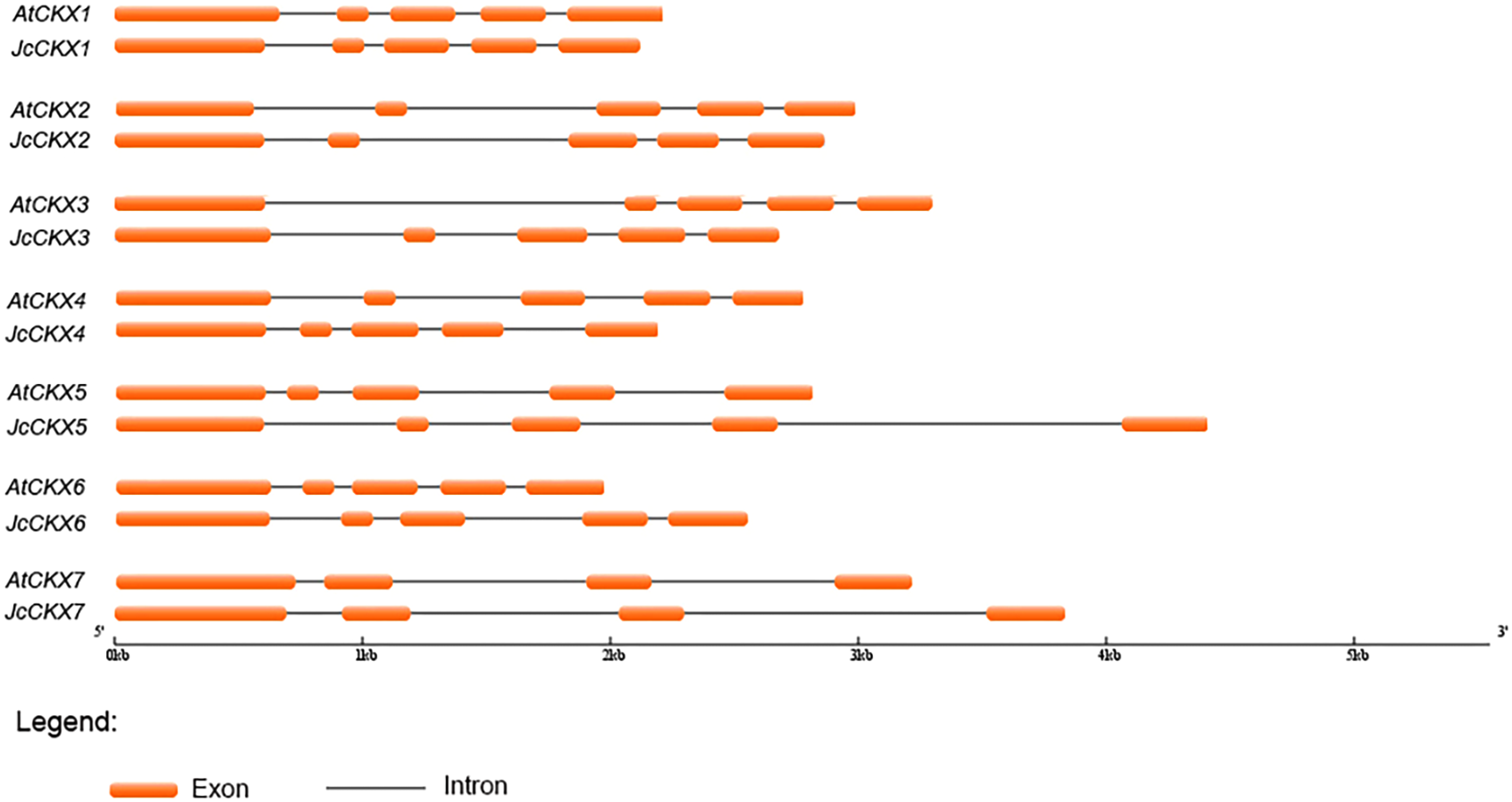{kind=link}
Phylogenetic analysis of JcIPTs, JcCYP735A, and JcCKXs
To analyse the phylogenetic relationships between orthologous genes, phylogenetic analysis were performed. IPT, CYP735A, and CKX family members from Arabidopsis thaliana, Ricinus communis, and Oryza sativa were compared with those from Jatropha. Orthologues of IPT1, 2, 3, 5, and 7 formed a clade, while IPT9 formed a single clade (Fig. 4A). JcCYP735A along with other dicotyledon CYP735As formed a clade, while CYP735A3 and 4 of O. sativa formed another clade (Fig. 4B). Orthologues of CKX1, 5, 6, and 7 formed a clade, while those of CKX2, 3, and 4 formed a separate clade (Fig. 5). These results showed that JcIPTs, JcCYP735A, and JcCKXs were most closely related to genes from R. communis, which also belongs to the Euphorbiaceae family.
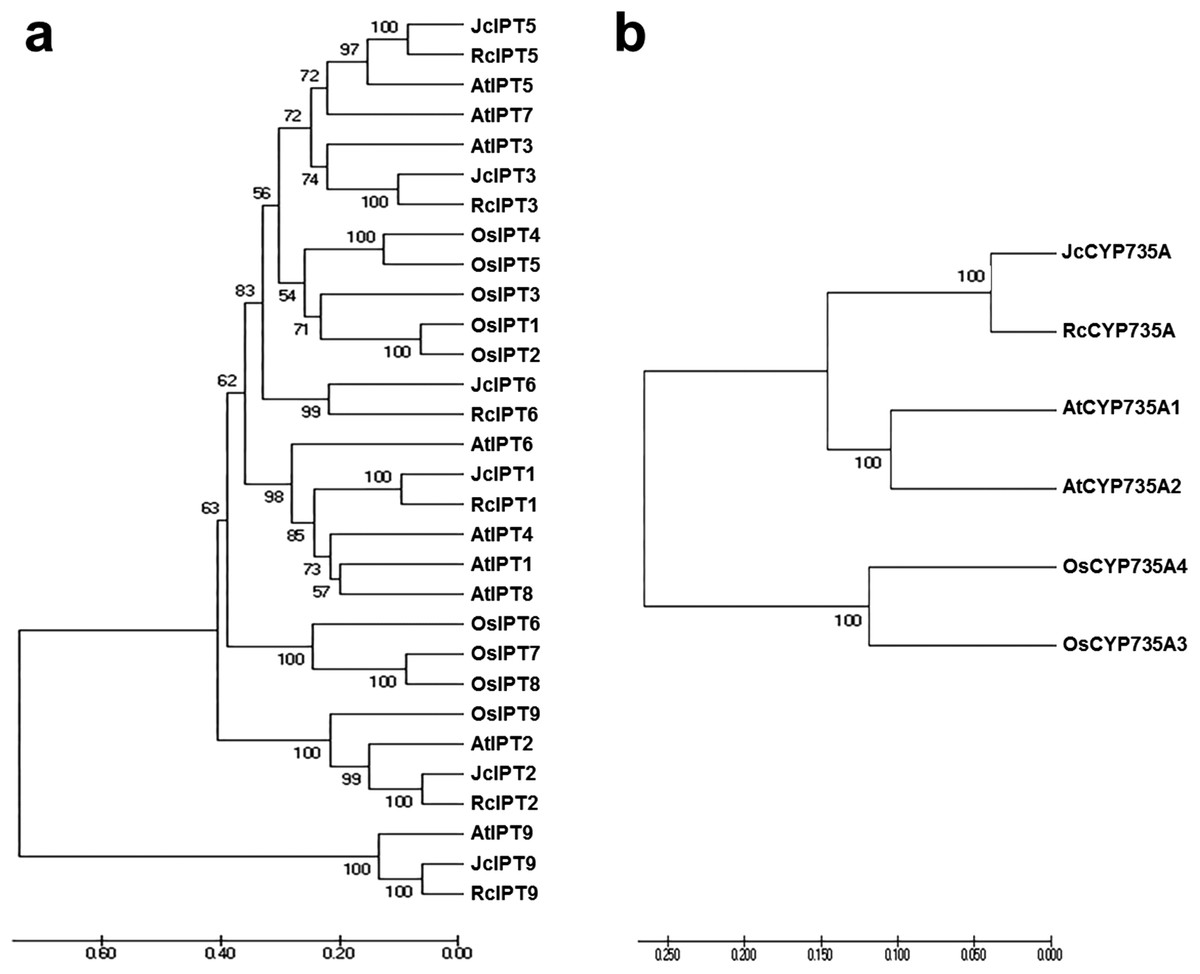
Figure 4: Neighbor-joining phylogenetic tree for IPT and CYP735A family members in various species.
