Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities †


1
USDA, ARS, New England Plant, Soil, and Water Laboratory, University of Maine, Orono, ME 04469, USA
2
Department of Plant Pathology, University of Georgia, Athens, GA 30602, USA
*
Author to whom correspondence should be addressed.
†
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
‡
Portions of this work were a part of Marin T. Brewer’s master thesis.
Agriculture 2020, 10(4), 128; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10040128
Submission received: 12 March 2020
/
Revised: 7 April 2020
/
Accepted: 9 April 2020
/
Published: 14 April 2020
(This article belongs to the Section Crop Protection, Diseases, Pests and Weeds)
Abstract
:Rotation crops and biocontrol amendments were investigated for suppression of Rhizoctonia solani on potato (Solanum tuberosum) and their interactive effects on soil microbial communities. Greenhouse trials were conducted to evaluate selected rotation crops, including barley, common and “Lemtal” ryegrass, clover, potato, and combinations of barley with ryegrass or clover, for their effects on populations of R. solani and Rhizoctonia disease. Potato and clover preceding potato resulted in higher disease severity than most other rotations, whereas ryegrass reduced stem canker severity. In addition, all ryegrass treatments resulted in substantially higher populations of R. zeae. Field trials evaluating selected biocontrol treatments in combination with different rotations were conducted at two locations in Maine. Potatoes were treated with the biocontrol organisms Laetisaria arvalis, Trichoderma virens, or Bacillus subtilis and planted following rotation crops of barley and ryegrass, barley and clover, or potato. The barley/ryegrass rotation significantly reduced incidence and severity of stem canker and increased tuber yield at one location. Efficacy of the biocontrol treatments varied by rotation and location, with L. arvalis and T. virens reducing black scurf in some rotations and increasing some aspects of tuber yield at one location. Soil microbial community characteristics differed among rotation crops and biocontrol treatments. Significant crop by biocontrol interactions were observed demonstrating the complex interactions among rotation crops, biocontrol treatments, and soil microbial communities, as well as indicating that biocontrol can be enhanced within beneficial rotations.
1. Introduction
Rhizoctonia solani Kühn is a persistent pathogen of potato (Solanum tuberosum L.) that causes stem and stolon canker and is also responsible for black scurf, a tuber disease caused by the formation of sclerotia, the long-term survival structures of the pathogen, on newly formed tubers [1,2,3,4]. These symptoms result in reduced tuber quality and yield. Rhizoctonia solani is classified into numerous discrete Anastomosis Groups (AGs), and AG-3 is the predominant AG responsible for Rhizoctonia disease of potato in the U.S. and throughout most of the world [1]. Current cultural and chemical controls, including crop rotation, reduced time to emergence, and fungicide seed treatments, are not always practical or effective, and Rhizoctonia disease of potato continues to be a serious problem wherever potatoes are grown [1,2,3,4].
Effective, alternative disease control options that are consistent with sustainable production practices are needed. Improved crop rotations and biocontrol organisms represent an integrated approach that may effectively reduce Rhizoctonia disease, improve yield, and contribute to agricultural sustainability and environmental quality. Crop rotations can suppress disease by allowing inoculum levels to decline in the absence of the host, directly inhibiting the pathogen by producing a toxic compound, increasing specific antagonists that inhibit the pathogen, or by increasing general microbial populations that compete with the pathogen [5,6,7]. Crop plants are primary drivers of changes in soil microbial communities and may be responsible for changes that increase soil microbial activity, diversity, populations of plant-beneficial organisms, and antagonism towards pathogens, potentially resulting in disease suppression and improved yield [8,9,10]. Previous research in potato systems has demonstrated that each type of rotation crop produces distinctive changes in soil microbial community characteristics, as well as that these characteristics may be related to disease characteristics [6,11,12].
Presently, barley (Hordeum vulgare L.) or oats (Avena sativa L.) underseeded with a cover crop of red clover (Trifolium pratense L.) in a 2-year rotation with potato is a standard rotation practiced on many commercial farms in Maine [13]. Barley rotations can suppress Rhizoctonia diseases to some degree [6,11,14]; however, more effective cover crops may be available for use in conjunction with barley [14]. Legume rotations have been implicated in decreased yields of subsequent crops, mainly as a result of increases in disease by R. solani [6,14,15]. Observations in the field have suggested that ryegrass (Lolium multiflorum Lam.), and possibly a particular variety known as “Lemtal”, may be more effective as a cover crop with barley for suppressing stem canker and black scurf, but this has not been sufficiently documented and tested.
Another potentially sustainable method of disease suppression is the addition of microbial antagonists. Many biocontrol organisms have shown suppressive activity towards R. solani in various pathosystems [16,17,18], as well as on potato [19,20,21], but disease control has not been consistent. Previously, Brewer and Larkin [22] evaluated many of these organisms against Rhizoctonia disease on potato in greenhouse trials and observed strains of the rhizosphere bacteria Bacillus subtilis and the fungi Trichoderma virens and Laetisaria arvalis to be among the most effective biocontrol organisms tested, reducing stem canker and black scurf by 30% to 60%. In subsequent trials, commercially available formulations of B. subtilis and T. virens provided some control of Rhizoctonia disease of potato in the field [23,24]. One strategy to improve disease control is to combine the use of biocontrol organisms with more effective crop rotations [22,25,26]. Different crop rotations and biological control treatments have been shown to have significant and distinctive effects on soil microbial communities [5,11,14,27,28]. These changes may include overall increases in microbial populations, population changes within specific groups of organisms, or changes in the structural and functional characteristics of the soil microbial communities. In addition, there may be significant interactive effects of the rotations and treatments on each other, as well as on the soil microbial communities and disease relationships. Biological control may be enhanced or reduced depending on the specific rotation [28].
Another important soilborne potato disease that occurs wherever potatoes are grown is common scab, caused by the actinomycete Streptomyces scabies [4]. Common scab also results in tuber surface lesions that can reduce tuber quality and yield [4]. Several biocontrol organisms have also shown activity against this pathogen [29,30] Although the focus of this investigation is on Rhizoctonia disease, because common scab routinely occurs in Maine potato fields, potential treatment effects on common scab will also be evaluated.
Traditionally, soil dilution plating on artificial media was used to estimate populations of different components of soil microbial communities, but this approach has many limitations, such as only measuring the small proportion of microbes that are culturable on media, generally not distinguishing active organisms from dormant propagules, and not assessing soil microbial communities as a whole [31]. Community level approaches that include the full range of soil microbes are needed for assessing the changes and effects on soil microbial communities. The use of whole soil fatty acid methyl ester (FAME) analysis, which is based on profiling the array of fatty acids produced by soil microorganisms, is one such approach that accounts for a much broader array of microbes, is very sensitive in detecting changes in soil microbial communities, and is a relatively quick and simple technique to compare and evaluate soil microbial communities [6,27,32]. Although this technique cannot identify the specific taxa composition of the microbial communities, it can effectively show changes in community structure associated with different cropping systems and the addition of biological amendments [11,14,27,28,33,34].
The purpose of this research was to assess whether the integrated use of a specific crop rotation, barley and ryegrass, and selected biocontrol organisms could effectively reduce Rhizoctonia disease of potato in the field, as well as to evaluate their interactions and effects on soil microbial communities. The biocontrol organisms studied were T. virens GL-21, L. arvalis ZH-1, and B. subtilis GBO3.
2. Materials and Methods
2.1. Inoculum Preparation
Inoculum of Rhizoctonia solani RS31B, a virulent isolate of AG-3 recovered from a potato plant in Maine [22], was used for all experiments. Inoculum was prepared by transferring four plugs of potato dextrose agar (PDA, Difco Laboratories, Inc., Sparks, MD, USA) containing 7-to 14-day-old cultures of R. solani to petri dishes filled with 20 to 30 g of sterile organic cracked wheat. The grain was prepared by adding 3 mL deionized water per 5 g of organic cracked wheat and autoclaving for 60 min on two consecutive days [22]. The inoculated wheat was incubated at room temperature (21 to 25 °C) in the dark for 8 days, then air dried in a sterile hood for 48 h, passed through a 2.0 mm sieve, and stored in a paper bag at 4 to 5 °C for no more than 1 month, until needed. Viability of inoculum was confirmed by plating on PDA.
2.2. Greenhouse Rotation Trials
Crop treatments tested in the greenhouse experiments included two varieties of annual ryegrass (“Lemtal” and a generic ryegrass just referred to as common), barley (“Robust”), red clover (“Cinnamon”), barley and Lemtal ryegrass together, barley and red clover together, and potato. Potato variety “Shepody” was used for all experiments, as it is susceptible to multiple soilborne diseases, such as Rhizoctonia stem canker and black scurf, and common scab. Field soil from Newport, ME, a Nokomis sandy loam (coarse-loamy, mixed, frigid Typic Haplorthod), was used for these experiments. The soil was sieved through a 6 × 6 mm screen and combined with sterile sand at a 3:1 w/w soil to sand ratio. Pathogen inoculum was incorporated with the soil mix at a rate of 4 g per kilogram and incubated for 24 h at room temperature.
Two kilograms of pathogen-infested soil mix was added to each 25 cm × 17 cm × 8 cm plastic potting tray and trays were seeded in the following amounts for each respective rotation treatment: annual ryegrass, 0.67 g; barley, 4 g; clover with Nitro-Fix clover/alfalfa inoculant (Trace Chemicals LLC, Pekin, IL, USA), 0.5 g; and potato, four cut seedpieces treated for 2 min in 2% formaldehyde [22] and green-sprouted for two weeks [35]. These seeding rates were chosen because they resulted in dense growth of each crop. Rates used in the greenhouse studies were several times higher than those used in commercial fields in order to maximize short-term cropping effects. The experimental design was a randomized complete block with four replications per treatment (each potting tray was a replicate), and the experiment was repeated following the same protocols.
Crops were grown in the greenhouse for approximately one month under normal greenhouse conditions (sunlight, regular watering, 15–27 °C), and the treatments containing barley, ryegrass, or clover were cut to soil level and replanted to keep crop growth active. All crops were grown for approximately one more month so that the length of active crop growth was comparable to that of field crops in Maine. Crop biomass from all treatments was cut and incorporated into the soil by hand and, approximately one month later, four potato seedpieces, treated as described above, were planted and grown for approximately 3 weeks.
