
Occurrence and Distribution of Major Viruses Infecting Eggplant in Lebanon and Molecular Characterization of a Local Potato Virus X Isolate

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Serological Assay
2.3. Biological Assay
2.4. Electron Microscopy
2.5. Total Nucleic Acids Extraction, cDNA Synthesis and PCR
2.6. Cloning, Sequencing and Phylogenetic Analysis
3. Results
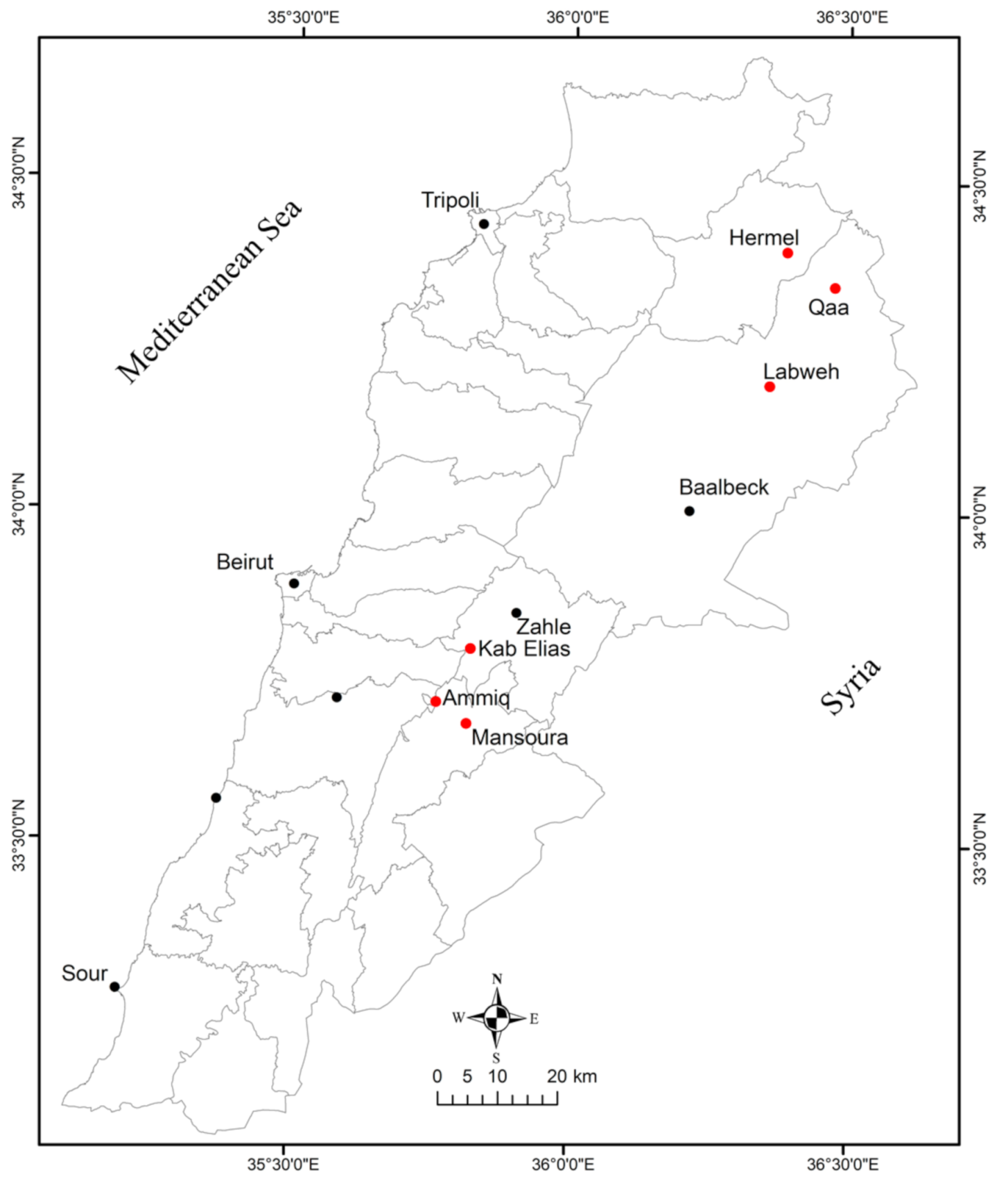
3.1. Survey, Visual Inspections and Virus Distribution in Bekaa Valley
3.2. Biological Assay
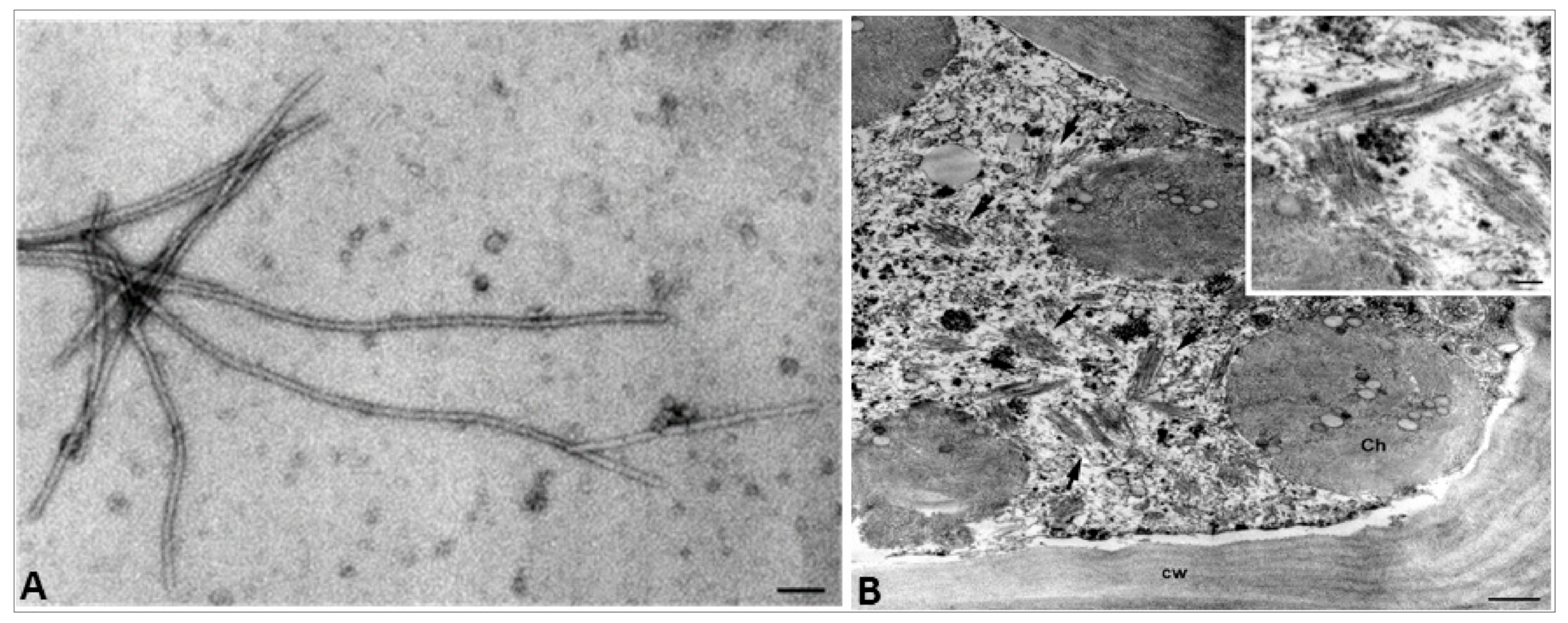
3.3. Electron Microscopy
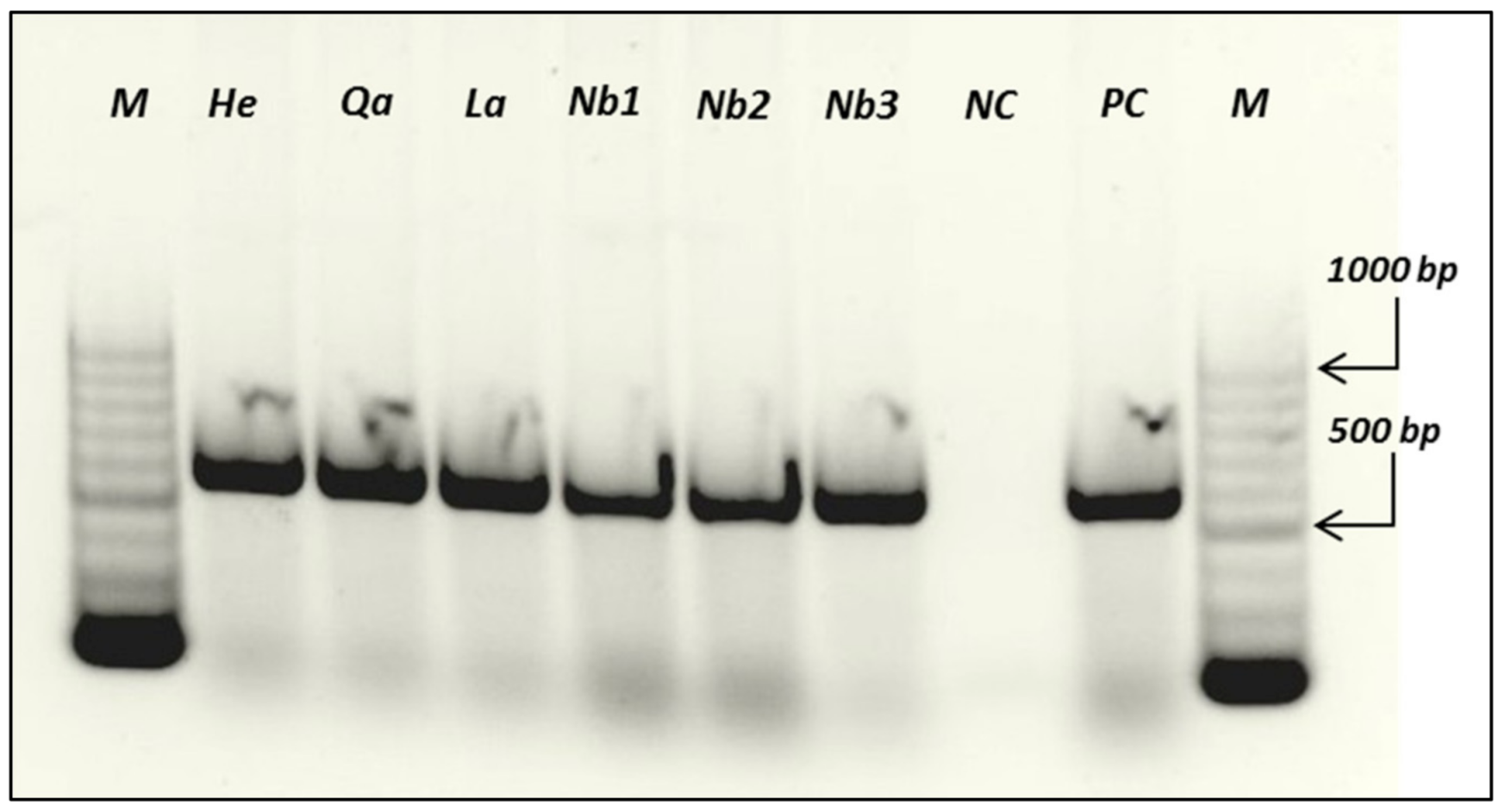
3.4. Molecular Detection of PVX by RT-PCR
3.5. Association of Virus Infection with Plant Symptoms
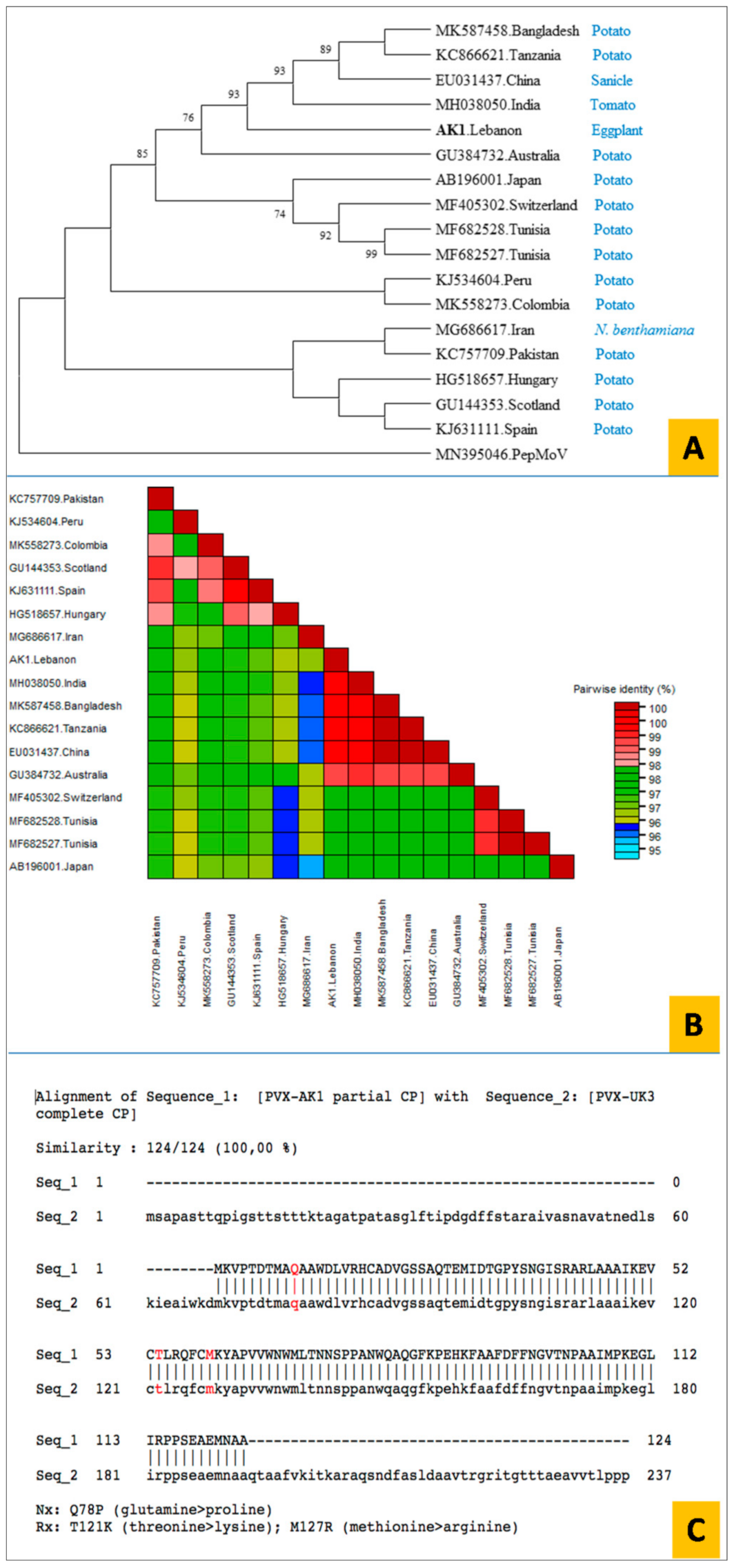
