
Advances in cultivation, wastewater treatment application, bioactive components of Caulerpa lentillifera and their biotechnological applications
- Published
- Accepted
- Received
- Academic Editor
- Junkuo Gao
- Subject Areas
- Natural Resource Management, Environmental Contamination and Remediation
- Keywords
- Caulerpa lentillifera, Cultivation, Wastewater treatment, Bioactive components
- Copyright
- © 2019 Chen et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. Advances in cultivation, wastewater treatment application, bioactive components of Caulerpa lentillifera and their biotechnological applications. PeerJ 7:e6118 https://doi.org/10.7717/peerj.6118
Abstract
The edible seaweed Caulerpa lentillifera, a powerful natural food source that is rich in protein, minerals, dietary fibers, vitamins, saturated fatty acids and unsaturated fatty acids, has been mass cultured in some Asian countries and has been the focus of researchers in recent years. Here, the operational conditions of its culture, application in wastewater treatment, and bioactive components are summarized and comparatively analyzed. Based on previous studies, salinity, nutrient concentrations, irradiance and temperature are stress factors for algal growth. Moreover, dried Caulerpa lentillifera seaweed is efficient in the biosorption of heavy metals and cationic dyes in wastewater, and fresh seaweed can be introduced as a biofilter in aquaculture system treatment. In addition, among the rich bioactive compounds in Caulerpa lentillifera, the phenolic compounds show the potential ability for regulating glucose metabolism in vivo. Polysaccharides and oligosaccharides exhibit anticoagulant, immunomodulatory effects and cancer-preventing activity. Siphonaxanthin is a compound with attractive novel functions in cancer-preventing activity and lipogenesis-inhibiting effects. Furthermore, the antioxidant activity of siphonaxanthin extracted from Caulerpa lentillifera could be stronger than that of astaxanthin. This review offers an overview of studies of Caulerpa lentillifera addressing various aspects including cultivation, wastewater treatment and biological active components which may provide valuable information for the cultivation and utilization of this green alga.
Introduction
As shown in Fig. 1, Caulerpa lentillifera, green seaweed with high economic value, is naturally distributed in tropical and subtropical regions, such as South China Sea, Southeast Asia, Japan, Okinawa, Taiwan and Oceania (Paul et al., 2014). As reported in literatures, this green seaweed was documented for the first time on Red Sea coast (Agardh, 1837), and then was observed at many other locations, especially in Indo-Pacific region (Hackett, 1977; Taylor, 1977; Menez & Calumpong, 1982; Coppejans & Beeckman, 1990; Phillips, Conacher & Horrocks, 1999; Schils & Coppejans, 2003; Titlyanov, Titlyanova & Pharm, 2012). Because its upright branches resemble grapes, C. lentillifera is also called “sea grapes” (Guo et al., 2015a), and it can grow on sand and rock bottoms in the upper sublittoral zone of tropical coral reefs (Horstmann, 1983; Mao et al., 2011). Because of its good taste, C. lentillifera is often cooked as salad in some Asian countries. In addition, C. lentillifera is rich in polyunsaturated fatty acids (PUFAs) (Saito et al., 2010), multiple essential amino acids, minerals, dietary fibers, vitamin A and Vitamin C (Matanjun et al., 2009) and has low levels of lipids (Niwano et al., 2009). Therefore, there has been increasing demand and rising market prices for C. lentillifera in some Asian countries recently. However, although this alga is widely cultivated in Philippines (Zemkewhite & Ohno, 1999), Okinawa (Kurashima et al., 2003), Taiwan Island (Shi, 2008), Fujian and Hainan provinces in China (Wang, 2011), the commercial-scale production of C. lentillifera is still not sufficient, and its productivity does not meet the demand. It might be due to lack of optimum cultivation conditions of the alga. Therefore, it is important to obtain the best culture conditions to increase the productivity of C. lentillifera.

Figure 1: Caulerpa lentillifera grown in Huang Hai, China (supplied by Xiaolin Chen).
{kind=link}
Currently, the main research of this alga focuses on the treatment of wastewater and development of bioactive components. C. lentillifera has shown potential ability to remove basic dyes from waste streams (Marungrueng & Pavasant, 2006), heavy metals from industrial wastewater (Pavasant et al., 2006; Apiratikul & Pavasant, 2008), and nutrients from aquaculture effluents (Paul & De Nys, 2008), especially NO3-N (Guo et al., 2015b).
