Glyphosate Resistance in Amaranthus viridis in Brazilian Citrus Orchards

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material and Seedling Propagation in Greenhouse
2.2. Glyphosate Dose-Response Assays
2.3. Shikimic Acid Accumulation
2.4. EPSPS Enzyme Activity Assays
2.5. Statistical Analysis
3. Results
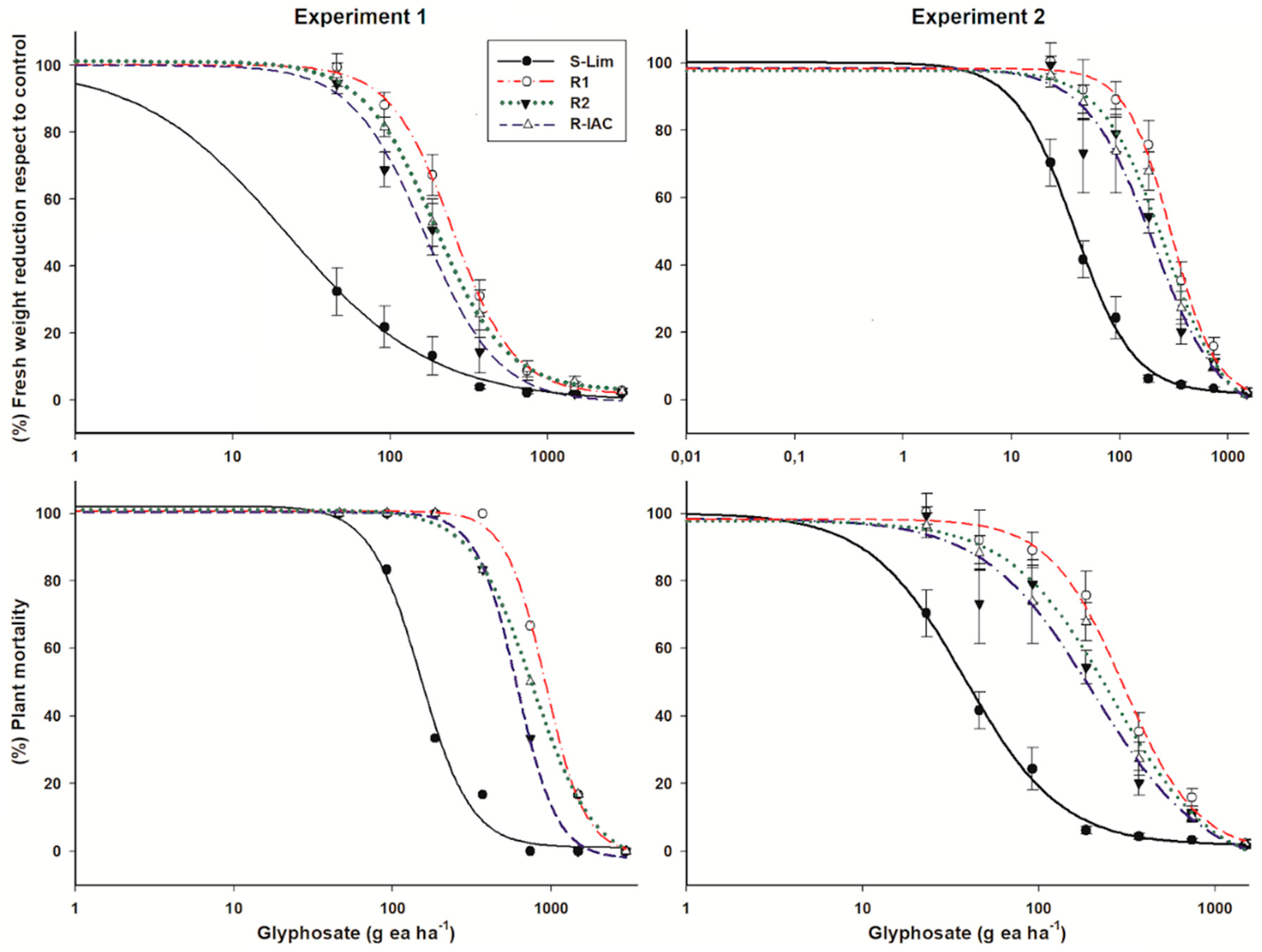
3.1. Fresh Weight Reduction and Plant Survival
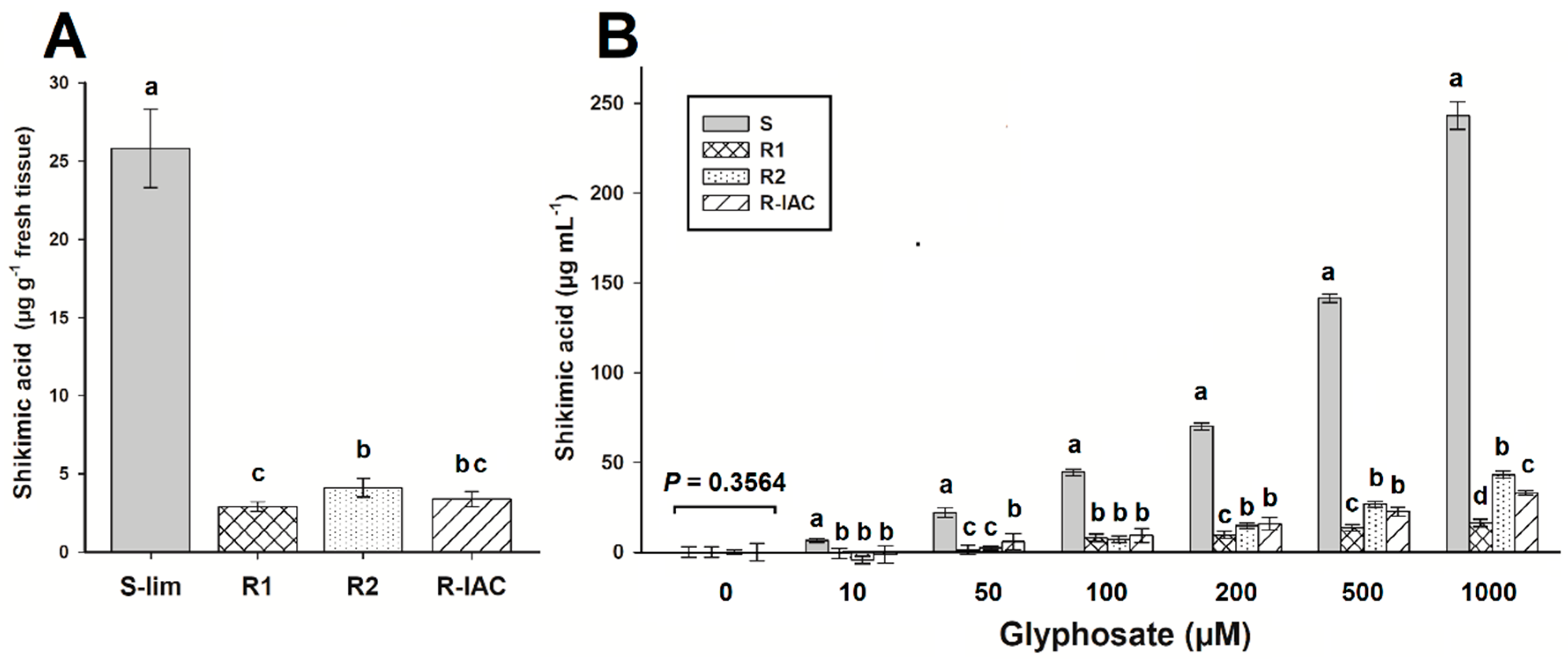