Soils were sampled after infestation with R. solani, after two months of crop growth, and one month after crop incorporation to determine the effects of crop growth and incorporation on populations of R. solani. Six cores (8 cm deep × 2 cm diameter) were taken and combined into one composite sample for each potting tray (four potting trays per treatment). Samples were plated on five plates of semi-selective media [36] amended with neomycin sulfate (700 mg/L), pyroxychlor (90 mg/L), and benomyl (2 mg/L) using the soil pellet sampler method [37]. Plates were incubated at room temperature for 24 h, and Rhizoctonia-like mycelia emanating from the soil pellets were enumerated by observation at 10× to 20× magnification using a dissecting microscope. The total number of R. solani propagules per gram of soil was calculated from the percent of pellets colonized. This number was subjected to the multiple colonization correction formula [38].
After 3 weeks of growth, total plant emergence was noted and all potato plants were harvested, washed, and rated for stem canker on a scale of 0 to 5 (0 = no disease symptoms; 1 = brown discoloration of stems; 2 = cankers covering <25% of the stem circumference; 3 = 25 to 75% coverage by cankers; 4 = >75% coverage by stem cankers; and 5 = stem completely nipped off or death of the plant). Differences in potato shoot size were detected, so shoots were also rated for size on a scale of 0 to 3 (0 = no growth; 1 = shoot height <2 cm; 2 = shoot height of 2 to 5 cm; and 3 = shoot height >5 cm).
2.3. Field Set-Up and Design
The field experiment was conducted over two field seasons on research plots in Newport (location 1) and Presque Isle (location 2), Maine. The Newport site is located in central Maine (N 44°52′, W 69°17′) and the Presque Isle site is in northern Maine (N 46°38′, W 68°00′), approximately 300 km northeast of the Newport site. Soil at the Newport site is a Nokomis sandy loam (coarse-loamy), and soil at the Presque Isle site is a Caribou sandy loam (fine-loamy), and both are mixed, frigid Typic Haplorthods. The Newport site had previously been planted to millet, ryegrass, or was fallow the previous 5 years, with potatoes grown within the past 10 years. The Presque Isle site had been in sod the previous 3 to 9 years, but it also had potatoes grown within the past ten years. All procedures were conducted similarly at the two locations, except that planting, harvesting, and sampling was approximately one week later for location 2 than location 1. The experimental design was a randomized complete block split-plot with four replicate plots per treatment. Three rotation crop treatments, barley underseeded with “Lemtal” ryegrass, barley underseeded with red clover, and potato, were planted in early June of the first year and followed by potato planted in early June of the second year. The main plots (18 m × 3.6 m) contained the rotation crops, and the subplots (4.5 m × 3.6 m, four potato rows each) consisted of three different biocontrol treatments applied to the potato crop and a control plot with no biocontrol amendment. Prior to planting the rotation crops, R. solani inoculum was added to all field plots at a rate of 64 kg/ha and incorporated into the soil approximately 15 cm by disk or harrow.
Barley was planted at a rate of 135 kg/ha with 45 kg N/ha added using a commercial 10-10-10 fertilizer. The cover crops, clover and ryegrass, were planted with the barley at rates of 17 kg/ha and 23 kg/ha, respectively. Nitro-Fix clover/alfalfa inoculant (Trace Chemicals LLC) was added to the clover seed prior to planting. The potatoes were hand-planted with 35 cm spacing within rows and 0.9 m spacing between rows and fertilized at a rate of 169 kg N/ha. Imidacloprid (Bayer CropScience, Research Triangle Park, NC, USA) and chlorothalonil (Syngenta Crop Protection, Greensboro, NC, USA) were applied to potato plots at recommended rates as needed for control of Colorado potato beetle and late blight (Phytophthora infestans), respectively. Crops were managed and cultivated using recommended production practices.
The barley was harvested in early fall, with the cover crops allowed to overwinter and then incorporated by chisel plow the following spring one month prior to potato planting. Biocontrol treatments, including Bacillus subtilis GBO3 (Kodiak, Bayer CropScience, Research Triangle Park, NC, USA), Trichoderma virens GL-21 (SoilGard, Certis USA, Columbia, MD, USA), or Laetisaria arvalis ZH-1 (ATCC #62715), were added by hand as in-furrow amendments to the potato crop at planting. No organisms were added to the control plot. Potato plots were cultivated and maintained as previously described. In early October, six linear meters of tubers were harvested by hand from the middle two rows of each plot.
2.4. Biocontrol Treatment Preparation
B. subtilis inoculum was prepared by adding deionized water to plates of the isolate grown on 0.1% tryptic soy agar (Difco Laboratories, Franklin Lakes, NJ, USA). The slurry was transferred to 4 L of trypticase soy broth (Difco Laboratories) and incubated on a platform shaker for three days at 28 °C. The broth was added to 6 L of deionized water, and the preparation was added in-furrow at a rate of approximately 104 CFU/cm3 soil (109 CFU/linear meter). T. virens was added in-furrow as the SoilGard formulation (Certis USA), as directed by the manufacturer at a rate of 0.1% (g/cm3). Four plugs of L. arvalis cultures actively growing on PDA were added to petri dishes of sterile cracked wheat, prepared as mentioned earlier, and grown at 26 °C for 5 days. The inoculum was dried at room temperature (21 to 25 °C) for 24 h and added at a 0.1% (g/cm3) incorporation rate. All treatments were used in the field immediately after preparation.
2.5. Disease and Yield Assessments
Shoot emergence was determined by counting the number of plants visible in the middle two rows of each four-row plot over several weeks in late June and early July. Stem canker was assessed in mid-August. Two plants were harvested from each of the first and fourth rows of every plot, and the stems of the plants were rinsed with water to remove soil so that cankers would be visible. Stems were rated on a scale of 0 to 5, as described for the greenhouse trials. When two or more shoots emerged from one seed piece, the average rating of all shoots was recorded.
After harvest, tubers were washed and graded into three categories based on size, and total and marketable yield were determined. The categories consisted of small (<4.7 cm), medium (4.7 to 5.6 cm), and large (>5.6 cm) tuber sizes. Marketable yield included the medium and large size classes. In addition, the percentage of misshapen tubers (by weight) was determined. Thirty tubers from the marketable yield category in each plot were arbitrarily chosen and rated for black scurf and common scab, if present, based on the percent surface area of each tuber covered by sclerotia or lesions.
2.6. Soil Rhizoctonia Populations
Initial soil samples were taken prior to and after the addition of R. solani inoculum in May, in late-June after emergence of the crops, and in mid-August in year 1. In the following year (year 2), soil samples were taken in May prior to incorporation of the cover crops, in June after incorporation and immediately prior to potato planting, and in mid-August during the potato growing season. Samples consisted of 10 cores (15 cm deep by 2 cm diameter) taken from each plot and combined into one composite sample per plot. Samples were sieved (3.35 mm mesh) and kept at 5 °C until processed. Inoculum levels of R. solani were determined for all soil samples taken in year 1 and the two samples taken prior to planting in year 2 by the soil pellet sampler method [37], as described previously.
2.7. Analysis of Microbial Communities Using FAME Profiles
Whole soil fatty acid methyl ester (FAME) analysis was used to evaluate changes and differences in soil microbial communities. This analysis was performed on the initial soil samples of year 1 to obtain base level fatty acid data. The two sets of samples taken prior to planting in year 2 were analyzed to observe changes in the microbial communities prior to and after incorporation of crop residues. The mid-August soil samples from year 2 were also assayed to determine effects of both previous rotation crops and biocontrol organisms on soil microflora. Fatty acids were extracted from three soil subsamples per plot (4 g per subsample, four plots per treatment) according to a modification of the Microbial Identification System (MIS, MIDI, Inc, Newark, DE, USA), as previously described by Larkin [23]. Fatty acid composition was determined by gas chromatography using an automated procedure by MIDI on an HP 6890 gas chromatograph (Hewlett-Packard, Wilmington, DE, USA). Fatty acids were identified by software developed for the MIS. Only fatty acids that accounted for at least 0.25% of the total fatty acid content were used for analysis [27]. In addition, dicarboxylic acids and those with a chain length greater than 20 carbons were not included in the analyses. With these criteria, analyses consisted of 41 to 43 unique fatty acids. Fatty acids were also categorized by structural classes, including saturated straight chain, monounsaturated, polyunsaturated, branched, and hydroxy fatty acid classes. These classes and select individual fatty acids were used as indicators (biomarkers) for particular microorganism groups [6].
2.8. Statistical Analysis
All significant treatment effects were detected using analysis of variance and means were separated using Fisher’s least significant difference at p = 0.05. Fatty acid data was analyzed by principal components analysis (PCA) and canonical variates analysis (CVA), which maximizes group differences [39]. Principal components and canonical variates were subjected to multivariate analysis of variance and one-way ANOVA was run on individual components and key fatty acid groups. SAS (ver. 9.4, SAS Institute, Cary, NC, USA) general linear models procedures were used to carry out statistical analyses.