In recent years, some bioactive components of C. lentillifera, such as phenolic compounds, polysaccharides and pigments, and their biological potentials, including antioxidant, anti-diabetic and anticancer activities, have been documented. Therefore, in order to further understand and make better use of this seaweed, we summarized researches of its cultivation conditions, wastewater treatment abilities, and bioactive components along with their biological activity.
Methods
Survey methodology
Two main databases were used to obtain related literature including Web of Science and Google Scholar from 1900–2018. The selected references were listed after the acknowledgement. From the previous researches, the results were reviewed.
Cultivation conditions for C. lentillifera
According to previous literatures (shown in Table 1), we concluded that salinity, nutrient concentration, irradiance and temperature were all stress factors for growth during all periods, when these factors change and they will correspondingly affect the physiology of the alga such as growth rate, chlorophyll concentration etc. Therefore, it is important to study the optimum factors for massive culture of the alga. Deraxbudsarakom et al. (2003) suggested that a salinity range of 25–30‰ was suitable for the normal growth of C. lentillifera when the alga was cultured by shrimp farm effluent at laboratory. Wang (2011) showed that the maximum growth of C. lentillifera supplied by Fujian China occured at a salinity of approximately 36‰, which was cultured with filtered seawater added with salt; later, a study by Guo et al. (2015b) confirmed this result. C. lentillifera transported from Okinawa Japan did not survive at salinities of 5‰ and 55‰ cultured by sterile seawater. This study indicated that the specific growth rate (SGR) for C. lentillifera was different among all the groups. The maximum SGR was obtained at a salinity of 35‰, and this result was consistent with the maximum chlorophyll content and the ratio of fluorescence (Fv/Fm). At salinities of 20‰ and 45‰, only stolons regenerated from branches. However, new branches grew from stolons at salinities 30‰–40‰. Therefore, studies suggested that the optimal salinity concentration for the growth of C. lentillifera was between 30‰–40‰.
| Cultivation conditions | Effect | Reference |
|---|---|---|
| Salinity | Suitable salinity range of 25–30‰; the maximum growth at a salinity of approximately 35–36‰ | Deraxbudsarakom et al. (2003), Wang (2011) and Guo et al. (2015b) |
| Nitrogen and phosphorus | Optimal for the rapid growth at 0.6 mmol/L NO3-N and N:P ratio of 8:1; Highest SGR at a 0.1 mmol/L PO4-P and 0.5 mmol/L NO3-N; Nitrogen types (NaNO3 and NH4NO3) can significantly promote the growth of the alga; NH4-N:NO3-N ratios of 1:1 and 1:5 were the most favorable ratios for the growth of the alga | Deraxbudsarakom et al. (2003), Guo et al. (2015b), Wang et al. (2017), Liu et al. (2016) |
| Phytohormones | 6-BA and GA could induce the growth of the alga, but IAA could increase the intracellular crude polysaccharide content | Tao et al. (2017) |
| Temperature | Ideal temperature range 22–28 °C | Friedlander et al. (2006) and Guo et al. (2015a) |
Nitrogen (N) and phosphorus (P) are two essential nutrients for the growth of C. lentillifera and they must be taken from the environment. Deraxbudsarakom et al. (2003) concluded that a 0.6 mmol/L NO3-N concentration and N:P ratio of 8:1 were optimal for the rapid growth of C. lentillifera with salinity 25–30‰. However, Guo et al. (2015b) reported that SGR of C. lentillifera from Okinawa was the highest at 0.1 mmol/L PO4-P concentration and 0.5 mmol/L NO3-N concentration (approximate N:P ratio of 5:1, water temperature 25 °C and light of 40 umol photons/(cm2 s)), which was slightly different from the results of Deraxbudsarakom et al. (2003). In addition to nitrogen concentration, different oxidation states of N also had effects on the biomass production of C. lentillifera. For example, Wang et al. (2017) used four different nutrient salts (NaNO3, NH4NO3, CO(NH2)2 and NH4HCO3) to cultivate C. lentillifera supplied by Ocean University of China with temperature 27 °C, light of 145.45 umol photons/(cm2 s) and salinity 30‰. The results showed that nitrate (NaNO3 and NH4NO3) can significantly promote the growth of the alga. Under a concentration of 20 mg/L NH4NO3, the relative growth rate of the alga was the highest. In addition, Liu et al. (2016) indicated that NH4-N: NO3-N ratios of 1:1 and 1:5 were the most favorable ratios for the growth of the alga. In conclusion, the optimal concentration of NO3-N was 0.1 mmol/L-0.6 mmol/L and the optimal N: P was 5:1-8:1 for the growth of C. lentillifera.