3.2. Shikimic Acid Accumulation
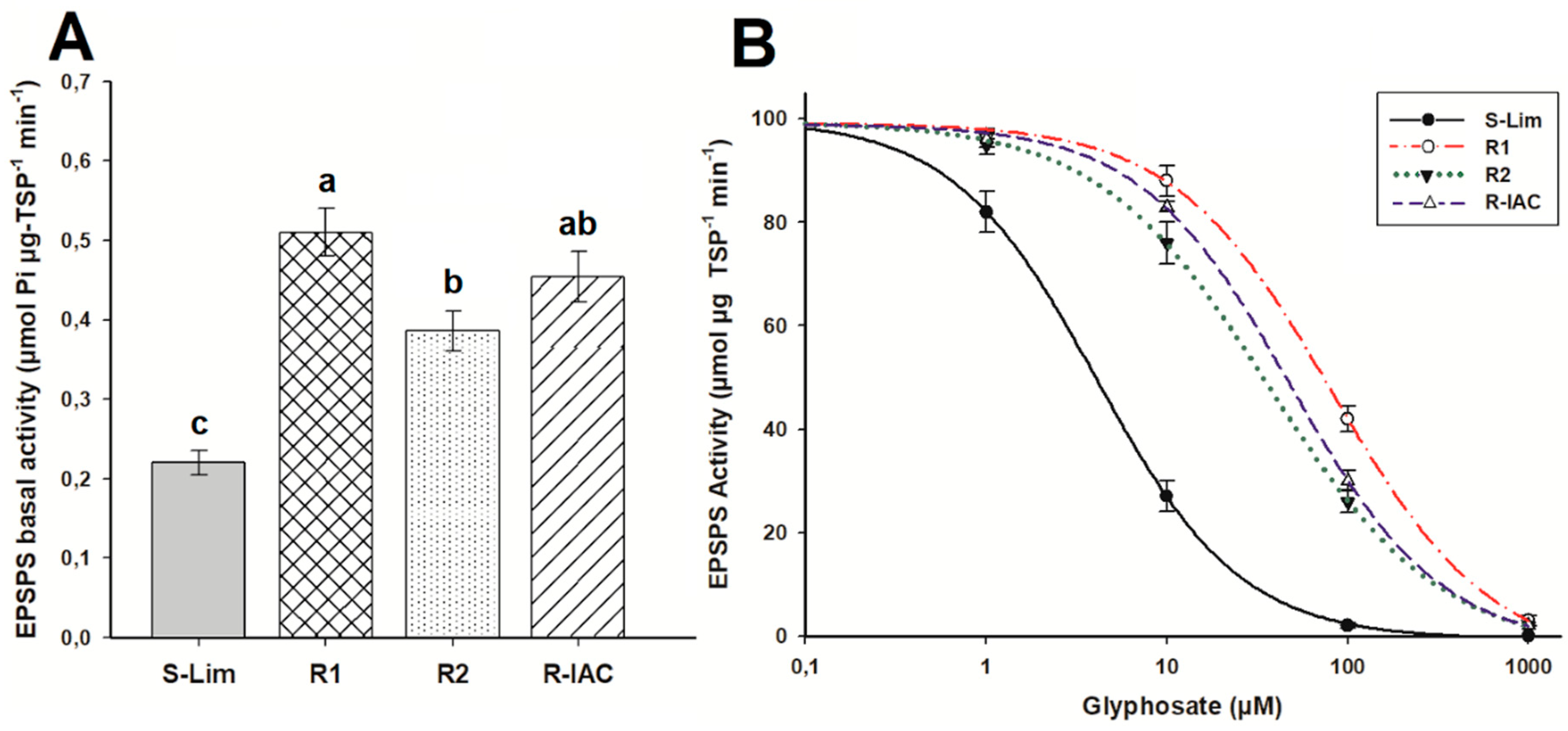
3.3. EPSPS Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- U.S. Department of Agriculture [USDA]. Citrus: World Markets and Trade. Available online: https://apps.fas.usda.gov/psdonline/circulars/citrus.pdf (accessed on 17 August 2019).
- Instituto Brasileiro de Geografia e Estatística [IBGE]. Produção Agrícola Municipal. 2017. Available online: http://www.sidra.ibge.gov.br/bda/tabela/protabl.asp?c=1613&z=p&o=18&i=p (accessed on 14 August 2019).
- Martinelli, R.; Monquero, P.A.; Fontanetti, A.; Conceição, P.M.; Azevedo, F.A. Ecological mowing: An option for sustainable weed management in young citrus orchards. Weed Technol. 2017, 31, 260–268. [Google Scholar] [CrossRef]
- Food and Agriculture Organization [FAO]. FAOSTAT: Statistical Database. Available online: http://faostat.fao.org/site/567/default.aspx (accessed on 17 August 2019).