(A) IPT family members; (B) CYP735A family members. At, Arabidopsis thaliana; Jc, Jatropha curcas; Os, Oryza sativa; Rc, Ricinus communis.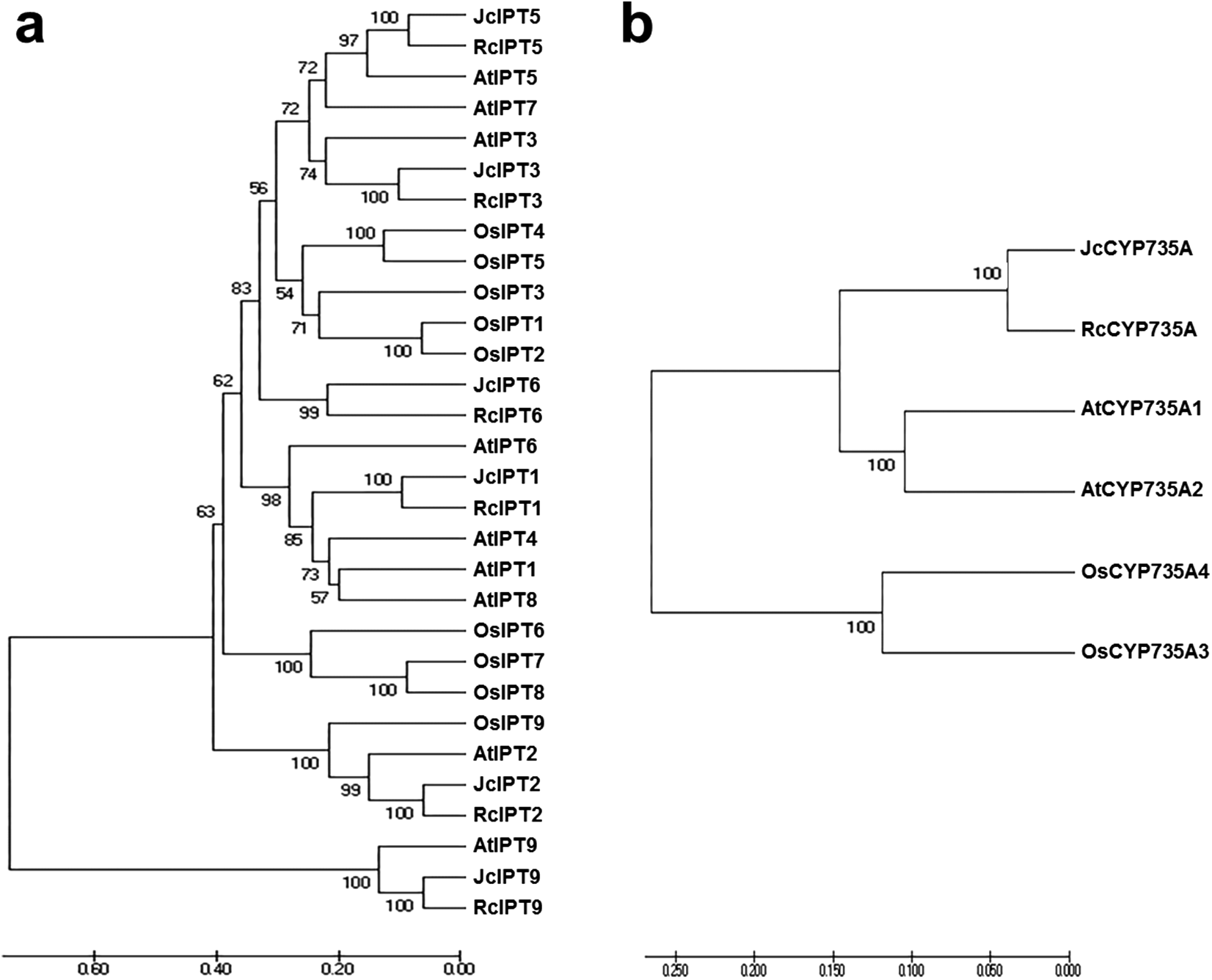{kind=link}
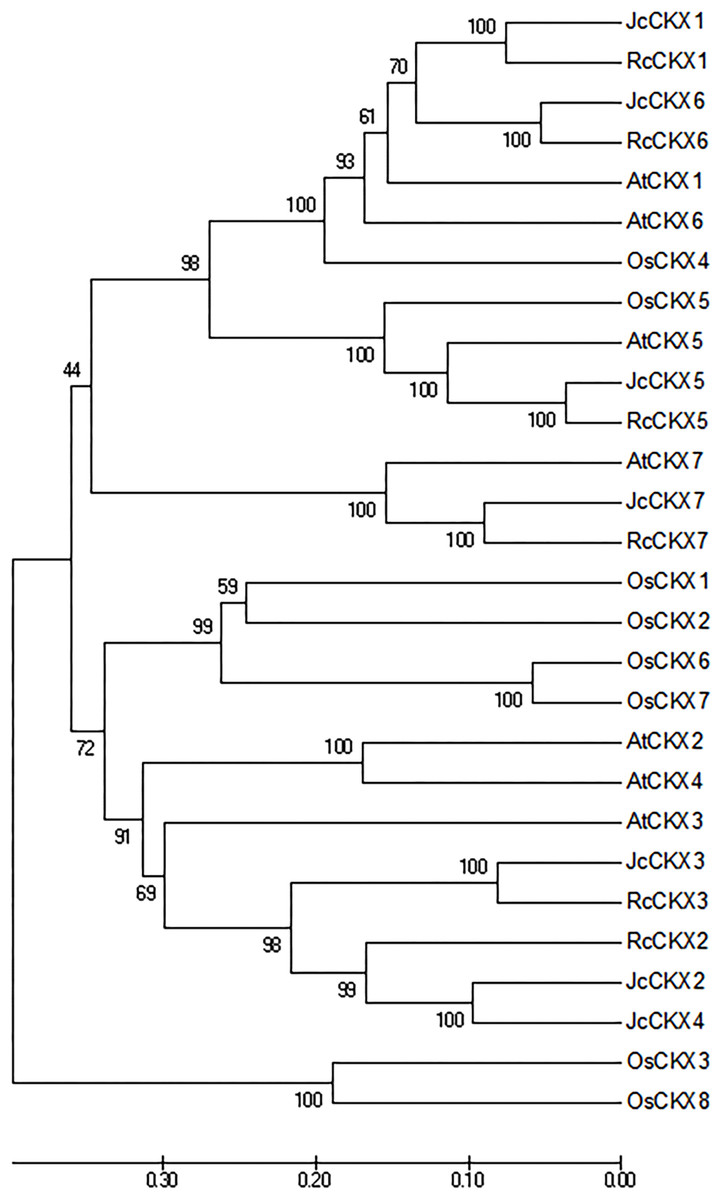
Figure 5: Neighbor-joining phylogenetic tree for CKX family members in various species.