Different phytohormones, such as gibberellin (GA), 6-benzyl aminopurine (6-BA) and indoleacetic acid (IAA), have also been shown to be efficient for the growth of C. lentillifera (Tao et al., 2017). The results (Tao et al., 2017) revealed that 0.8 and 1.4 mg/L 6-BA could induce a relatively high weight gain rate and SGR of C. lentillifera and that 11 mg/L GA was the optimal concentration for rapid growth, while IAA showed no obvious effect on the biomass of C. lentillifera. In addition, compared to GA, which had no significant effect on the production of crude polysaccharides in C. lentillifera, IAA increased the intracellular crude polysaccharide content.
Temperature has a major effect on the kinetics of cellular enzymes, and irradiance is an essential source of photosynthetic activity in algae. Hence, the growth of C. lentillifera is also induced by temperatures and irradiances at certain degrees. A previous study showed that C. lentillifera started to become soft and decay and the productivity of biomass decreased sharply when the temperature reduced to 18 °C. Moreover, Guo et al. (2015a) found that the biomass of the alga reached the maximum of 6.932 ± 0.396% day−1 at 27.5 °C and 40 µmol photons/(m2 s). In addition, the authors also found that higher irradiances (40–100 umol photons/(m2 s)) could decrease the chlorophyll content and rbcL expression. An experiment by Wu et al. (2017) further confirmed that different levels of light quality showed different effects on the growth and photosynthetic pigment contents of C. lentillifera. The concrete results showed that the light treatment of a blue/red ratio of 5/1 had significant beneficial effects on the fresh weight/length ratio, the fresh weight of regenerated vertical branches and the diameter of regenerated spherical ramuli. However, the contents of total chlorophyll, chlorophyll a, chlorophyll b and carotenoids significantly increased under full blue light. A comprehensive analysis suggested that 5/1 for blue/red and full white were suitable for indoor culture of C. lentillifera. In summary, the optimal temperature for the growth of C. lentillifera was about 20–28 °C. And more blue light or full white treatment would be benefit for the cultivation.
Besides the above cultivation parameters, the origin of the alga such as different area might lead to different growth results. However, there was no reference to introduce the research.
With the development of culture research, different applications of C. lentillifera have been studied. And wastewater treatment was early studied.
Wastewater treatment by C. lentillifera
As mentioned in documents, C. lentillifera has been studied as biosorption material to treat wastewater, such as heavy metal wastewater, toxic dye-contaminated wastewater and aquaculture wastewater(shown in Table 2). There are several advantages to apply seaweeds as biosorbent, including their wide availability, low cost, high metal sorption capacity, reasonably regular quality, and relatively simple application. Pavasant et al. (2006) proved the ability of dried C. lentillifera to absorb Cu2+, Cd2+, Pb2+ and Zn2+. Moreover, the removal efficiency of the alga rose with an increased pH 2–8 (temperature 21 ± 2 °C), and the sorption process of all metal ions only took 20 min which was much faster than that of alginate/Mauritanian clay (with diffusion coefficient 4–8 × 10−7 cm2/S; Ely et al., 2011). The sorption of heavy metals on the biosorbents mainly included two steps (Pavasant et al., 2006):
| Alga types | Wastewater types | References |
|---|---|---|
| Dried alga | Cu2+, Cd2+, Pb2+ and Zn2+ | Pavasant et al. (2006) and Apiratikul & Pavasant (2006) |
| Dried alga | Cationic dyes: Astrazon Blue FGRL (AB), Astrazon Red GTLN (AR), and methylene blue (MB) | Marungrueng & Pavasant (2006), Marungrueng & Pavasant (2007), Ncibi, Mahjoub & Seffen (2007), Cengiz & Cavas (2008), Punjongharn, Meevasana & Pavasant (2008) |
| Fresh alga | Used as a biofilter in aquaculture systems for nutrient absorption, especially NO3-N | Paul & De Nys (2008), Liu et al. (2016), Chokwiwattanawanit (2000) |
-
The metal ions were initially taken up onto the surface of the cells;
-
They were bioaccumulated within the cells due to the metal uptake metabolism.