- Singh, M.; Sharma, S.D. Benefits of triazine herbicides and other weed control technology in citrus management. In The Triazine Herbicides—50 Years Revolutionizing Agriculture, 1st ed.; Lebaron, H.M., McFarland, J.E., Burnside, O.C., Eds.; Elsevier: San Diego, CA, USA, 2008; pp. 199–209. [Google Scholar]
- La Cruz, R.A.-D.; Domínguez-Martínez, P.A.; Da Silveira, H.M.; Cruz-Hipólito, H.E.; Palma-Bautista, C.; Vazquez-Garcia, J.G.; Domínguez-Valenzuela, J.A.; De Prado, R. Management of glyphosate-resistant weeds in Mexican citrus groves: Chemical alternatives and economic viability. Plants 2019, 8, 325. [Google Scholar] [CrossRef] [Green Version]
- Bracamonte, E.; Da Silveira, H.M.; La Cruz, R.A.-D.; Domínguez-Valenzuela, J.A.; Cruz-Hipolito, H.E.; De Prado, R. From tolerance to resistance: Mechanisms governing the differential response to glyphosate in Chloris barbata. Pest Manag. Sci. 2018, 74, 1118–1124. [Google Scholar] [CrossRef]
- Caetano, R.S.X.; Christoffoleti, P.J.; Victoria-Filho, R. Weed seed bank of a ’Pera’ citrus orchard. Sci. Agric. 2001, 58, 509–517. [Google Scholar] [CrossRef]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef]
- Vázquez-García, J.G.; Castro, P.; Torra, J.; Alcántara-de la Cruz, R.; De Prado, R. Resistance evolution to EPSPS inhibiting herbicides in false barley (Hordeum murinum) harvested in Southern Spain. Agronomy 2020, 10, 992. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: http://www.weedscience.org (accessed on 30 June 2020).
- Moreira, M.S.; Nicolai, M.; Carvalho, S.J.P.; Christoffoleti, P.J. Glyphosate-resistance in Conyza canadensis and C. Bonariensis. Planta Daninha 2008, 25, 157–164. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, L.B.; Cruz-Hipólito, H.; González-Torralva, F.; Alves, P.; Christoffoleti, P.; De Prado, R. Detection of sourgrass (Digitaria insularis) biotypes resistant to glyphosate in Brazil. Weed Sci. 2011, 59, 171–176. [Google Scholar] [CrossRef]
- Alcántara-de la Cruz, R.; Oliveira, G.M.; Carvalho, L.B.; Silva, M.F.G.F. Herbicide resistance in Brazil: Status, impacts, and future challenges. In Herbicides-Current Research and Case Studies in Use, 2nd ed.; Ferreira, K.M., Ed.; IntechOpen: Londod, UK, 2020. [Google Scholar] [CrossRef] [Green Version]
- Sellers, B.A.; Smeda, R.J.; Johnson, W.G.; Kendig, J.A.; Ellersick, M.R. Comparative growth of six Amaranthus species in Missouri. Weed Sci. 2003, 51, 329–333. [Google Scholar] [CrossRef]
- García, M.J.; Palma-Bautista, C.; Rojano-Delgado, A.M.; Bracamonte, E.; Portugal, J.; La Cruz, R.A.-D.; De Prado, R. The triple amino acid substitution TAP-IVS in the EPSPS gene confers high glyphosate resistance to the superweed Amaranthus hybridus. Int. J. Mol. Sci. 2019, 20, 2396. [Google Scholar] [CrossRef] [Green Version]
- Thomas, W.E.; Burke, I.C.; Spears, J.F.; Wilcut, J.W. Influence of environmental factors on slender amaranth (Amaranthus viridis) germination. Weed Sci. 2006, 54, 316–320. [Google Scholar] [CrossRef]
- Raimondi, M.; Oliveira, J.R.; Constantin, J.; Rios, F.; Gemelli, A.; Raimondi, R. Dose-response curve to soil applied herbicides and susceptibility evaluation of different Amaranthus species using model identity. Planta Daninha 2015, 33, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Francischini, A.C.; Constantin, J.; Oliveira, R.S., Jr.; Santos, G.; Braz, G.B.P.; Dan, H.A. First report of Amaranthus viridis resistence to herbicides. Planta Daninha 2014, 32, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Küpper, A.; Borgato, E.A.; Patterson, E.; Netto, A.G.; Nicolai, M.; De Carvalho, S.J.P.; Nissen, S.J.; Gaines, T.A.; Christoffoleti, P. Multiple resistance to glyphosate and acetolactate synthase inhibitors in Palmer amaranth (Amaranthus palmeri) identified in Brazil. Weed Sci. 2017, 65, 317–326. [Google Scholar] [CrossRef]
- Cromartie, T.H.; Polge, N. Method of Detecting Shikimic Acid. U.S. Patent 006482654B1, 8 February 2002. Available online: https://patents.google.com/patent/US6482654B1/en (accessed on 30 June 2020).