At, Arabidopsis thaliana; Jc, Jatropha curcas; Os, Oryza sativa; Rc, Ricinus communis.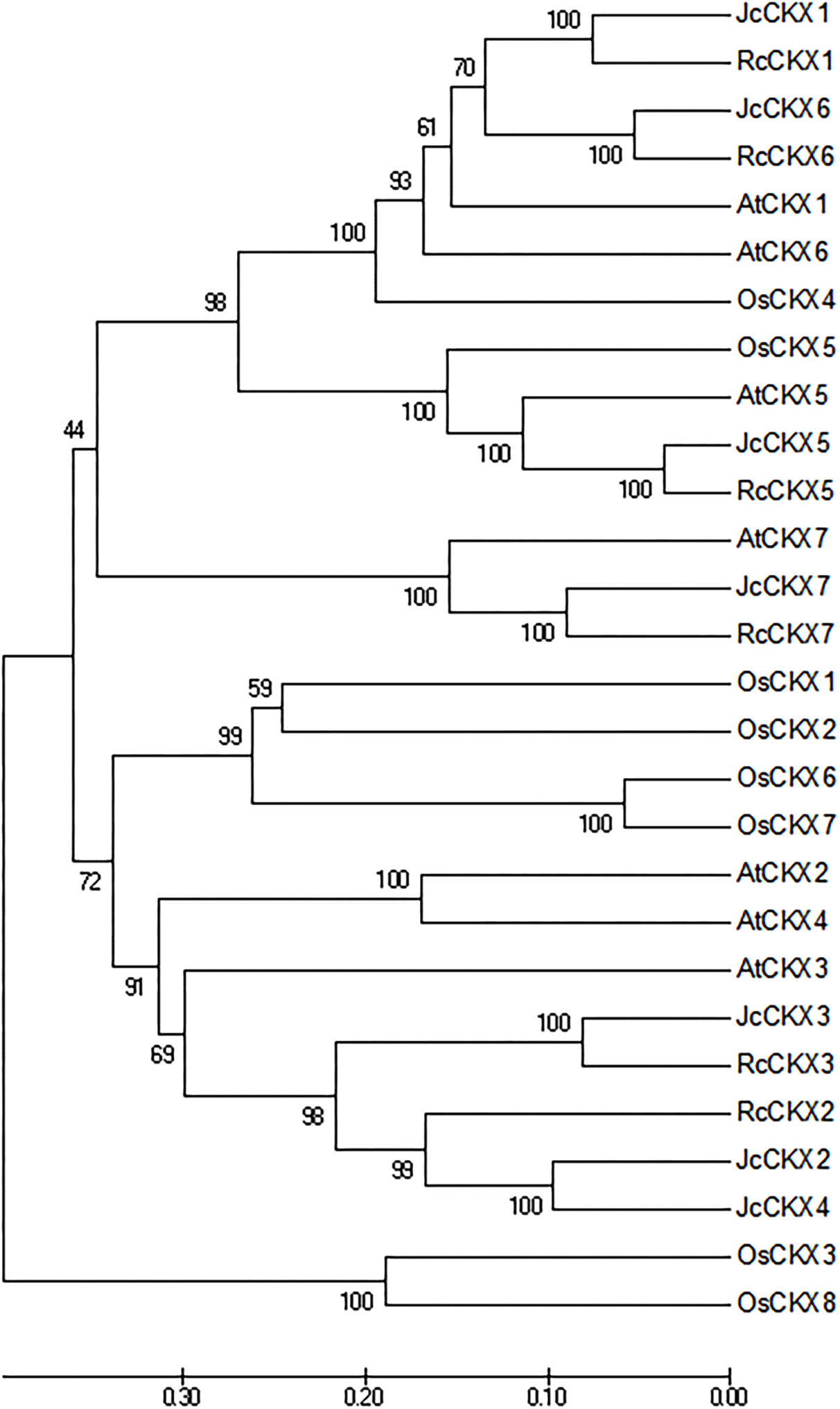{kind=link}
Expression patterns of JcIPTs, JcCYP735A, and JcCKXs in different tissues
In order to gain more information of these gene family members in Jatropha, the temporal and spatial expression patterns of these genes were analyzed using qRT-PCR. JcIPT1 was mainly expressed in roots (Fig. 6A). JcIPT2 was mainly expressed in roots, shoot apical meristems, mature leaves, and flower buds (Fig. 6B). JcIPT3 showed much higher expression levels in stems and mature leaves than other tissues (Fig. 6C). JcIPT5 exhibited high expression levels in roots and mature leaves (Fig. 6D). JcIPT9 only showed high expression levels in mature leaves (Fig. 6F). The expression of JcIPT6 was not detected in most of the plant tissues indicated above. After analysing more tissues (Fig. S1), we found that JcIPT6 began to be expressed in seeds a few days after fertilization, with the strongest expression observed in seeds at 10 days after fertilization; the expression levels then decreased rapidly. In seeds at 20 days after fertilization, JcIPT6 expression decreased by a factor of 5 compared with seeds at 10 days after fertilization (Fig. 6E). The expression levels of JcCYP735A were higher in roots, flower buds, and seeds than other tissues (Fig. 7A). JcCKX1 was mainly expressed in flower buds, roots, and female flowers (Fig. 7B). JcCKX2 showed very strong expression in female flowers and seeds (Fig. 7C). JcCKX3 was highly expressed in male flowers (Fig. 7D). JcCKX4 exhibited high expression levels in mature leaves and female flowers, and extremely high expression in seeds (Fig. 7E). JcCKX5 was mainly expressed in stems, young leaves, and fruit (Fig. 7F). JcCKX6 was expressed in all tissues (Fig. 7G). JcCKX7 was mainly expressed in roots (Fig. 7H).