3. Results
3.1. Greenhouse Rotation Trials
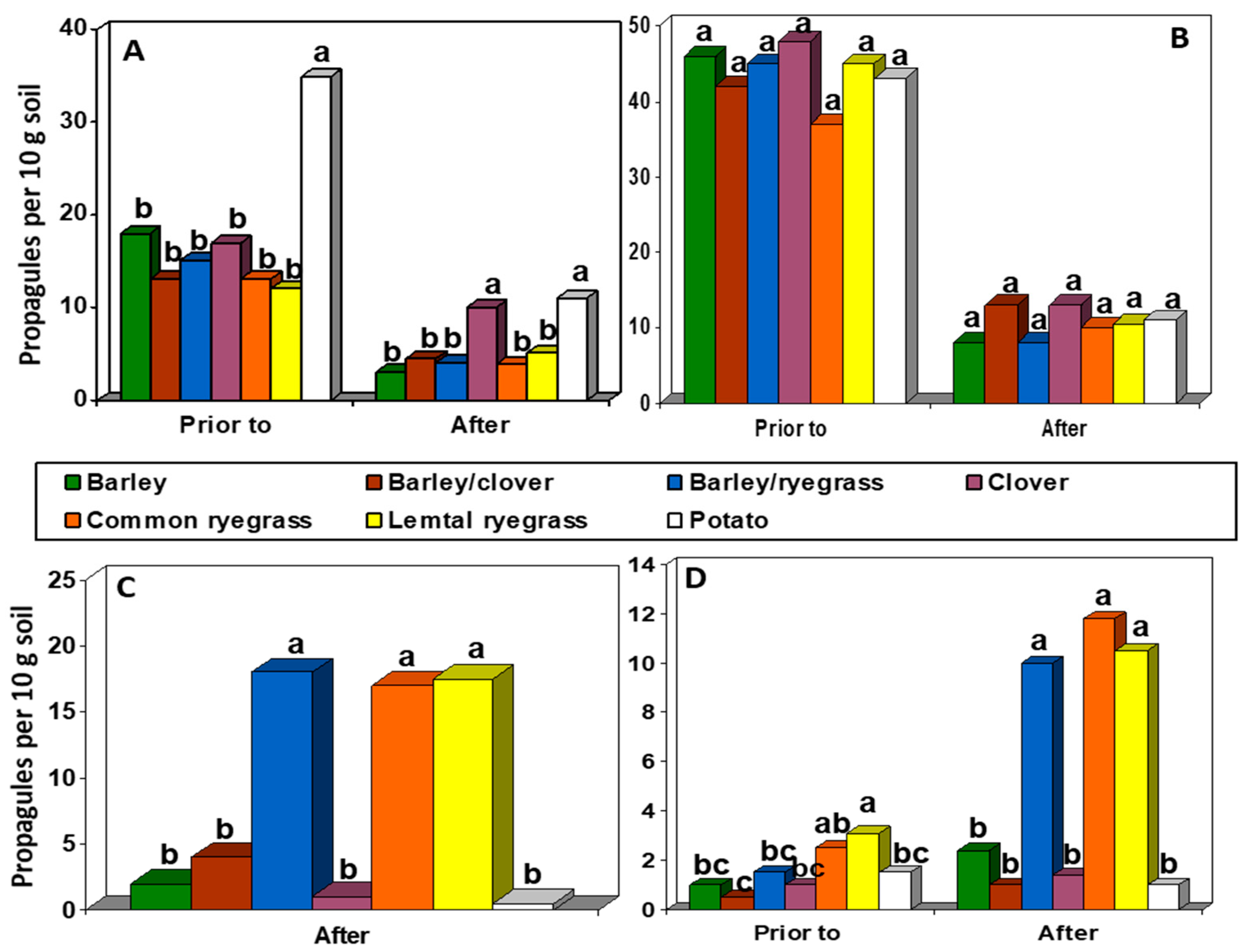
The effects of different rotation crops on soil populations of R. solani and Rhizoctonia disease of potato were examined under controlled conditions in two greenhouse experiments. Initial populations of R. solani, after infestation, were approximately 250 propagules per 10 g soil in both experiments. Populations of R. solani were substantially lower after two months of crop growth and even lower after crop residue incorporation in both experiments (Figure 1). In experiment 1, the potato rotation resulted in higher populations of R. solani than all other rotations prior to residue incorporation. After residue incorporation, populations were higher in both clover and potato than all other rotations (Figure 1A). In experiment 2, there were no differences among crops at either sampling time (Figure 1B).
In addition to R. solani, another different Rhizoctonia sp. was also observed. This organism was identified as R. zeae based on the size, shape, and color of mycelia and sclerotia [40]. R. zeae was detected in soil from most rotations. In both experiments, populations of R. zeae were much greater after residue incorporation in rotations containing ryegrass (barley/ryegrass, Lemtal ryegrass, and common ryegrass) than the other rotations (Figure 1C,D). In experiment 2, although there were some differences in population levels of R. zeae among rotations prior to residue incorporation, differences were much more pronounced after incorporation (Figure 1D).
Stem canker severity of potato shoots also varied based on the previous rotation crop. The clover and potato rotations had higher disease levels than barley, barley/clover, and Lemtal ryegrass rotations in experiment 1 and higher disease levels than the barley/ryegrass, Lemtal ryegrass, and common ryegrass rotations in experiment 2 (Table 1). When averaged over both experiments, the potato and clover rotations resulted in higher disease severity (1.47 for potato and 1.52 for clover) than all other rotations (0.90 to 1.10). In addition, the barley/clover and common ryegrass rotations showed lower incidence of stem discoloration than the potato rotation in experiment 1, with the Lemtal ryegrass rotation reducing incidence relative to potato in experiment 2 (Table 1). The incidence of stem canker was lower in the barley, barley/ryegrass, and common ryegrass rotations in experiment 1, and the barley/ryegrass and common ryegrass rotations in experiment 2, relative to the clover rotation (Table 1).
There were also significant differences in potato shoot size as determined by preceding rotation crop in both experiments. In experiment 1, barley/clover, both ryegrass treatments, and barley/ryegrass rotations yielded larger shoots than the clover rotation (Table 1). Barley/clover and both ryegrass rotations also yielded larger shoots than the potato rotation. In experiment 2, the barley/clover, barley/ryegrass, Lemtal ryegrass, and potato rotations all yielded larger shoots than the clover rotation.
3.2. Field Trial Emergence and Disease Ratings
Potato shoot emergence four weeks after planting was comparable among all rotations at both field locations, but overall emergence was lower at location 1 than location 2 (average 79.6 and 94.1% for location 1 and 2, respectively). At location 1, emergence was greater for the L. arvalis treatment than T. virens (Table 2), whereas emergence was comparable among biocontrol treatments at location 2, averaging 92 to 97%.
Differences in stem canker were detected among rotation crops at location 1. Incidence of stem canker was significantly lower in both barley rotations than in continuous potato, and stem canker severity was significantly lower in the barley/ryegrass rotation than the potato rotation (Table 2). However, no differences in stem canker were detected among rotation crops or biocontrol treatments at location 2 (severity values of 1.6–1.7 and incidences of 53 to 56% among all treatments). There was no rotation by biocontrol interaction for stem canker at either location.
Incidence and severity of black scurf was higher at location 1 than location 2, with uniformly low disease levels at location 2 (severity 0.2 to 0.3% tuber surface covered with sclerotia, incidence 15 to 21%) and no differences detected among rotation crops or biocontrol treatments. At location 1, there were significant rotation crop by biocontrol interactions for black scurf incidence and severity. Although within the potato rotation there were no differences among biocontrol treatments, within the barley/ryegrass rotation, black scurf severity was lower with each of the biocontrol treatments than in the control and lower in the L. arvalis treatments than in the T. virens treatments (Table 3). The incidence of tubers with severe black scurf (coverage of 2% or greater) was also reduced by all three biocontrol treatments relative to the control, with the L. arvalis and B. subtilis treatments also reducing incidence relative to T. virens. Within the barley/clover rotation, black scurf severity, incidence of black scurf, and the incidence of severe scurf were all lower with T. virens than all other treatments (Table 3).
After tubers were harvested and washed, substantial amounts of common scab were detected on tubers and rated based on the percentage of each tuber surface covered by scab lesions. There were no significant rotation or biocontrol treatment effects or interactions for scab at location 1, which had high incidences (85.8 to 95.6%) but low scab severity (2.0 to 2.5% coverage) across all samples. Scab severity was much higher at location 2 (22.5 to 30.9% coverage), and the barley/clover rotation resulted in greater scab severity (30.9% coverage) than the barley/ryegrass and potato rotations (22 to 26% coverage). However, there were no biocontrol effects or rotation by biocontrol interactions.
3.3. Tuber Yield
Preceding rotation crops significantly affected tuber yield at location 1, but not at location 2. There were no rotation crop by biocontrol interactions at either location. At location 1, the barley/ryegrass rotation resulted in significantly higher total, marketable and large size class yield than the potato rotation (Table 4), with increases averaging 18, 24, and 40% for these three yield groups, respectively. The potato rotation produced more small and medium size class tubers than both the barley/clover and barley/ryegrass rotations. The barley/ryegrass and barley/clover rotations also resulted in a greater percentage of tubers that were marketable (Figure 2A) or in the large size class (Figure 2B) than the potato rotation.
Biocontrol treatments significantly affected some aspects of tuber yield at location 2 but not at location 1 (Table 4). At location 2, L. arvalis and T. virens treatments resulted in a greater percentage of tubers in the marketable (Figure 2C) and larger size classes (Figure 2D) than the control, as well as greater yield of large size tubers, but total yield was not statistically different among treatments (Table 4). L. arvalis and T. virens treatments also resulted in fewer tubers in the small and medium size classes than the control.
3.4. Soil Rhizoctonia Populations
Soil populations of R. solani in the field trials were generally too low to detect any differences (0 to 0.5 propagules per 10 g soil) among treatments. At location 1, R. zeae was also frequently isolated at the Spring sampling times (both prior to and after incorporation of rotation crop residues) but was not associated with a particular rotation crop. R. zeae was not isolated from location 2 soil.