Step 1 involved passive transport, and it took place quite rapidly, i.e., within 20–30 min, while Step 2 took much longer to complete. In this case, the alga was dried and no longer active, so the sorption could only take place on the surface of the cell, which controlled the whole sorption process. Therefore, it took place only 20 min. Furthermore, the sorption process followed the Langmuir isotherm, and the maximum sorption capacities were Pb2+ > Cu2+ > Cd2+ > Zn2+.
In another study, the authors (Apiratikul & Pavasant, 2008) continued to use dried C. lentillifera to study the biosorption process of Cu2+, Cd2+ and Pb2+, and the sorption kinetics best followed the pseudo second-order kinetic model: (1)
In Eq. (1), q (mg/g) is the amount of the metal adsorbed at time t (min), qe (mmolKg−1) is the amount of the metal adsorbed at the time of equilibrium, and k is the equilibrium rate constant. The values for qe, k and R2 were listed in Table 3.
| Parameters | Cu2+ | Cd2+ | Pb2+ |
|---|---|---|---|
| qe (mmol Kg−1) | 6.14 | 3.97 | 2.64 |
| K (Kg mol−1 min−1) | 254 | 621 | 2,036 |
| R2 | 0.999 | 1.000 | 1.000 |
In addition, the sorption isotherm data fit the Langmuir isotherm model: (2)
In Eq. (2), qe represents the amount of metal ion taken up per unit mass of the biomass at equilibrium (mol/kg), qmax is the maximum amount of metal ion taken up per unit mass of the biomass (mol/kg), b is the Langmuir affinity constant (m3/mol), and Ce is the equilibrium concentration of the heavy metal ion in solution (mol/m3). In addition, according to Dubinin-Radushkevich model, the sorption energies are 4–6 kJ/mol, as the process involves a physical electrostatic force. Ion exchange is believed to be a principal mechanism of the sorption, and metal ions such as Ca2+, Mg2+ and Mn2+ are the main ions released from the algal biomass. In addition, the binary component systems composed of Cu2+, Cd2+ and Pb2+ were also studied for the sorption of dried C. lentillifera. The experimental data was effectively described by the partial competitive binary isotherm model. In addition, the secondary metal ion always reduced the total sorption capacity of the previous metal ions, which implied that the concomitant metal ions competed for the same pooled binding sites during the algal biomass sorption process, and Pb2+ was the most adsorbed metal ion according to the study. The batch scale experiments by fixed bed column also showed that sorption capacities for various metals could also be prioritized with the same order: Pb2+ > Cu2+ > Cd2+. These results were beneficial for the further design and scaling up of the system (Apiratikul & Pavasant, 2008; Apiratikul & Pavasant, 2006).