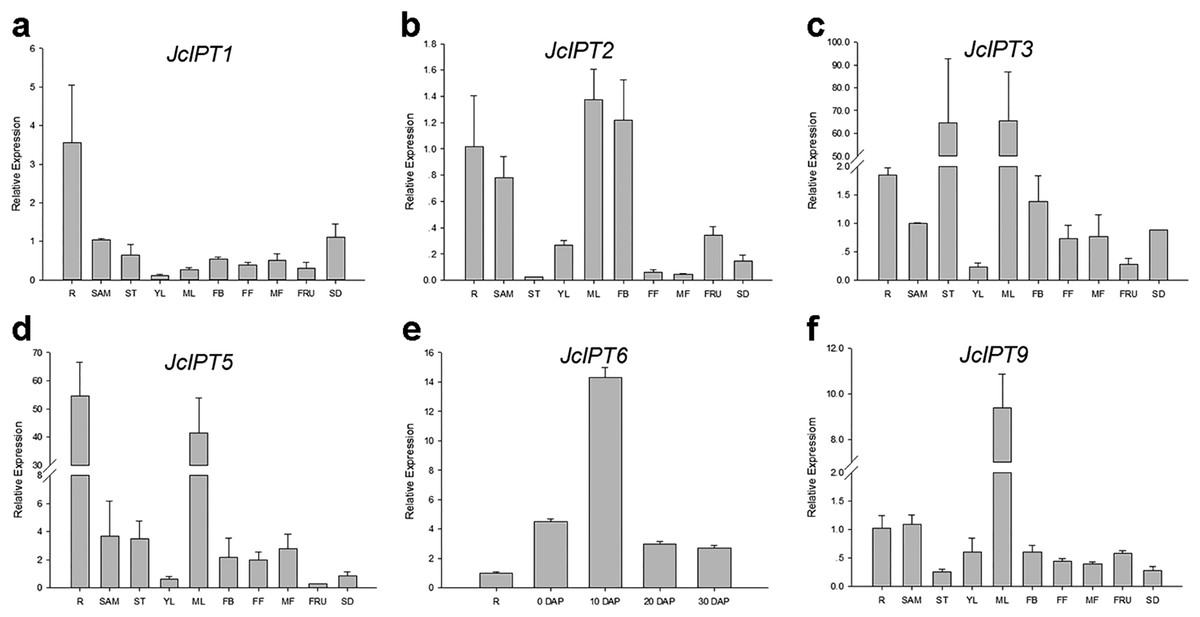
Figure 6: Expression of JcIPTs in various Jatropha tissues.
(A–F) are expression patterns of JcIPT1, JcIPT2, JcIPT3, JcIPT5, JcIPT6, and JcIPT9, respectively. The qRT-PCR results were obtained from two independent biological replicates and three technical replicates for each sample. R, roots; SAM, shoot apical meristems; ST, stems; YL, young leaves; ML, mature leaves; FB, flower buds; FF, female flowers; MF, male flowers; FRU, fruits; SD, seeds; 0 DAP, unfertilized ovules; 10 DAP, seeds at 10 days after pollination; 20 DAP, seeds at 20 days after pollination; 30 DAP, seeds at 30 days after pollination; DAP, days after pollination.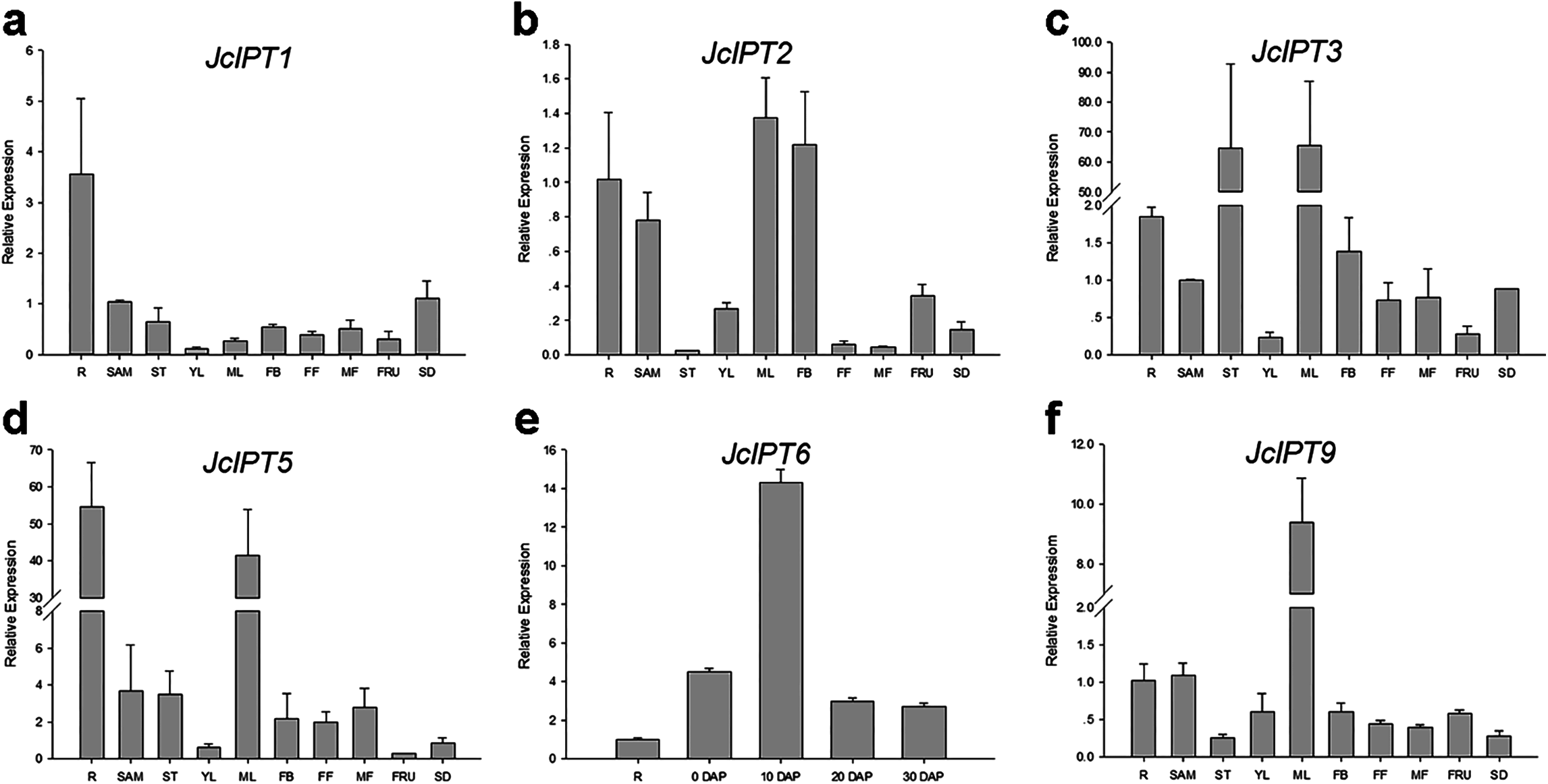{kind=link}
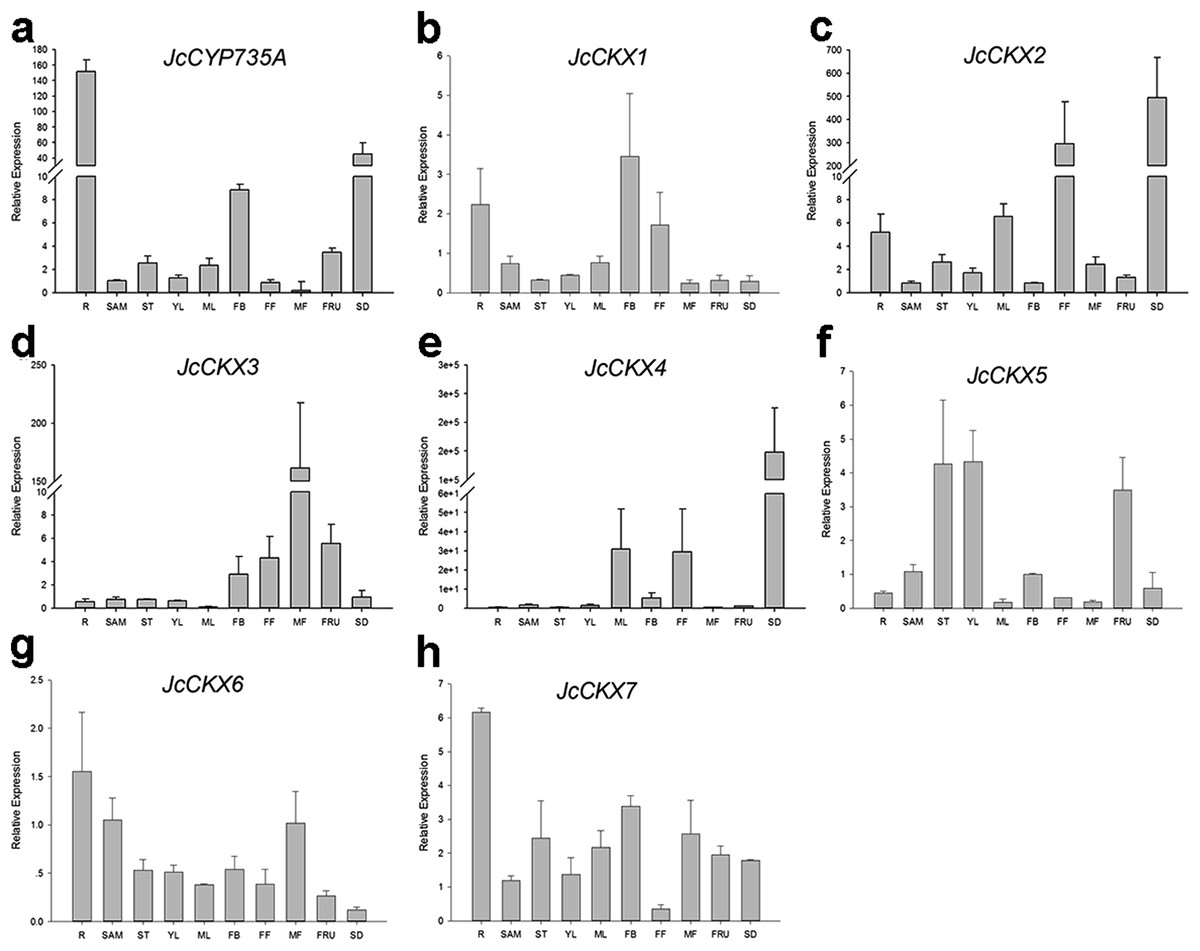
Figure 7: Expression of JcCYP735A and JcCKXs in various Jatropha tissues.
(A–H) are expression patterns of JcCYP735A, JcCKX1, JcCKX2, JcCKX3, JcCKX4, JcCKX5, JcCKX6, and JcCKX7, respectively. Values in the y-axis of (E) are displayed in scientific notation. e, exponent. The qRT-PCR results were obtained from two independent biological replicates and three technical replicates for