3.5. Analysis of Microbial Communities Using FAME Profiles
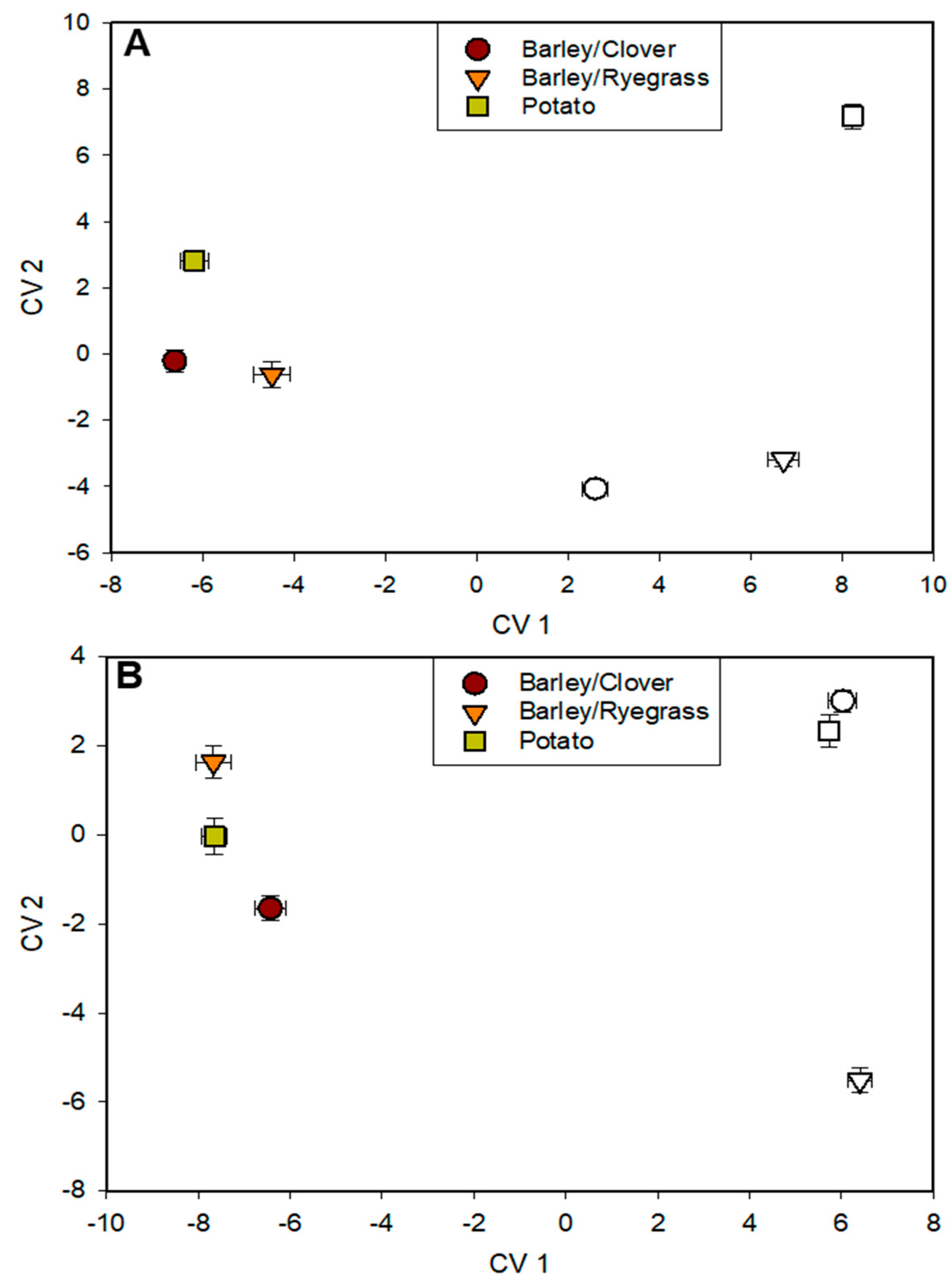
Fatty acid profiles revealed distinct differences in soil microbial communities associated with preceding rotation crops, as well as differences between soils prior to and after incorporation of crop residues at both locations, as indicated by canonical variates analyses. At location 1, graphical depiction of the first two canonical variates (CV1 and CV2) representing the soil FAME profiles from each rotation were very different prior to incorporation, and incorporation resulted in further changes, with each rotation maintaining distinct profiles but also showing greater similarity than before incorporation (Figure 3A). Differences primarily relating to crop incorporation, as well as differences between the barley/ryegrass and barley/clover rotations, were primarily delineated along CV1, whereas differences separating the barley rotations from the potato rotation were primarily delineated along CV2. The fatty acids that contributed most to CV1 at location 1 were 12:0, 13:0 anteiso, 18:3 ω6c, 16:1 ω5c, 17:1 ω3c, and C:16 N alcohol. Those that contributed most to CV2 included 18:2 ω6c, 16:1 ω5c, 16:1 ω7c, 12:0, C16 N alcohol, and 17:1 ω3c. At location 2, profiles associated with the barley/ryegrass rotation were very distinct from the other rotations prior to crop incorporation, and incorporation resulted in reductions in CV1 and consolidation of characteristics among the rotations, although each rotation maintained a distinct profile delineated along CV2 (Figure 3B). CV1 was represented mostly by contributions from 13:0 anteiso, C16 N alcohol, 18:0 2OH, 16:1 ω7c 18:1 ω9c, 17:1 ω3c, 18:3 ω6c, and CV2 by 13:0 anteiso, 16:1 ω5c, 18:2 ω6c, 18:1 ω9c, C16 N alcohol, and 18:0 2OH.
In August, FAME profiles were influenced by both rotation crop and biocontrol treatments at both locations, with biocontrol treatments producing somewhat different effects depending on the rotation crop (Figure 4). For example, at location 1, L. arvalis treatments resulted in greater CV1 and CV2 values than the control within the barley/ryegrass rotation, but, in the potato rotation, lower values for all biocontrol treatments were observed, and lower values for B. subtilis treatments were observed in both rotations (Figure 4A). Variability within CV1 was determined primarily by 12:0, 13:0 anteiso, 16:1 ω5c, 18:1 ω9c, and 18:2 ω6c. The fatty acids that accounted for the most variability in CV2 were 10:0 3 OH, 16:0, 17:1 ω7c, 18:3 ω6c, 16:0, and 18:1 ω9c. At location 2, differences in soil FAME profiles associated with rotation crops were primarily delineated along CV1, whereas differences among biocontrol treatments within rotations were separated along CV2 (Figure 4B). However, the B. subtilis treatments resulted in more similar profiles regardless of rotation, whereas the other biocontrol treatments had different effects on FAME profiles depending on the rotation. Fatty acids contributing most to CV1 were 13:0, 16:1 ω5c, 17:1 ω7c, 18:2 ω6c, and 18:1 ω9c, and most contributing to CV2 were 10:0 3OH, 12:0 3OH, 15:0 iso, 16:1 ω7c, 16:0, 17:1 ω3c, and 18:3 ω6c.
Differences in fatty acid profiles were also expressed as the proportion of extracted fatty acids in specific structural classes, which also showed differences among preceding rotation crops both prior to and after crop residue incorporation at both locations (Table 5). Overall, the proportion of straight chain saturated and branched fatty acids tended to be greater prior to residue incorporation than after incorporation. The fungal to bacterial fatty acid ratio was greater after residue incorporation. The potato rotation resulted in higher levels of straight chain saturated and branched fatty acids and lower levels of polyunsaturated fatty acids and monounsaturated to saturated fatty acid ratios than the barley rotations both prior to and after residue incorporation. Prior to residue incorporation, continuous potato had a lower fungal to bacterial ratio, while after residue incorporation, it had a lower proportion of monounsaturated fatty acids than the barley rotations at both locations.
There were also differences among rotations and biocontrol treatments in the proportion of fatty acids in specific structural classes and ratios of those proportions in the August soil samples. There were significant rotation crop effects and lesser biocontrol effects observed at both locations. There were some consistent effects observed at both locations, such as generally lower ratios of monounsaturated to saturated fatty acids and lower fungi to bacteria ratios in the potato rotation compared to the barley rotations (Table 6). At location 2, both potato and barley/clover rotations resulted in a greater proportion of hydroxyl fatty acids than barley/ryegrass, the potato rotation resulted in a lower ratio of monounsaturated to saturated fatty acids than both barley rotations, and the barley/ryegrass rotation had a higher fungi/bacteria ratio than both barley/clover and potato rotations (Table 6). There were also significant biocontrol effects at location 2. T. virens resulted in a greater proportion of polyunsaturated fatty acids than the control, whereas the control resulted in a greater proportion of branched fatty acids than L. arvalis and T. virens and a greater gram-positive to gram-negative ratio than L. arvalis. L. arvalis also resulted in a greater proportion of hydroxyl group containing fatty acids than B. subtilis. At location 1, there were significant rotation crop by biocontrol interactions for saturated and polyunsaturated fatty acid classes and the ratio of fungal to bacterial fatty acids. There were also significant differences among biocontrol treatments within at least one of the rotation crops for the remaining fatty acid classes and class ratios presented (Table 6). However, differences among biocontrol treatments were confined to individual rotations, with no consistent trends observed over multiple different rotations.
4. Discussion
This research demonstrated the potential benefits of replacing clover as a cover crop in barley rotations with ryegrass, for better control of Rhizoctonia disease and improved potato yield. This research also documented the complex interactions among crop rotations, biocontrol treatments, and soil microbial communities and the potential for improved integrated disease management with effective combinations of crop rotations and biocontrol organisms.
Ryegrass rotations reduced severity of Rhizoctonia stem canker alone in the greenhouse and at one of two field locations when combined with barley. An increase in tuber yield with the barley/ryegrass rotation also occurred at the same location where disease reductions were detected. Stem canker can result in severe losses in yield [41], and the reductions in stem canker by the barley/ryegrass rotation may have led to the increase in yield. Reductions in severity were not detected at one of the two locations. It is hypothesized that this might be because both the ryegrass and clover cover crops were not fully established the following spring due to a hard winter and severe weed pressure at that location that substantially reduced cover crop stands from the previous fall. It has been well-established that cover crop biomass production is integral to effects on soil properties [42,43]. As for whether the “Lemtal” ryegrass variety performed better than common ryegrass, based on the greenhouse tests, both types resulted in comparable suppression of Rhizoctonia disease, with no indication of any significant difference in their ability to reduce disease or their suitability as a cover crop. Previous indications of the efficacy of Lemtal ryegrass were based primarily on anecdotal observations of successful plantings, with no direct comparisons among different ryegrass varieties. Thus, there is no direct evidence that Lemtal ryegrass provides any better disease control than other ryegrass varieties.
This study indicates that ryegrass is a more suitable cover crop than clover for control of Rhizoctonia disease of potato. In the greenhouse, a rotation with clover alone increased R. solani populations as much as or more than potato rotations. These increases in populations and disease levels by clover and other legumes have been observed in other studies [6,15,44]. R. solani AG-3 can produce stem lesions on clover in controlled studies, and leguminous crops, including clover, have shown increases in lesions caused by R. solani on subsequent susceptible crops [14,15]. However, one previous study observed lower soilborne disease levels for black scurf and common scab with a red clover rotation relative to barley and ryegrass, even though red clover resulted in higher plant-parasitic nematode populations and lower tuber yield than ryegrass [45]. That study involved somewhat different soil and environmental conditions than the present study. The beneficial effects observed with the barley/ryegrass rotation were not observed with barley/clover in the present research. The barley/ryegrass combination can potentially provide better disease control, as was also demonstrated in a previous direct comparison of these combinations [14].