Dried C. lentillifera has also been utilized to treat cationic dyes, which are widely used in the textile industry, because dried C. lentillifera contains many functional groups (O-H, COOH, NH2 and S=O) that exhibit chemical binding affinity toward several positively charged ions, and these characteristics might also be showed by other algae. Overall, dried C. lentillifera was proved to effectively absorb Astrazon Blue FGRL (AB), Astrazon Red GTLN (AR), and methylene blue (MB). The maximum sorption capacity of MB was 417 mg/g which was greater than that of active carbon (Marungrueng & Pavasant, 2007). Some parameters, including the initial dye concentration, pH, temperature, salinity, alga size and dosage, have important effects on the sorption process. In concrete, the adsorption rate constants increased with a decrease of the initial dye concentration. At low dye concentrations (20–80 mg/L), the application of an increasing amount of the alga resulted in a higher percentage of the removed dye(more than 95%) but a lower amount of the dye adsorbed per unit mass (Marungrueng & Pavasant, 2006). For MB adsorption, pH of 7–11 might be appropriate because this pH range can supply advantageous surface binding sites of the alga for the ionization of the dye molecule (Ncibi, Mahjoub & Seffen, 2007). Marungrueng & Pavasant (2006) reported that high temperatures, such as 70 °C, could reduce the adsorption of FGRL, while the maximum adsorption capacity was obtained at 50 °C (qm for langmuir was 49.26 mg g−1). In terms of alga size, a small size of 0.1–0.84 mm resulted in the highest adsorption capacity, followed by intermediate (0.84–2.0 mm) and larger sizes (larger than 2.0 mm) because the small size provided the most surface area and total pore volume for the adsorption of the dye. Additionally, salinity was another stress factor in the system, and high salinity caused a decrease in adsorption capacity due to the competition between Na+ and the dye cations for the binding sites on the algal surface and electrical repulsion (Punjongharn, Meevasana & Pavasant, 2008). Furthermore, pseudo second-order kinetic model and Langmuir model could describe the kinetic adsorption and adsorption isotherms process well, respectively (Punjongharn, Meevasana & Pavasant, 2008; Marungrueng & Pavasant, 2006; Cengiz & Cavas, 2008). The sorption process is controlled by both film and pore diffusion (Marungrueng & Pavasant, 2007).
As a method of wastewater treatment, dried C. lentillifera can adsorb heavy metals and dyes, and fresh C. lentillifera can be used as a biofilter in aquaculture systems because it has a significant capacity for nutrient absorption, especially that of NO3-N (Paul & De Nys, 2008; Liu et al., 2016). C. lentillifera was successfully applied at a hatchery scale to a recycling aquaculture system for juvenile spotted babylons (Babylonia areolata), and the results revealed that it had a positive effect on the survival rate of spotted babylons, seawater quality and the biomass of C. lentillifera (Chaitanawisuti, Santhaweesuk & Kritsanapuntu, 2011). In addition, it has often been cultured in shrimp ponds used as water treatment methods (Chokwiwattanawanit, 2000).
Besides the application in wastewater treatment, like other algae, bioactive components of C. lentillifera and their bioactive potentials have also been studied in recent years.
Bioactive components of C. lentillifera and their biological potentials
C. lentillifera contains abundant proteins (10.41% DW (dry weight)), PUFAs (polyunsaturated fatty acids, 16.76% total fatty acids), and total dietary fiber (32.99% DW) (Matanjun et al., 2009; Nagappan & Vairappan, 2014), and the alga is also rich in some bioactive components (shown in Table 4).
| Components | Biological activity | References |
|---|---|---|
| Phenolic compounds | Radical-scavenging activity and reducing power ability; Stimulated insulin secretion in pancreatic β-cells and enhanced glucose uptake | Matanjun et al. (2008), Nguyen, Ueng & Tsai (2011), Sharma & Rhyu (2014), Sharma, Kim & Rhyu (2017), Sharma, Kim & Rhyu (2015), Abouzid et al. (2014) |
| Polysaccharides | Increase the phosphorylation of p38 MAPK; Inhibit the proliferation of MCF-7 | Maeda et al. (2012a); Maeda et al. (2012b) |
| Siphonaxanthin | cancer-preventing action; Inhibit adipogenesis; | Ganesan et al. (2011); Li et al. (2015); Zheng et al. (2018) |
The total contents of phenolic compounds of dried C. lentillifera differed due to the climate and environment in which the alga grew (Ito & Hori, 1989). Nguyen, Ueng & Tsai (2011) reported that the total phenolic content of thermally dried and freeze-dried C. lentillifera were 1.30 mg and 2.04 mg gallic acid equivalent (GAE)/g of dry weight, respectively, which were significantly lower than the data reported by Matajun (30.86% of dry weight; Matanjun et al., 2008). As reported in the literature, the phenolic compounds of C. lentillifera are often extracted using ethanol, methanol or diethyl ether and show different biological activities. The methanolic and diethyl ether extracts showed better radical-scavenging activity (2.16 mM/mg dry extract by TEAC method) and reducing power ability (362.11 uM/mg dry extract by FRAP method) than those in other brown and red seaweeds (1.63 mM/mg dry extract by TEAC method and 225.00 uM/mg dry extract by FRAP method for Eucheuma cottonii; 1.66 mM/mg dry extract by TEAC method and 268.86 uM/mg dry extract by FRAP method) (Matanjun et al., 2008). The ethanol extracts had strong hydrogen peroxide-scavenging activity (94.81% with 60 ppm) and weak DPPH-scavenging (IC50 was greater than 100 ppm), weak ferric ion-reducing activity (1.93–1.94 ug ascorbic acid equivalent/ml for 20 ppm extract) and weak FIC activity (not exceeding 70% with100 ppm) (Nguyen, Ueng & Tsai, 2011). In addition, the ethanol extracts also stimulated insulin secretion in pancreatic β-cells and enhanced glucose uptake by decreasing dipeptidyl peptidase-IV, α-glucosidase and protein-tyrosine phosphatase 1B activities using RIN and 3T3-L1 cells as models (Sharma & Rhyu, 2014; Sharma, Kim & Rhyu, 2017) and regulated glucose metabolism via the PI3K/AKT signaling pathway in myocytes using L6 cells (Sharma, Kim & Rhyu, 2015; Abouzid et al., 2014), which could ameliorate insulin resistance.