each sample. R, roots; SAM, shoot apical meristems; ST, stems; YL, young leaves; ML, mature leaves; FB, flower buds; FF, female flowers; MF, male flowers; FRU, fruits; SD, seeds.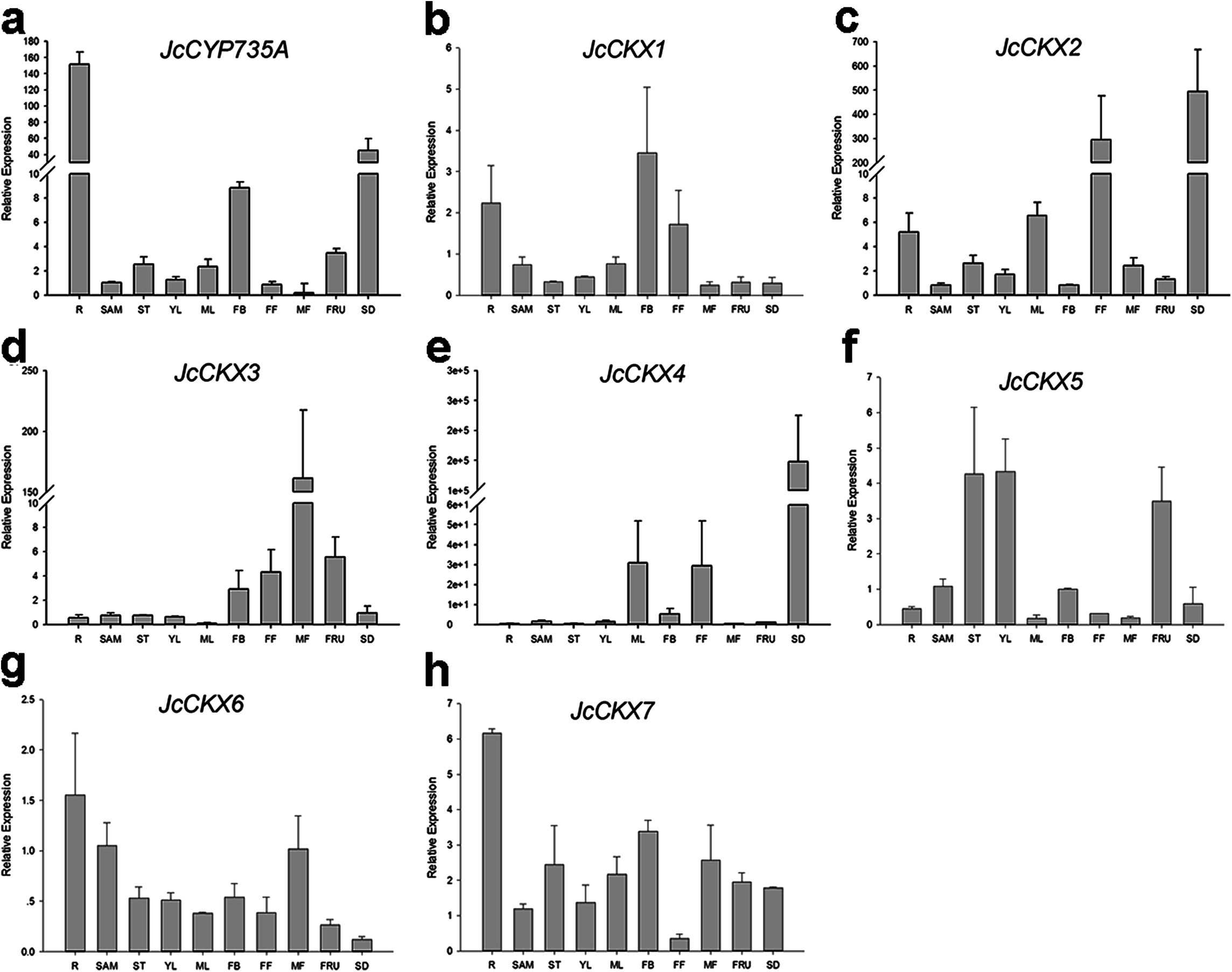{kind=link}
Endogenous CK contents in flower buds and seeds
In order to learn more about the distribution of endogenous cytokinins, we measured the contents of endogenous CKs in Jatropha flower buds and seeds at different developmental stages (Fig. S1). Different profiles were observed for each CK variant with flower bud and seed development in Jatropha (Table 1). The contents of iP and its variant iP-riboside (iPR) increased approximately 177-fold and nine-fold, respectively, from the flower bud 1 (FB1) stage to the flower bud 2 (FB2) stage. Compared with the FB1 stage, tZ content was approximately doubled in the FB2 stage, while tZR content was reduced by 93%, resulting in a decrease in the amount of total active tZ variants. Conversely, the contents of tZ variants increased remarkably during seed development; compared with ovules, the tZ content increased 187-fold and the tZR content increased 496-fold in seeds at 20 DAP.