- Dayan, F.; Owens, D.K.; Corniani, N.; Silva, F.M.L.; Watson, S.B.; Howell, J.; Shaner, D.L. Biochemical markers and enzyme assays for herbicide mode of action and resistance studies. Weed Sci. 2015, 63, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Escalada, M.; Gil-Monreal, M.; Zabalza, A.; Royuela, M. Characterization of the Amaranthus palmeri physiological response to glyphosate in susceptible and resistant populations. J. Agric. Food Chem. 2016, 64, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Lorentz, L.; Gaines, T.A.; Nissen, S.J.; Westra, P.; Strek, H.J.; Dehne, H.W.; Ruiz-Santaella, J.P.; Beffa, R. Characterization of glyphosate resistance in Amaranthus tuberculatus populations. J. Agric. Food Chem. 2016, 62, 8134–8142. [Google Scholar] [CrossRef]
- Agência de Defesa Agropecuária do Paraná [ADAPAR]. Trade Label of Roundup Original DI. Available online: http://www.adapar.pr.gov.br/arquivos/File/defis/DFI/Bulas/Herbicidas/rounduporiginaldi190118.pdf (accessed on 30 June 2020).
- Heap, I. Criteria for Confirmation of Herbicide-Resistant Weeds—With Specific Emphasis on Confirming Low Level Resistance. Available online: http://www.weedscience.org/Documents/ResistanceCriterion.pdf (accessed on 30 June 2020).
- Takano, H.K.; Oliveira, R.S., Jr.; Constantin, J.; Braz, G.B.P.; Gheno, E.A. Goosegrass resistant to glyphosate in Brazil. Planta Daninha 2017, 35, e017163071. [Google Scholar] [CrossRef] [Green Version]
- Alcántara-de la Cruz, R.; Fernández-Moreno, P.T.; Ozuna, C.V. Target and non-target site mechanisms developed by glyphosate-resistant hairy beggarticks (Bidens pilosa L.). Front. Plant Sci. 2016, 7, 1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, R.A.; Scott, R.C.; Dayan, F.E.; Burgos, N.R. EPSPS Gene amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp. multiflorum) populations from Arkansas (United States). J. Agric. Food Chem. 2015, 63, 5885–5893. [Google Scholar] [PubMed]
- Shaner, D.L.; Nadler-Hassar, T.; Henry, W.B.; Koger, C.H. A rapid in vivo shikimate accumulation assay with excised leaf discs. Weed Sci. 2005, 53, 769–774. [Google Scholar] [CrossRef]
- Gaines, T.A.; Zhang, W.; Wang, D.; Bukun, B.; Chisholm, S.T.; Shaner, D.L.; Nissen, S.J.; Patzoldt, W.L.; Tranel, P.J.; Culpepper, A.S.; et al. Gene amplification confers glyphosate resistance in Amaranthus palmeri. Proc. Natl. Acad. Sci. USA 2010, 107, 1029–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandula, V.K.; Ray, J.D.; Ribeiro, D.N.; Pan, Z.; Reddy, K.N. Glyphosate resistance in tall waterhemp (Amaranthus tuberculatus) from Mississippi is due to both altered target-site and nontarget-site mechanisms. Weed Sci. 2013, 61, 374–383. [Google Scholar] [CrossRef]