Polysaccharides are important components of C. lentillifera due to their broad spectrum of biological activity. The crude extract of C. lentillifera showed anticoagulant property using albino rabbits and the blood of adult dogs. And it exhibited approximate effect of aspirin (Arenajo et al., 2017). Shevchenko et al. (2009) extracted three polysaccharide fractions, water-soluble P1, P2 and base-soluble P3. The molecular weights of these polysaccharides were 20–60 KDa, 20–40 KDa and more than 70 KDa, respectively. All of the monosaccharide components of these three factions included glucose (Glc), galactose (Gal), mannose (Man) and xylose (Xyl); among these components, glucose was the majority monosaccharide. Moreover, IR spectra of the polysaccharides indicated that the three fractions lacked sulfated groups. However, these results were not inconsistent with those from another study of Maeda et al. (2012a), which reported that the purified polysaccharides (SP1) contained sulfated xylogalactan with a molecular mass >100 KDa. This xylogalactan is mainly composed of galactose, xylose and small quantities of glucose and uronic acid, with 44% sulfation. Furthermore, the SP1 could enhance NO production and activate macrophage cells via NF-κB and increase the phosphorylation of p38 MAPK, which indicates that they can activate RAW 264.7 cells. In another report, β-1,3-xylooligosaccharides could inhibit the proliferation of MCF-7 human breast cancer cells and induce the condensation of chromatin, the degradation of PARP, and the activation of caspase-3/7, which indicates that oligosaccharides can induce apoptosis in MCF-7 cells (Maeda et al., 2012b).
Recently, valuable pigments are attracting increasing attention because of their important biological activity. Worth mentioning is siphonaxanthin, a novel and oxidative metabolite of lutein, which is found in C. lentillifera. As shown in Fig. 2, its structure contains a conjugated system of 8 C=C double bonds and 1 keto group located at C-8, similar to fucoxanthin. In addition, at the C-19 position, siphonaxanthin has an extra hydroxyl group, which might make it more beneficial than other carotenoids (Ganesan et al., 2011; Walton, Britton & Goodwin, 1973).
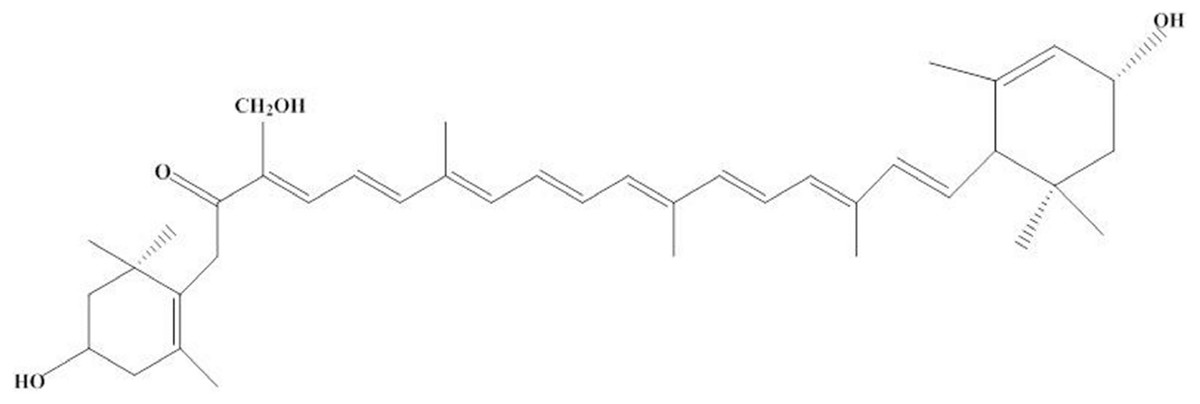
Figure 2: Structure of siphoxanthin.