| Samples | iP | iPR | iP9G | tZ | tZR | tZ9G | DZ | DZR |
|---|---|---|---|---|---|---|---|---|
| FB1 | 0.14 ± 0.01 | 1.50 ± 0.03 | N.D. | 1.21 ± 0.03 | 12.55 ± 0.78 | N.Q. | 1.27 ± 0.09 | 9.24 ± 0.15 |
| FB2 | 24.86 ± 0.83 | 15.24 ± 0.24 | N.D. | 2.49 ± 0.17 | 0.89 ± 0.08 | N.D. | 3.31 ± 0.19 | 0.97 ± 0.03 |
| 0 DAP | 3.97 ± 0.40 | 1.20 ± 0.07 | N.D. | 0.79 ± 0.08 | 0.08 ± 0.007 | N.D. | 0.5 ± 0.06 | N.D. |
| 10 DAP | 3.96 ± 0.19 | 0.71 ± 0.02 | N.D. | 22.61 ± 2.42 | 1.56 ± 0.09 | 0.06 ± 0.001 | 6.42 ± 0.24 | 0.18 ± 0.02 |
| 20 DAP | 0.31 ± 0.01 | 0.78 ± 0.05 | N.D. | 149.28 ± 11.60 | 39.76 ± 1.80 | 0.75 ± 0.03 | 28.90 ± 2.68 | 16.56 ± 0.86 |
Notes:
FB1, flower buds of less than 5 mm in length; FB2, flower buds of two to three cm in length; 0 DAP, unfertilized ovule; 10 DAP, seeds of 10 days after pollination; 20 DAP, seeds of 20 days after pollination; DAP, days after pollination; N.D., not detected; N.Q., not quantified.
Jccyp735a mutants generated by CRISPR-Cas9 system showed retarded growth
To explore the biological function of JcCYP735A in Jatropha, we generated Jatropha transformants with JcCYP735A knocked out using the CRISPR-Cas9 system (Fig. 8). Three homozygous mutant lines, L1, L2, and L3, were obtained by DNA sequencing (Fig. 8A). Endogenous contents of CKs in the leaves of two lines of Jccyp735a mutants (L2 and L3) and WT plants were examined. The results showed that the concentrations of tZ and tZR, and cZ and cZR significantly decreased, whereas the concentrations of iP and iPR significantly increased in Jccyp735a mutants compared with those of the WT (Fig. 8B). Jccyp735a mutants showed severely retarded growth, and the mutant plants were only approximately a quarter the height of the WT plants (Figs. 8C and 8D).
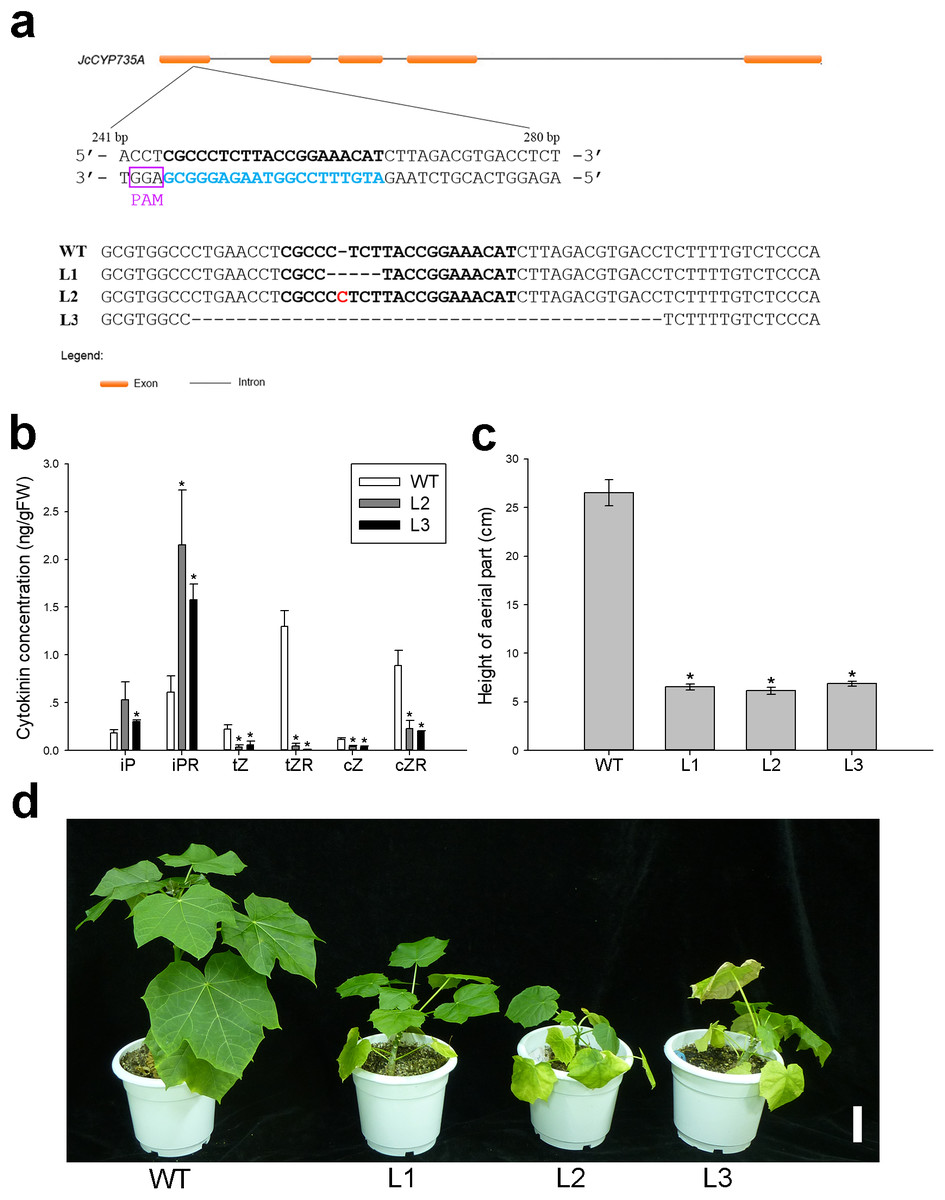
Figure 8: Generation and phenotypic variation of Jccyp735a mutants of Jatropha obtained by the CRISPR-Cas9 system.