- Riggins, C.W.; Peng, Y.H.; Stewart, C.N.; Tranel, P.J. Characterization of de novo transcriptome for waterhemp (Amaranthus tuberculatus) using GS-FLX 454 pyrosequencing and its application for studies of herbicide target-site genes. Pest Manag. Sci. 2010, 66, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Gaines, T.A.; Patterson, E.L.; Neve, P. Molecular mechanisms of adaptive evolution revealed by global selection for glyphosate resistance. New Phytol. 2019, 223, 1770–1775. [Google Scholar] [CrossRef] [Green Version]
- Giacomini, D.A.; Westra, P.; Ward, S.M. Variable inheritance of amplified EPSPS gene copies in glyphosate-resistant Palmer Amaranth (Amaranthus palmeri). Weed Sci. 2019, 67, 176–182. [Google Scholar] [CrossRef]
- Yu, Q.; Jalaludin, A.; Han, H.; Chen, M.; Sammons, R.D.; Powles, S.B. Evolution of a double amino acid substitution in the 5-enolpyruvylshikimate-3-phosphate synthase in Eleusine indica conferring high-level glyphosate resistance. Plant Physiol. 2015, 167, 1440–1447. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pop. | Fresh Weight Reduction | Plant Mortality | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| c | d | b | GR50 ± CI | RF 2 | c | d | b | LD50 ± CI | RF 2 | |
| Experiment I | ||||||||||
| S-Lim | 0.3 | 99.9 | 0.9 | 22.0 ± 7.4 | - | 1.9 | 102.2 | 2.8 | 149.5 ± 11.8 | - |
| R1 | 1.8 | 100.2 | 2.2 | 248.4 ± 17.7 | 11.3 | 1.1 | 100.7 | 3.4 | 918.9 ± 31.4 | 6.1 |
| R2 | 0.6 | 100.1 | 1.9 | 165.9 ± 16.1 | 7.5 | 2.2 | 100.3 | 3.4 | 607.8 ± 15.1 | 4.1 |
| R-IAC | 2.9 | 101.3 | 1.9 | 192.2 ± 17.4 | 8.7 | 4.1 | 101.1 | 2.3 | 768.3 ± 32.6 | 5.1 |
| Experiment II | ||||||||||
| S-Lim | 1.7 | 100.0 | 1.6 | 38.1 ± 3.8 | - | 0.4 | 101.4 | 3.4 | 113.1 ± 8.1 | - |
| R1 | 0.4 | 98.4 | 2.1 | 301.8 ± 39.4 | 7.9 | 0 | 99.4 | 3.4 | 706.3 ± 45.4 | 6.2 |
| R2 | 5.5 | 98.4 | 1.4 | 205.5 ± 42.5 | 5.4 | 4.6 | 96.7 | 3.6 | 639.7 ± 75.1 | 5.7 |
| R-IAC | 3.7 | 97.9 | 1.5 | 259.5 ± 52.4 | 6.8 | 4.9 | 97.0 | 2.7 | 726.7 ± 59.2 | 6.4 |
| Pop. | c | d | b | I50 ± CI | RF 2 |
|---|---|---|---|---|---|
| S-Lim | 0.5 | 99.9 | 1.1 | 4.0 ± 0.3 | - |
| R1 | 1.1 | 99.2 | 0.9 | 82.4 ± 6.3 | 20.6 |
| R2 | 2.6 | 99.4 | 0.9 | 36.4 ± 3.6 | 9.1 |
| R-IAC | 2.1 | 98.9 | 1.0 | 48.4 ± 4.4 | 12.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcántara-de la Cruz, R.; Amaral, G.d.S.; Oliveira, G.M.d.; Rufino, L.R.; Azevedo, F.A.d.; Carvalho, L.B.d.; Silva, M.F.d.G.F.d. Glyphosate Resistance in Amaranthus viridis in Brazilian Citrus Orchards. Agriculture 2020, 10, 304. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070304
Alcántara-de la Cruz R, Amaral GdS, Oliveira GMd, Rufino LR, Azevedo FAd, Carvalho LBd, Silva MFdGFd. Glyphosate Resistance in Amaranthus viridis in Brazilian Citrus Orchards. Agriculture. 2020; 10(7):304. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070304
Chicago/Turabian StyleAlcántara-de la Cruz, Ricardo, Gabriel da Silva Amaral, Guilherme Moraes de Oliveira, Luiz Renato Rufino, Fernando Alves de Azevedo, Leonardo Bianco de Carvalho, and Maria Fátima das Graças Fernandes da Silva. 2020. "Glyphosate Resistance in Amaranthus viridis in Brazilian Citrus Orchards" Agriculture 10, no. 7: 304. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070304