{kind=link}
Siphonaxanthin is a specific keto-carotenoid that mainly exists in green algae, such as Codium fragile, C. lentillifera, Umbraulva japonica, and Caulerpa racemosa. The content of siphonaxanthin is approximately 0.03%–0.1% of its dry weight (Sugawara et al., 2014). Initially, this keto-carotenoid was proved to facilitate the highly efficient energy transfer of carotenoids to chlorophylls (Akimoto et al., 2008). Moreover, it might have a largely light-harvesting function in the green light-rich underwater habitat to reduce light damage (Wang et al., 2013). In addition to its physiological functions, siphonaxanthin has been found to show many biological activities. It was involved in cancer-preventing action in human leukemia HL-60 cells by increasing in TUNEL-positive cells and chromatin condensation in the cells by decreasing the expression of Bcl-2 but up-regulating the expression of DR5. Furthermore, the anticancer activity of siphonaxanthin was stronger than that of fucoxanthin and siphonein which is an esterified form of siphonaxanthin (Ganesan et al., 2011). In addition, siphonaxanthin can show antiobesity effect by inhibiting adipogenesis in 3T3-L1 preadipocytes and lipid accumulation in the white adipose tissue of KK-Ay mice and inhibiting protein kinase B phosphorylation and regulating the expression of CEBPA (enhancer binding protein α), PPARG (peroxisome proliferator activated receptor γ), FABP4 (fatty acid binding protein 4) and SCD1 (stearoyl coenzyme A desaturase 1) (Li et al., 2015). Zheng et al. (2018) found that siphonaxanthin can inhibit lipogenesis in hepatocytes by suppressing the excess accumulation of triacylglycerols induced by liver X receptor α agonist and down-regulating nuclear transcription factors with HepG2 cell line.
Subheadings
-
Salinity, nutrients concentration, irradiance and temperature were the most important factors to influence Caulerpa lentillifera growth.
-
Dried seaweed could be used as biosorbent for heavy metals and cationic dyes, and fresh seaweed could be biofilter for the aquaculture system.
-
The phenolic compounds showed good antioxidant activity and could regulate glucose metabolism.
-
Polysaccharides and oligosaccharides exhibited immunodulatory effects and cancer-preventing activity.
-
Siphonaxanthin as a novel function compound showed cancer-preventing activity and lipogenesis inhibiting effect.
-
In conclusion, C. lentillifera need be further studied for more functions such as antiviral, anti-inflammatory areas.
Conclusion
The green seaweed C. lentillifera is quite common and popular in Southeast Asian countries and Japan due to its delicious taste and abundant nutrients. During the past 30 years, it has been mass cultivated in some Asian countries, such as the Philippines and Malaysia. And some cultivation conditions, such as the nutrient concentration, salinity, irradiance and temperature, have been studied in relation to the growth of C. lentillifera. In addition, this species has been applied to treat wastewater using heavy metal, cationic dye biosorption and aquaculture system. Recently, some bioactive components, such as phenolic compounds, polysaccharides, and siphonaxanthin, have been extracted from C. lentillifera, and their biological potentials have also been analyzed by cells. In conclusion, these compounds showed high antioxidant, anticoagulant and immunostimulatory, hypoglycemic, cancer-prevention and lipogenesis inhibition activities, etc. in vitro. It is believed that this seaweed will be a new source of health products with its cultivation at an increasing scale. In addition, perhaps C. lentillifera will be used as the resource of biofuel or CO2 fixation just like other algae with further research.