(A) Different types of JcCYP735A mutation generated by the CRISPR-Cas9-mediated gene silencing in the progenies (T1 generation) of the three transgenic lines L1, L2, and L3. The blue characters show the selected target site sequences. The characters in purple box show the protospacer adjacent motif (PAM) sequences. The red character indicates the nucleotide insertion. WT, wild-type. (B–D) Cytokinin concentrations (B), height (C), and appearance (D) of the four-month-old seedlings (T1) of Jccyp735a mutant lines and WT. Error bars represent the standard deviation (SD) of three (B) or five (C) biological replicates. Asterisks indicate statistically significant differences compared with WT (p < 0.05). In (D), bar = 5 cm. Photo by Li Cai.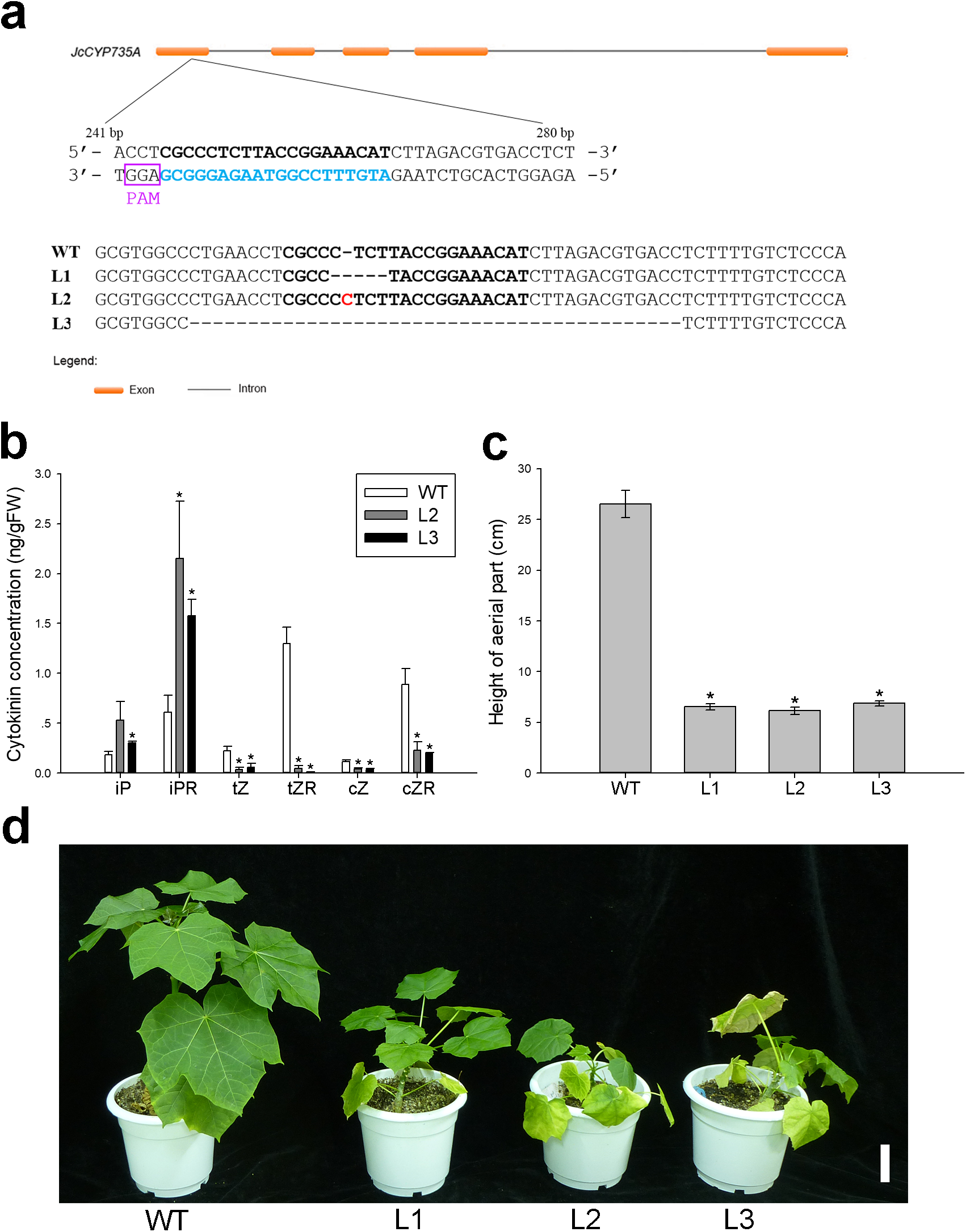{kind=link}
Discussion
In our study, six IPT family members were identified in Jatropha. The number of IPT genes differs among plant species; for example, there are six IPTs in R. communis, while there are nine IPTs in both Arabidopsis (Kakimoto, 2001; Takei, Sakakibara & Sugiyama, 2001) and rice (Fig. 4A). JcIPTs have the same number of exons as those in Arabidopsis, except JcIPT6, which has two more exons than AtIPT6. The third exon of JcIPT6 has almost the same number of base pairs as the only exon in AtIPT6 (Fig. 2A). It appears that the other two small exons were lost during evolution. Expression pattern analysis revealed that different JcIPT members were expressed in different tissues in Jatropha (Fig. 6). JcIPT1 had an expression pattern similar to that of AtIPT1 and was mostly expressed in roots, shoot apical meristems, and seeds (Miyawaki, Matsumoto-Kitano & Kakimoto, 2004). JcIPT2 and JcIPT9 were assigned to the same cluster as their orthologues (Fig. 4A). In Arabidopsis, AtIPT2 and AtIPT9 are expressed ubiquitously, with stronger expression in proliferating tissues, including the root and shoot apical meristems and leaf primordia (Miyawaki, Matsumoto-Kitano & Kakimoto, 2004). Similarly, JcIPT2 and JcIPT9 were expressed ubiquitously in Jatropha. However, the strongest expression of both was in mature leaves instead of meristems and young leaves (Figs. 6B and 6F). Considering that AtIPT2 and AtIPT9 are tRNA-IPTs, which only catalyze cZ biosynthesis (Golovko et al., 2002), cZ might be produced mainly in mature leaves of Jatropha. AtIPT3 was found to be responsible for nitrate-dependent cytokinin biosynthesis and is predominantly expressed in the phloem (Miyawaki, Matsumoto-Kitano & Kakimoto, 2004; Takei et al., 2002). In our study, JcIPT3 was strongly expressed in stems and mature leaves, which contain abundant phloem (Fig. 6C). A study by Miyawaki, Matsumoto-Kitano & Kakimoto (2004) showed that GUS activity was not detected in Arabidopsis transformants carrying AtIPT6::GUS. RT-PCR analysis indicated that it was abundant in siliques. Analogously, we did not detect JcIPT6 in most tissues of Jatropha, including fruits. We found that JcIPT6 was mainly expressed in seeds at 10 days after pollination, and its expression decreased rapidly thereafter (Fig. 6E). It is possible that the previous study missed this critical phase in the seeds chosen for GUS staining (Miyawaki, Matsumoto-Kitano & Kakimoto, 2004). Our results suggested that different JcIPT family members play different roles in the development of Jatropha and that some JcIPT members could be used to cultivate high-yield varieties of Jatropha using transgenic technology.