A possible mechanism of disease suppression by ryegrass is that it stimulates microbial activity [42]. In addition, the ryegrass rotation could stimulate specific antagonists that reduce disease. In the present study, ryegrass rotations increased populations of R. zeae in the greenhouse. Although R. zeae has mostly been considered a pathogen of turfgrasses [46,47], isolates recovered in Maine did not cause disease symptoms on either ryegrass or barley (Brewer, unpublished). R. zeae can cause some lesions on potato, but to a much lesser extent than R. solani AG-3 [48]. R. zeae has shown potential biocontrol activity against R. solani and may compete with R. solani for colonization or infection of roots [49]. In previous greenhouse biocontrol trials, an isolate of R. zeae from this field location was among the most effective biocontrol isolates tested for reducing both stem canker and black scurf [22]. Additional tests need to be conducted to determine the role, if any, of R. zeae and suppression of Rhizoctonia disease by ryegrass rotations.
The barley/ryegrass rotations did not reduce black scurf in the present study. Differences in black scurf among rotations may not have been observed in the present study because severity levels were relatively low. In addition, disease measurements were taken after only one complete rotation cycle, and it may take more rotation cycles before differences can be detected. Cover crop effects on soil properties often take several seasons to fully develop, with longer-term studies generally showing greater effects than short-term trials [45,50]. In a separate field study, in Maine, where a barley/ryegrass rotation was directly compared to a barley/clover rotation, a significant reduction in black scurf, as well as common scab, was observed with the barley/ryegrass rotation over multiple field seasons [14]. More recent ongoing trials have also indicated consistent reductions in black scurf with barley/ryegrass rotations relative to some other rotations [51].
FAME profiles, as represented by canonical variates analyses and the proportions of fatty acid structural classes and biomarkers, revealed distinct differences for all rotations, indicating that specific changes in soil microbial community characteristics occurred based on the preceding crop. This supports the growing body of literature demonstrating the considerable influence of plants and plant residues on shaping the soil microbiome [8,9,10,52]. Many fatty acids and ratios of fatty acids have been identified as particularly useful biomarkers [30,53]. The proportion of monounsaturated fatty acids was higher in the barley/ryegrass rotation than the other rotations, particularly after residue incorporation, and the ratios of monounsaturated to saturated fatty acids were higher in both barley rotations than continuous potato. Monounsaturated fatty acids are a biomarker for gram negative bacteria, as well as some fungi (16:1 ω5c and 18:1 ω9c), and these fatty acids tend to increase in soil with aerobic conditions, organic inputs, and high substrate availability [6,53,54]. The ratio of monounsaturated to saturated fatty acids has been used as an indicator to evaluate communities and environmental conditions, with observed ratios of greater than 1 for cultivated soils with good C content and organic inputs, and values less than 1 representing low substrate, low organic input soils [53,55]. Our results showing lower values for the potato rotation than the barley rotations are consistent with other studies, which have also shown even greater differences (with continuous potato values decreasing, barley and other rotation values increasing over time) the longer the rotations are in place [6,11,14].
The proportions of polyunsaturated fatty acids, which are associated with fungi, were also higher in the barley rotations than the potato rotation, and also tended to increase after crop residue incorporation. Hydroxyl group fatty acids are indicators of gram-negative bacteria, whereas branched fatty acids are indicators of gram-positive bacteria [27,32]. Results of this study indicate that the proportion of gram-positive bacteria was higher in continuous potato than the barley rotations, and the ratio of fungi to bacteria was higher in the barley rotations, especially barley/ryegrass, than the continuous potato rotation. These results are similar to those observed between grass-grain rotations and continuous potato treatments in other crop rotation studies in Maine [6,12,14,27].
FAME profiles also revealed differences in soil microbial communities associated with different biocontrol treatments at both locations. However, the biocontrol treatments affected FAME profiles differently within each rotation, resulting in interactions between the rotations and biocontrol treatments on their effects. Interestingly, although, for the most part, rotations appeared to exert a greater effect on the soil microbial communities than the biocontrol treatments, at location 2, the B. subtilis treatment resulted in similar microbial characteristics regardless of the rotation, indicating that the addition of B. subtilis had a greater effect on defining the microbial characteristics than rotations for that soil. Proportions of fatty acid structural classes, class ratios, and biomarkers were also affected by biocontrol treatments. At location 1, fatty acid classes and ratios associated with the biocontrol treatments varied considerably among rotations, demonstrating the complex effects among rotations and biocontrol treatments on soil microbial communities. Previous research has documented that biocontrol organisms can be greatly affected by the indigenous microbial communities present but can also have significant effects on those microbial communities [24,34,56]. In one study [28], a mixture of various biocontrol organisms was more effective within a barley/ryegrass rotation than either a barley/clover or potato rotation, indicating the importance of a rotation that adequately supports the biocontrol activities of the added microbial agent. Both B. subtilis and T. virens has been previously shown to engage multiple mechanisms of action for disease control, including the induction of host defense responses, competition, and antibiosis [57,58], whereas L. arvalis is believed to function primarily as a mycoparasite in reducing pathogen populations [20].
Many management practices for sustainable disease control affect soil microbiology characteristics. Changes in microbial activity or community structure can be mechanisms by which effective rotation crops and biocontrol reduce disease [5,6,11]. Although community level approaches, such as FAME analyses, do not specifically identify the microorganisms that are responsible for the changes observed, these approaches are very sensitive to detecting changes in soil microbial characteristics and do provide useful information regarding the types of changes and groups of organisms involved. Larkin and Honeycutt [6] demonstrated that many associations exist between microbial populations, FAME analysis, tuber yield, and Rhizoctonia disease of potato. In that study, the incidence and severity of black scurf in potato cropping systems were negatively correlated with FAME principal components and the ratio of monounsaturated to saturated fatty acids. Total and marketable yield were also negatively correlated with scurf incidence and positively correlated with fungal populations. In the present study, the barley/ryegrass rotation resulted in a greater proportion of monounsaturated fatty acids, a greater ratio of monounsaturated to saturated fatty acids, and an increase in the ratio of fungi to bacteria over the potato rotation. The barley/ryegrass rotation also increased yield and reduced stem canker, which may be influenced by specific changes in soil microbial communities.
Numerous interactions between crop rotations and biocontrol organisms were detected. Biocontrol treatments did provide some control of black scurf within the barley rotations at one location, although control by specific treatments varied within the different rotations. L. arvalis and B. subtilis reduced black scurf within the barley/ryegrass rotation, whereas T. virens reduced black scurf severity within the barley/clover rotation. L. arvalis and T. virens treatments increased the percentage of marketable and large size class tubers. Biocontrol by rotation interactions were evident in black scurf results, microbial populations, FAME profiles, and fatty acid structural classes. L. arvalis appeared to be more effective in the barley/ryegrass rotation, whereas T. virens was more effective in the barley/clover rotation at location 1.
Overall, both effective crop rotations and the addition of biological antagonists are potentially sustainable disease management practices that may reduce losses in tuber yield and quality from R. solani and other soilborne diseases of potato. The use of biocontrol within beneficial rotations may substantially improve efficacy against R. solani. This integrated approach demonstrates that microbial antagonists and effective crop rotations can potentially increase yield and suppress Rhizoctonia disease of potato, but the complex interactions involving crop rotations, biological amendments, and soil microbial communities and their roles in disease suppression needs to be better understood before the full potential of this approach can be achieved.
Author Contributions
R.P.L. conceived and planned the research, analyzed, and interpreted the data, and wrote, edited, and revised the manuscript. M.T.B. planned and set-up the experiments, collected, and analyzed the data, and wrote the initial draft of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We would like to thank Beth Case and Lindsay Colley for their technical assistance and Dave Torrey and Dwight Cowperthwaite for maintenance of the field plots.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tsror, L. Biology, ecology and management of Rhizoctonia solani on potato. J. Phytopathol. 2010, 158, 649–658. [Google Scholar] [CrossRef]
- Powelson, M.L.; Rowe, R.C. Management of diseases caused by seedborne and soilborne fungi and fungus-like pathogens. In Potato Health Management, 2nd ed.; Johnson, D.A., Ed.; APS Press: St. Paul, MN, USA, 2008; pp. 149–158. [Google Scholar]
- Secor, G.A.; Gudmestad, N.C. Managing fungal diseases of potato. Can. J. Plant Pathol. 1999, 21, 213–221. [Google Scholar] [CrossRef]
- Stevenson, W.R.; Loria, R.; Franc, G.D.; Weingartner, D.P. (Eds.) Compendium of Potato Diseases, 2nd ed.; APS Press: St. Paul, MN, USA, 2001; 106p. [Google Scholar]
- Huber, D.M.; Sumner, D.R. Suppressive soil amendments for the control of Rhizoctonia species. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology, and Disease Control; Sneh, B., Jabaji-Hare, S., Neate, S., Dijst, G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 433–443. [Google Scholar]
- Larkin, R.P.; Honeycutt, C.W. Effects of different 3-year cropping systems on soil microbial communities and soilborne disease of potato. Phytopathology 2006, 96, 68–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, R.P. Soil health paradigms and implications for disease management. Ann. Rev. Phytopathol. 2015, 53, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Ann. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef]