Although none of the ATP/ADP JcIPTs were found to be highly expressed in flowers, iP, iPR, and tZ contents increased with the development of flower buds (Table 1). It is known that CKs can be transported through the plant vascular system (Hirose et al., 2007). JcCYP735A was found to be highly expressed in flower buds (Fig. 7A). iP-type CKs may be transported into flowers, and some of them may then be used to generate tZ-type CKs via JcCYP735A. In addition, it has been reported that the tZ concentration is up-regulated in the early development of tomato fruits (Matsuo et al., 2012). Our study showed that JcCYP735A was highly expressed and that the tZ concentration increased with seed development in Jatropha (Table 1). Thus, JcCYP735A might play important roles in seed development by controlling tZ biosynthesis. Similar to R. communis (Chan et al., 2010), only one CYP735A gene was found in Jatropha, although there are two CYP735A members in Arabidopsis and rice (Takei, Yamaya & Sakakibara, 2004; Tsai et al., 2012).
Unlike JcIPT or JcCYP735A family, the JcCKX family in Jatropha has the same number of members as that in Arabidopsis (Fig. 3). In addition, the JcCKX and AtCKX orthologues contain the same number of exons, with five exons in CKX1-6 and 4 exons in CKX7 (Fig. 3). Further expression analysis showed that JcCKX2 was mostly expressed in female flowers, whereas JcCKX3 was mostly expressed in male flowers (Figs. 7C and 7D). Both JcCKX2 and JcCKX4 were expressed strongly in seeds (Figs. 7C and 7E). These tissue-specific expression genes may be chosen to adjust the CK content in these tissues using transgenic methods. It has been reported that reduced expression of OsCKX2 causes cytokinin accumulation in inflorescence meristems and increases the number of reproductive organs, resulting in enhanced grain yield (Ashikari et al., 2005). Decreased expression of CKX orthologues may also lead to increased yield in Jatropha. Moreover, overexpression of some CKX members can also improve resistance. Overexpression of CKX1 or CKX2 in Arabidopsis and other species causes elongation of the primary root and increases root branching (Galuszka et al., 2004; Mrízová et al., 2013; Pospisilova et al., 2016; Werner et al., 2001; Yang et al., 2003), while overexpression of AtCKX7 results in an opposite phenotype (Kollmer et al., 2014). Specific expression of JcCKX1 or JcCKX2 in roots might be used to transform a shallow root system into a deep root system to improve the growth and lodging resistance of Jatropha. Furthermore, root system development might enhance tolerance to drought stress. Remarkably, JcCKX4 expression was much higher in seeds than other tissues (Fig. 7E), suggesting that JcCKX4 may be a key enzyme regulating cytokinin levels to affect seed development.
In early flower bud development, the content of iP-type CKs increased significantly while that of tZ-type CKs decreased. This result indicated that iP-type CKs participate more in early flower bud development than tZ-type CKs. In tomato, iP-type CK contents decrease during fruit ripening (Matsuo et al., 2012). By contrast, in early seed development, the content of tZ-type CKs increased substantially, while that of iP-type CKs decreased. This result, which is in accordance with high expression of JcCYP735A in seeds, suggested that tZ-type CKs are dominant in early seed development. Many differences were observed in CK contents in different periods of Jatropha flower bud and seed development. Our results indicate that iP-type CKs can be used to improve the number of flowers in Jatropha, while tZ-type CKs can be used to enlarge seeds.
The single cyp735a1 or cyp735a2 mutant Arabidopsis showed slight decreases in tZ and tZR concentration compared with that of WT, while the double mutants showed great decreases (Kiba et al., 2013). In addition, cyp735a1 cyp735a2 double mutants exhibited retarded shoot growth (Kiba et al., 2013). Similarly, in this study, Jccyp735a mutants showed substantial decreases in tZ and tZR concentrations (Fig. 8B), which is consistent with that only a single member of JcCYP735A was found in Jatropha genome (Wu et al., 2015). We noticed, however, tZ and tZR did not completely disappear in Jccyp735a mutants (Fig. 8B), which may result from conversion of cZ and cZR. This notion is supported by the fact that the concentrations of cZ and cZR also significantly decreased in Jccyp735a mutants (Fig. 8B), and that cis–trans isomerase activity for interconversion between cZ-type and tZ-type CKs has been reported in several plant species (Bassil, Mok & Mok, 1993; Kudo et al., 2012; Suttle & Banowetz, 2000).
Conclusion
In this study, we isolated the members of the JcIPT, JcCYP735A, and JcCKX gene families and analyzed their temporal and spatial expression patterns. Different family members exhibited different expression patterns. Different types of CKs seemed to influence the development of flower buds and seeds, respectively. The analysis of the Jccyp735a mutants revealed that JcCYP735A plays an important role in tZ biosynthesis in Jatropha. These results will be helpful for further function studies of cytokinin metabolic genes and improving agronomic characteristics of Jatropha by genetic engineering of cytokinin metabolism.
Supplemental Information
Fig. S1. Flower buds, ovules and seeds of Jatropha used in this study.
(A) Flower buds from the FB1 stage. Bar = 1 cm. (B) Flower buds from the FB2 stage. Bar = 1 cm. (C) Unfertilized Jatropha pistil, from which the ovule was obtained. Bar = 1 mm. (D) Morphology of unfertilized ovules. Bar = 1 mm. (E) Morphology of fruits and seeds. The first row contains fruits at different developmental periods. The second row shows the seeds of the corresponding fruits. Bar = 1 cm. 0 DAP, unfertilized ovules; 10 DAP, seeds at 10 days after pollination; 20 DAP, seeds at 20 days after pollination; 30 DAP, seeds at 30 days after pollination; DAP, days after pollination. Photos by Li Cai.
{kind=link}
Table S1. GenBank accession numbers of the gene sequences in this work.
Raw data.