- O’Donnell, A.G.; Seasman, M.; Macrae, A.; Waite, I.; Davies, J.T. Plants and fertilizers as drivers of change in microbial community structure. Plant Soil 2001, 232, 135–145. [Google Scholar] [CrossRef]
- Welbaum, G.E.; Sturz, A.V.; Dong, Z.; Nowak, J. Managing soil microorganisms to improve productivity of agro-ecosystems. Crit. Rev. Plant Sci. 2004, 23, 175–193. [Google Scholar] [CrossRef]
- Larkin, R.P.; Honeycutt, C.W.; Griffin, T.S.; Olanya, O.M.; Halloran, J.M.; He, Z. Effects of different potato cropping system approaches and water management on soilborne diseases and soil microbial communities. Phytopathology 2011, 101, 55–67. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S.; Honeycutt, C.W.; Olanya, O.M.; He, Z.; Halloran, J.M. Cumulative and residual effects of different potato cropping system management strategies on soilborne diseases and soil microbial communities over time. Plant Pathol. 2017, 66, 437–449. [Google Scholar] [CrossRef]
- Halloran, J.M.; Griffin, T.S.; Honeycutt, C.W. Economic analysis of potential rotation crops for Maine potato cropping systems. Am. J. Potato Res. 2005, 82, 155–162. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, G.S.; Honeycutt, C.W. Rotation and cover crop effects on soilborne potato diseases, tuber yield, and soil microbial communities. Plant Dis. 2010, 94, 1491–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabney, S.M.; Schreiber, J.D.; Rothrock, C.S.; Johnson, J.R. Cover crops affect sorghum seedling growth. Agron. J. 1996, 88, 961–970. [Google Scholar] [CrossRef]
- Asaka, O.; Shoda, M. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilus RB14. Appl. Environ. Microbiol. 1996, 62, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.R.; Adkins, P.G. Versatility of fungal and bacterial isolates for biological control of damping-off disease caused by Rhizoctonia solani and Pythium spp. Biol. Control 1999, 15, 10–18. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, N.; Yong, X.; Yang, X.; Shen, Q. Biocontrol of Rhizoctonia solani damping-off disease in cucumber with Bacillus pumilus SQR-N43. Microbiol. Res. 2012, 167, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Hijri, M. Screening, identification and evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS Microbiol. Lett. 2010, 311, 152–159. [Google Scholar] [CrossRef]
- Murdoch, C.W.; Leach, S.S. Evaluation of Laetisaria arvalis as a biological control agent of Rhizoctonia solani on white potato. Am. Potato J. 1993, 70, 625–633. [Google Scholar] [CrossRef]
- Tsror, L.; Barak, R.; Sneh, B. Biological control of black scurf on potato under organic management. Crop Prot. 2001, 20, 145–150. [Google Scholar] [CrossRef]
- Brewer, M.T.; Larkin, R.P. Efficacy of several potential biocontrol organisms against Rhizoctonia solani on potato. Crop Prot. 2005, 24, 939–950. [Google Scholar] [CrossRef]
- Larkin, R.P.; Tavantzis, S. Use of biocontrol organisms and compost amendments for improved control of soilborne diseases and increased potato production. Am. J. Pot. Res. 2013, 90, 261–270. [Google Scholar] [CrossRef]
- Larkin, R.P. Impacts of biocontrol products on Rhizoctonia disease of potato and soil microbial communities, and their persistence in soil. Crop Prot. 2016, 90, 96–105. [Google Scholar] [CrossRef]
- Bernard, E.; Larkin, R.P.; Tavantzis, S.; Erich, M.S.; Alyokhin, A.; Gross, S. Rapeseed rotation, compost and biocontrol amendments reduce soilborne diseases and increase tuber yield in organic and conventional potato production systems. Plant Soil 2014, 374, 611–627. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; Grünwald, N.J.; Bolda, M. Cultural methods and soil nutrient status in low and high input agricultural systems, as they affect Rhizoctonia species. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology, and Disease Control; Sneh, B., Jabaji-Hare, S., Neate, S., Dijst, G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 407–421. [Google Scholar]
- Larkin, R.P. Characterization of soil microbial communities under different potato cropping systems by microbial population dynamics, substrate utilization, and fatty acid profiles. Soil Biol. Biochem. 2003, 35, 1451–1466. [Google Scholar] [CrossRef]
- Larkin, R.P. Relative effects of biological amendments and crop rotations on soil microbial communities and soilborne diseases of potato. Soil Biol. Biochem. 2008, 40, 1341–1351. [Google Scholar] [CrossRef]
- Han, J.S.; Cheung, J.H.; Yoon, T.M.; Song, J.; Rajkarnakar, A.; Kim, W.G.; Yoo, I.D.; Yang, Y.Y.; Suh, J.W. Biological control agent of common scab disease by antagonistic strain Bacillus sp. sunhua. J. Appl. Microbiol. 2005, 99, 213–221. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Zhuang, L.; Yu, Y.; Liu, J.; Zhang, L.; Gao, Z.; Wu, Y.; Gao, W.; Ding, G.; et al. A rhizosphere-derived consortium of Bacillus subtilis and Trichoderma harzianum suppresses common scab of potato and increases yield. Comput. Struct. Biotechnol. J. 2019, 17, 645–653. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schliefer, K.H. Phylogenteic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar]
- Cavigelli, M.A.; Robertson, G.P.; Klug, M.J. Fatty acid methyl ester (FAME) profiles as measures of soil microbial community structure. Plant Soil 1995, 170, 99–113. [Google Scholar] [CrossRef]
- Ritchie, N.J.; Schutter, M.E.; Dick, R.P.; Myrold, D.D. Use and length of heterogeneity PCR and fatty acid methyl ester profiles to characterize microbial communities in soil. Appl. Environ. Microbiol. 2000, 66, 1668–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siciliano, S.D.; Germida, J.J. Biolog analysis and fatty acid methyl ester profiles indicate that Pseudomonad inoculants that promote phytorediation alter the root associated microbial community of Bromus biebersteinii. Soil Biol. Biochem. 1998, 30, 1717–1723. [Google Scholar] [CrossRef]
- Essah, S.Y.C.; Honeycutt, C.W. Tillage and seed-sprouting strategies to improve potato yield and quality in short season climates. Am. J. Potato Res. 2004, 81, 177–186. [Google Scholar] [CrossRef]
- Flowers, R.A. A selective media for isolation of Rhizoctonia from soil and plant tissue. Proc. Am. Phytopathol. Soc. 1976, 3, 219. [Google Scholar]
- Henis, Y.; Ghaffar, A.; Baker, R.; Gillespie, S.L. A new soil-sampler and its use for the study of population dynamics of Rhizoctonia solani in soil. Phytopathology 1978, 68, 371–376. [Google Scholar] [CrossRef]
- Sneh, B.; Katan, J.; Henis, Y.; Wahl, I. Methods for evaluating inoculum density of Rhizoctonia in naturally infested soil. Phytopathology 1966, 56, 74–78. [Google Scholar]
- Buyer, J.S.; Roberts, D.P.; Russek-Cohen, E. Microbial community structure and function in the spermosphere as affected by soil and seed type. Can. J. Microbiol. 1999, 45, 138–144. [Google Scholar] [CrossRef]
- Sneh, B.; Burpee, L.; Ogoshi, A. Identification of Rhizoctonia Species; APS Publishing: St. Paul, MN, USA, 1991. [Google Scholar]
- Read, P.J.; Hide, G.A.; Firmager, J.P.; Hall, S.M. Growth and yield of potatoes as affected by severity of stem canker (Rhizocrtonia solani). Potato Res. 1989, 32, 9–15. [Google Scholar] [CrossRef]
- Hubbard, R.K.; Strickland, T.C.; Phatak, S. Effects of cover crop systems on soil physical properties and carbon/nitrogen relationships in the coastal plain of southeastern USA. Soil Till. Res. 2013, 126, 276–283. [Google Scholar] [CrossRef]
- Schutter, M.E.; Dick, R.P. Microbial community profiles and activities among aggregates of winter fallow and cover-cropped soil. Soil Sci. Soc. Am. J. 2002, 66, 142–153. [Google Scholar] [CrossRef]
- Johnston, H.W.; Celetti, M.J.; Kimpinski, J.; Platt, H.W. Fungal pathogens and Pratylenchus penetrans associated with preceding crops of clover, winter wheat, and annual ryegrass and their influence on succeeding potato crops on Prince Edward Island. Am. Potato J. 1994, 71, 797–808. [Google Scholar] [CrossRef]
- Carter, M.R.; Kunelius, H.T.; Sanderson, J.B.; Kimpinski, J.; Platt, H.W.; Bolinder, M.A. Productivity parameters and soil health dynamics under long-term 2-year potato rotations in Atlantic Canada. Soil Till. Res. 2003, 72, 152–168. [Google Scholar] [CrossRef]
- Burpee, L.; Martin, B. Biology of Rhizoctonia species associated with turfgrasses. Plant Dis. 1992, 76, 112–117. [Google Scholar] [CrossRef]
- Martin, S.B.; Lucas, L.T. Pathogenicity of Rhizoctonia zeae on tall fescue and other turfgrasses. Plant Dis. 1983, 67, 676–678. [Google Scholar] [CrossRef]
- Carling, D.E.; Leiner, R.H. Effect of temperature on virulence of Rhizoctonia solani and other Rhizoctonia on potato. Phytopathology 1990, 80, 930–934. [Google Scholar] [CrossRef]
- Webb, K.M.; Harveson, R.M.; West, M.S. Evaluation of Rhizoctonia zeae as a potential biological control option for fungal root diseases of sugar beet. Ann. Appl. Biol. 2015, 167, 75–89. [Google Scholar] [CrossRef]
- Carter, M.R.; Sanderson, J.B. Influence of conservation tillage and rotation length on potato productivity, tuber disease and soil quality parameters on a fine sandy loam in eastern Canada. Soil Till. Res. 2001, 63, 1–13. [Google Scholar] [CrossRef]
- Larkin, R.P. Incorporating disease-suppressive rotation crops and organic amendments into improved potato cropping systems. Phytopathology 2017, 107. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142. [Google Scholar] [CrossRef]
- Zelles, L.; Bai, Q.Y.; Beck, T.; Beese, F. Signature fatty acids in phospholipids and lipopolysaccharides as indicators of microbial biomass and community structure in agricultural soils. Soil Biol. Biochem. 1992, 24, 317–323. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef]
- Zelles, L.; Bai, Q.Y.; Rackwitz, R.; Chadwick, D.; Beese, F. Determination of phospholipid- and lipopolysaccaride-derived fatty acids as an estimate of microbial biomass and community structures in soil. Biol. Fertil. Soils 1995, 19, 115–123. [Google Scholar] [CrossRef]
- Gilbert, G.S.; Parke, J.L.; Clayton, M.K.; Handelsman, J. Effects of an introduced bacterium on bacterial communities on roots. Ecology 1993, 74, 840–854. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. with special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef] [PubMed]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Soil populations of R. solani prior to and after incorporation of rotation crop residues in repeated greenhouse trials for (A) experiment 1 and (B) experiment 2 and soil populations of R. zeae in (C) experiment 1 after incorporation of residues and (D) experiment 2 both prior to and after incorporation of residues. Inoculum of R. solani Anastomosis Group (AG)-3 was added at the start of each experiment.
Figure 1.
Soil populations of R. solani prior to and after incorporation of rotation crop residues in repeated greenhouse trials for (A) experiment 1 and (B) experiment 2 and soil populations of R. zeae in (C) experiment 1 after incorporation of residues and (D) experiment 2 both prior to and after incorporation of residues. Inoculum of R. solani Anastomosis Group (AG)-3 was added at the start of each experiment.

Figure 2.
Percent yield for (A) marketable tubers and (B) large size class tubers by preceding rotation crop at location 1 (Central Maine) and for (C) marketable tubers and (D) large size class tubers by biocontrol treatment at location 2 (Northern Maine).
Figure 2.
Percent yield for (A) marketable tubers and (B) large size class tubers by preceding rotation crop at location 1 (Central Maine) and for (C) marketable tubers and (D) large size class tubers by biocontrol treatment at location 2 (Northern Maine).

Figure 3.
Effects of rotation crops on soil fatty acid methyl ester (FAME) profiles as represented by canonical variates (CV) 1 and 2 from canonical variates analysis from soils sampled prior to (open symbols) and one month after incorporation (filled symbols) of rotation crop residues in the Spring of year 2 at (A) location 1 (Central Maine) and (B) location 2 (Northern Maine).
Figure 3.
Effects of rotation crops on soil fatty acid methyl ester (FAME) profiles as represented by canonical variates (CV) 1 and 2 from canonical variates analysis from soils sampled prior to (open symbols) and one month after incorporation (filled symbols) of rotation crop residues in the Spring of year 2 at (A) location 1 (Central Maine) and (B) location 2 (Northern Maine).

Figure 4.
Effects of preceding rotation crop and biocontrol treatments on soil FAME profiles as represented by canonical variates (CV) 1 and 2 from canonical variates analysis from soils sampled in August of year 2 at (A) location 1 (Central Maine) and (B) location 2 (Northen Maine). Error bars represent the standard error of the mean for each CV. Biocontrol treatments represented by: Bs = B. subtilis; La = L. arvalis; Tv = T.virens; Con = nontreated control.
Figure 4.
Effects of preceding rotation crop and biocontrol treatments on soil FAME profiles as represented by canonical variates (CV) 1 and 2 from canonical variates analysis from soils sampled in August of year 2 at (A) location 1 (Central Maine) and (B) location 2 (Northen Maine). Error bars represent the standard error of the mean for each CV. Biocontrol treatments represented by: Bs = B. subtilis; La = L. arvalis; Tv = T.virens; Con = nontreated control.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of preceding rotation crops on stem canker and size of potato shoots in greenhouse experiments.
Table 1.
Effects of preceding rotation crops on stem canker and size of potato shoots in greenhouse experiments.
| Rotation Crop | Discolored Stem Incidence w (%) | Stem Canker Incidence x (%) | Stem Canker Severity (0–5) w | Shoot Size y |
|---|---|---|---|---|
| Experiment 1 | ||||
| Barley | 68.8 ab z | 0.0 c | 0.69 ab | 2.31 abc |
| Barley/clover | 62.5 b | 6.3 ab | 0.69 ab | 2.56 a |
| Barley/ryegrass | 87.5 ab | 12.5 bc | 1.00 ab | 2.37 ab |
| Clover | 68.8 ab | 37.5 a | 1.31 a | 1.93 c |
| Common ryegrass | 56.3 b | 12.5 bc | 1.06 ab | 2.44 a |
| Lemtal ryegrass | 87.5 ab | 18.8 ab | 0.69 b | 2.44 a |
| Potato | 93.8 a | 25.0 ab | 1.25 a | 2.00 bc |
| Experiment 2 | ||||
| Barley | 93.8 ab | 39.5 ab | 1.50 ab | 2.33 ab |
| Barley/clover | 87.5 ab | 31.3 ab | 1.52 ab | 2.62 a |
| Barley/ryegrass | 93.8 ab | 18.8 b | 1.12 bc | 2.73 a |
| Clover | 91.8 ab | 43.8 a | 1.79 a | 2.15 b |
| Common ryegrass | 83.3 ab | 14.5 b | 1.00 c | 2.38 ab |
| Lemtal ryegrass | 68.8 b | 33.3 ab | 1.08 bc | 2.69 a |
| Potato | 100.0 a | 37.5 ab | 1.73 a | 2.71 a |
w Discolored stems included those with a severity rating of 1 or greater. Disease severity ratings for stem canker were on a scale of 0 to 5 (0 = no symptoms; 1 = brown discoloration; 2 = cankers covering <25% of the stem circumference; 3 = 25% to 75% coverage by cankers; 4 = 75% coverage by stem cankers; and 5 = stem completely nipped off or death of the plant). x Incidence of stem canker was represented by the percentage of plants showing obvious canker symptoms (severity of 2 or greater). y Shoot size ratings were on a scale of 0 to 3 as follows: 0 = no growth; 1 = shoot height <2 cm; 2 = shoot height of 2 to 5 cm; and 3 = shoot height >5 cm. z Means within the same column for each experiment followed by the same letter are not significantly different according to Fisher’s Least Significant Difference (LSD) (p = 0.05).
Table 2.
Rhizoctonia disease ratings by preceding rotation crop or biocontrol treatment at location 1 (Central Maine).
Table 2.
Rhizoctonia disease ratings by preceding rotation crop or biocontrol treatment at location 1 (Central Maine).
| Shoot Emergence w (%) | Stem Canker | ||
|---|---|---|---|
| Severity x | Incidence y (%) | ||
| Preceding Rotation Crop (Mainplots) | |||
| Barley/ryegrass | 75.4 a z | 1.33 b | 25.0 b |
| Barley/clover | 79.3 a | 1.45 ab | 39.1 b |
| Potato | 84.2 a | 1.70 a | 59.4 a |
| Biocontrol Treatment (Subplots) | |||
| Laetisaria arvalis | 83.9 a | 1.53 a | 45.8 a |
| Trichoderma virens | 76.7 b | 1.56 a | 45.8 a |
| Bacillus subtilis | 77.6 ab | 1.46 a | 37.5 a |
| Control | 80.4 ab | 1.52 a | 35.4 a |
w Emergence values were taken approximately four weeks after planting. x Disease severity ratings for stem and stolon canker were on a scale of 0 to 5 (0 = no symptoms; 1 = brown discoloration; 2 = cankers covering <25% of the stem or stolon circumference; 3 = 25% to 75% coverage by cankers; 4 = 75% coverage by stem or stolon cankers; and 5 = stem or stolons completely nipped off or death of the plant). y Incidence of stem and stolon canker was represented by the percentage of plants showing obvious canker symptoms (severity of 2 or greater). z Means within the same column for each location and biocontrol treatment or preceding rotation crop followed by the same letter are not significantly different according to Fisher’s LSD (p = 0.05).
Table 3.
Black scurf ratings for biocontrol treatments as affected by preceding rotation crop at location 1 (Central Maine).
Table 3.
Black scurf ratings for biocontrol treatments as affected by preceding rotation crop at location 1 (Central Maine).
| Rotation Crop | Severity x | Incidence (%) | Incidence ≥ 2 y (%) |
|---|---|---|---|
| Biocontrol Treatment | |||
| Barley/Ryegrass | |||
| Laetisaria arvalis | 1.68 c z | 64.9 b | 43.3 c |
| Trichoderma virens | 2.21 b | 87.3 a | 55.8 b |
| Bacillus subtilis | 1.96 bc | 84.3 a | 43.4 c |
| Control | 2.69 a | 85.8 a | 71.8 a |
| Barley/Clover | |||
| Laetisaria arvalis | 1.76 a | 74.1 a | 49.9 a |
| Trichoderma virens | 1.02 b | 52.4 b | 25.8 b |
| Bacillus subtilis | 1.83 a | 70.8 a | 45.8 a |
| Control | 1.74 a | 83.4 a | 49.1 a |
| Potato | |||
| Laetisaria arvalis | 1.81 a | 75.8 a | 60.1 a |
| Trichoderma virens | 1.77 a | 79.0 a | 56.6 a |
| Bacillus subtilis | 1.98 a | 75.1 a | 57.5 a |
| Control | 1.80 a | 78.3 a | 60.0 a |
x Black scurf severity was based on the percent of the tuber surface covered by sclerotia. y Incidence of tubers with severity rating of 2% coverage or greater. z Means within the same column for each preceding rotation crop followed by the same letter are not significantly different according to Fisher’s LSD (p = 0.05).
Table 4.
Tuber yield by size class for preceding rotation crop or biocontrol treatment.
| Yield (Mg/ha) | ||||||
|---|---|---|---|---|---|---|
| Small v | Medium w | Large x | Total | Marketable y | Misshapen | |
| Location 1 (Central Maine) | ||||||
| Preceding Rotation Crop (Mainplots) | ||||||
| Barley/ryegrass | 0.87 b z | 2.30 b | 14.1 a | 17.3 a | 16.4 a | 1.31 a |
| Barley/clover | 0.92 b | 2.03 b | 12.4 ab | 15.4 ab | 14.5 ab | 1.58 a |
| Potato | 1.54 a | 3.05 a | 10.1 b | 14.7 b | 13.2 b | 1.62 a |
| Biocontrol Treatment (Subplots) | ||||||
| Laetisaria arvalis | 1.05 a | 2.64 a | 12.2 a | 15.9 a | 14.9 a | 1.45 a |
| Trichoderma virens | 1.19 a | 2.17 a | 12.1 a | 15.4 a | 14.3 a | 1.31 a |
| Bacillus subtilis | 1.01 a | 2.60 a | 12.4 a | 16.0 a | 15.0 a | 1.52 a |
| Control | 1.19 a | 2.44 a | 12.2 a | 15.8 a | 14.6 a | 1.74 a |
| Location 2 (Northern Maine) | ||||||
| Preceding Rotation Crop (Mainplots) | ||||||
| Barley/ryegrass | 1.72 a | 4.32 a | 10.6 a | 16.68 a | 15.0 a | 0.75 a |
| Barley/clover | 1.87 a | 4.27 a | 12.4 a | 18.51 a | 16.6 a | 0.80 a |
| Potato | 1.74 a | 5.50 a | 12.3 a | 19.55 a | 17.8 a | 0.78 a |
| Biocontrol Treatment (Subplots) | ||||||
| Laetisaria arvalis | 1.39 b | 4.09 b | 12.9 a | 18.4 a | 17.0 a | 0.66 a |
| Trichoderma virens | 1.46 b | 3.90 b | 12.3 ab | 17.7 a | 16.2 a | 0.87 a |
| Bacillus subtilis | 1.92 ab | 4.98 ab | 12.3 ab | 19.2 a | 17.3 a | 0.90 a |
| Control | 2.34 a | 5.82 a | 9.5 b | 17.6 a | 15.3 a | 0.67 a |
v Small size class consisted of tubers less than 47 mm in diameter. w Medium size class consisted of tubers 47 to 56 mm in diameter. x Large size class consisted of tubers greater than 56 mm in diameter. y Marketable class consisted of tubers in the medium and large size classes combined. z Means within the same column for each location and biocontrol treatment or preceding rotation crop followed by the same letter are not significantly different according to Fisher’s LSD (p = 0.05).
Table 5.
Effect of rotation crops both prior to and after residue incorporation in Spring year 2 on different fatty acid structural classes and class ratios.
Table 5.
Effect of rotation crops both prior to and after residue incorporation in Spring year 2 on different fatty acid structural classes and class ratios.
| Fatty Acid Classes (%) x | Class Ratios y | |||||||
|---|---|---|---|---|---|---|---|---|
| Sat | Mono | Poly | Branch | Hydrox | Mono/Sat | Gram+/Gram− | Fungi/Bact | |
| Location 1 (Central Maine) | ||||||||
| After Growth | ||||||||
| Barley/ryegrass | 32.5 b z | 43.0 a | 10.6 a | 20.8 b | 11.0 b | 1.33 a | 1.94 a | 0.34 a |
| Barley/clover | 32.9 b | 41.4 a | 11.0 a | 20.0 b | 11.9 ab | 1.27 a | 1.71 a | 0.35 a |
| Potato | 38.0 a | 42.1 a | 8.5 b | 23.2 a | 13.3 a | 1.11 b | 1.76 a | 0.24 b |
| After Incorporation | ||||||||
| Barley/ryegrass | 30.7 ab | 41.9 a | 12.4 a | 18.5 ab | 9.4 a | 1.37 a | 1.99 a | 0.45 a |
| Barley/clover | 30.1 b | 39.2 b | 11.9 a | 18.2 b | 10.6 a | 1.31 a | 1.78 a | 0.42 a |
| Potato | 32.3 a | 38.5 b | 12.0 a | 19.6 a | 10.0 a | 1.20 b | 2.02 a | 0.41 a |
| Location 2 (Northern Maine) | ||||||||
| After Growth | ||||||||
| Barley/ryegrass | 20.0 a | 22.5 a | 6.5 ab | 8.8 b | 12.5 a | 1.14 a | 0.72 b | 0.31 a |
| Barley/clover | 20.0 a | 22.3 a | 7.3 a | 9.4 b | 12.3 a | 1.12 a | 0.84 b | 0.35 a |
| Potato | 20.5 a | 22.5 a | 5.1 b | 10.4 a | 10.6 a | 1.10 a | 1.04 a | 0.25 a |
| After Incorporation | ||||||||
| Barley/ryegrass | 18.6 a | 24.7 a | 7.5 a | 8.9 a | 12.8 a | 1.36 a | 0.83 a | 0.37 a |
| Barley/clover | 19.8 a | 23.1 ab | 7.1 a | 9.1 a | 11.1 a | 1.18 ab | 0.89 a | 0.36 ab |
| Potato | 19.8 a | 21.3 b | 6.0 b | 9.2 a | 12.3 a | 1.09 b | 0.81 a | 0.28 b |
x These classes are composed of the proportion of fatty acids belonging to each structural class, where Sat = straight chain fatty acids (9:0, 10:0, 12:0, 13:0, 14:0, 15:0, 16:0), Mono = monounsaturated fatty acids (14:1 ω9c, 15:1 ω8c, 16:1 ω7c, 16:1 ω5c, 17:1 ω7c, 17:1 ω3c, 18:1 ω9c, 20:1 ω6c), Poly = polyunsaturated fatty acids (18:3 ω6c, 18:2 ω6c), Branch = saturated branched fatty acids (11:0 iso, 13:0 iso, 13:0 anteiso, 14:0 iso, 14:0 anteiso, 15:0 iso, 15:0 anteiso), Hydrox = fatty acids with hydroxyl groups (10:0 3OH, 12:0 2OH, 12:0 3OH, 15:0 iso 3OH, 16:0 2OH, 16:0 3OH, 18:0 2OH). y Ratios are of structural class proportions that are used as biomarkers for specific groups of organisms. Mono/sat = the ratio of monounsaturated fatty acids to straight chain saturated fatty acids, Gram+/gram− = the ratio of branched to hydroxy fatty acids, and Fungi/bact = the ratio of polyunsaturated fatty acids to branched and hydroxyl fatty acids. z Means within the same column for each location and sampling time followed by the same letter are not significantly different according to Fisher’s LSD (p = 0.05).
Table 6.
Effects of biocontrol treatments as affected by preceding rotation crop at location 1 and effects of preceding rotation crops and biocontrol treatments at location 2 in August year 2 on different fatty acid structural classes and class ratios.
Table 6.
Effects of biocontrol treatments as affected by preceding rotation crop at location 1 and effects of preceding rotation crops and biocontrol treatments at location 2 in August year 2 on different fatty acid structural classes and class ratios.
| Treatment | Fatty Acid Classes (%) x | Class Ratios y | ||||||
|---|---|---|---|---|---|---|---|---|
| Sat | Mono | Poly | Branch | Hydrox | Mono/Sat | Gram+/Gram− | Fungi/Bact | |
| Location 1 (Central Maine) | ||||||||
| Barley/Ryegrass | ||||||||
| L. arvalis | 26.3 a z | 26.9 a | 7.6 a | 14.5 a | 6.3 a | 1.04 a | 2.39 a | 0.37 ab |
| T. virens | 24.6 a | 26.8 a | 8.8 a | 13.1 b | 5.9 a | 1.11 a | 2.38 a | 0.47 a |
| B. subtilis | 26.7 a | 26.2 a | 6.9 a | 13.0 b | 6.3 a | 1.00 a | 2.24 a | 0.36 ab |
| control | 26.8 a | 26.7 a | 7.2 a | 13.6 ab | 7.3 a | 1.01 a | 2.02 a | 0.35 b |
| Barley/Clover | ||||||||
| L arvalis | 26.3 ab | 26.6 ab | 7.7 a | 14.0 a | 6.7 b | 1.02 b | 2.14 a | 0.38 a |
| T. virens | 24.5 b | 27.8 a | 7.8 a | 14.1 a | 6.2 b | 1.13 a | 2.32 a | 0.39 a |
| B. subtilis | 27.4 a | 25.9 b | 6.4 b | 13.8 a | 7.8 a | 0.98 b | 1.84 b | 0.30 b |
| control | 26.4 ab | 27.2 ab | 7.9 a | 13.9 a | 6.1 b | 1.04 ab | 2.38 a | 0.40 a |
| Potato | ||||||||
| L. arvalis | 29.4 a | 24.9 a | 5.2 b | 14.5 ab | 7.8 a | 0.88 a | 1.96 a | 0.24 b |
| T.virens | 27.9 ab | 26.6 a | 6.2 ab | 14.2 b | 7.9 a | 0.98 a | 2.05 a | 0.29 ab |
| B.subtilis | 26.6 ab | 25.0 a | 6.9 a | 15.5 a | 6.8 a | 0.96 a | 2.38 a | 0.34 a |
| control | 25.6 b | 24.6 a | 6.7 ab | 14.6 ab | 7.4 a | 0.97 a | 2.09 a | 0.32 ab |
| Location 2 (Northern Maine) | ||||||||
| Preceding Rotation Crop | ||||||||
| Barley/ryegrass | 29.7 a | 29.6 a | 24.1 a | 15.7 a | 13.8 b | 1.00 a | 1.18 a | 0.82 a |
| Barley/clover | 30.2 a | 29.4 a | 23.2 a | 16.0 a | 16.1 a | 0.98 a | 1.04 a | 0.73 b |
| Potato | 30.8 a | 28.7 a | 22.8 a | 15.5 a | 15.7 a | 0.93 b | 1.18 a | 0.74 b |
| Biocontrol Treatment | ||||||||
| L. arvalis | 30.7 a | 29.2 a | 23.5 ab | 15.5 b | 16.2 a | 0.96 a | 1.01 b | 0.75 a |
| T.virens | 30.1 a | 29.4 a | 23.9 a | 15.5 b | 15.3 ab | 0.98 a | 1.08 ab | 0.78 a |
| B.subtilis | 30.3 a | 29.2 a | 23.4 ab | 15.6 ab | 14.5 b | 0.97 a | 1.14 ab | 0.78 a |
| control | 30.0 a | 29.1 a | 22.9 b | 16.2 a | 14.8 ab | 0.98 a | 1.16 a | 0.74 a |
x These classes are composed of the proportion of fatty acids belonging to each structural class, where Sat = straight chain fatty acids (9:0, 10:0, 12:0, 13:0, 14:0, 15:0, 16:0), Mono = monounsaturated fatty acids (14:1 ω9c, 15:1 ω8c, 16:1 ω7c, 16:1 ω5c, 17:1 ω7c, 17:1 ω3c, 18:1 ω9c, 20:1 ω6c), Poly = polyunsaturated fatty acids (18:3 ω6c, 18:2 ω6c, 20:4 ω6c, 20:5 ω3c), Branch = saturated branched fatty acids (11:0 iso, 13:0 iso, 13:0 anteiso, 14:0 iso, 14:0 anteiso, 15:0 iso, 15:0 anteiso), Hydrox = fatty acids with hydroxyl groups (10:0 3OH, 12:0 2OH, 12:0 3OH, 15:0 iso 3OH, 16:0 2OH, 16:0 3OH). y Ratios are of structural class proportions that are used as biomarkers for specific groups of organisms. Mono/sat = the ratio of monounsaturated fatty acids to straight chain saturated fatty acids, Gram+/gram− = the ratio of branched to hydroxy fatty acids, and Fungi/bact = the ratio of polyunsaturated fatty acids to branched and hydroxyl fatty acids. z Means within the same column for each preceding rotation crop at location 1or preceding rotation crop or biocontrol treatment at location 2 followed by the same letter are not significantly different according to Fisher’s LSD (p = 0.05).
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Larkin, R.P.; Brewer, M.T. Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities. Agriculture 2020, 10, 128. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10040128
AMA Style
Larkin RP, Brewer MT. Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities. Agriculture. 2020; 10(4):128. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10040128
Chicago/Turabian StyleLarkin, Robert P., and Marin T. Brewer. 2020. "Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities" Agriculture 10, no. 4: 128. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10040128
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.