3.6. Cloning and Sequence Analysis of the PVX CP Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ministry of Agriculture (MoA) of Lebanon, FAO. Agriculture in Lebanon 2008–2009; Technical Report; Ministry of Agriculture/Food and Agriculture Organization: Beirut, Lebanon, 2010.
- Nassif, M.H. Groundwater Governance in Central Bekaa, Lebanon. In IWMI Project Report, Groundwater Governance in the Arab World–Taking Stock and Addressing Challenges; Report No. 9; IWMI: Beirut, Lebanon, 2016. [Google Scholar]
- Anonymous. Résultats Globaux du Module de Base du Recensement de l’agriculture 2010; Projet Observatoire Libanais de Développement Agricole: Beirut, Lebanon, 2012; p. 140. [Google Scholar]
- Beemster, A.B.R.; Rozendaal, A. Potato viruses: Properties and symptoms. In Viruses of Potatoes and Seed-Potato Production; De Borkz, J.A., Ed.; Pubdoc: Wageningen, The Netherlands, 1972; pp. 115–143. [Google Scholar]
- Hančinský, R.; Mihálik, D.; Mrkvová, M.; Candresse, T.; Glasa, M. Plant viruses Infecting solanaceae family members in the cultivated and wild environments: A review. Plants 2020, 9, 667. [Google Scholar] [CrossRef]
- Banttari, E.E.; Ellis, J.P.; Kurana, S.M.P. Diseases caused by viruses and virus-like pathogens. In Potato Health Management; Rowe, R.C., Ed.; APS PRESS: St. Paul, MN, USA, 1993; pp. 127–133. [Google Scholar]
- Abou-Jawdeh, Y.; Sobh, H.; Saad, A.T. Incidence of potato virus diseases and their significance for a seed certification program in Lebanon. Phytopathol. Mediterr. 2001, 40, 1113–1118. [Google Scholar]
- Choueiri, E.; El-Zammar, S.; Jreijiri, F.; Saad, A.T.; Afram, M.A.; Varveri, C. Records of potato viruses in Lebanon. J. Plant Pathol. 2002, 84, 139. [Google Scholar]
- Choueiri, E.; El-Zammar, S.; Jreijiri, F.; Mnayer, D.; Massaad, R.; Saad, A.T.; Hanna, L.; Varveri, C. Phytosanitary status of potato in Bekaa valley in Lebanon. Bull. OEPP 2004, 34, 117–121. [Google Scholar] [CrossRef]
- Abou-Jawdah, Y.; El Mohtar, C.; Sobh, H. First report of tomato spotted wilt virus on tomatoes in Lebanon. Plant Dis. 2006, 90, 378. [Google Scholar] [CrossRef] [PubMed]
- Fidan, H.; Sarıkaya, P. Tomato chlorosis virus and tomato yellow leaf curl virus causing mixed infection in protected eggplant (S. melongena) crops in Turkey. Acta Scientiarum polonorum. Hortorum. Cultus. 2020, 19, 81–89. [Google Scholar]
- Mansour, A.; Almusa, A.; Vetten, H.V.; Lesemann, D.E. Properties of a Cowpea mild Mottle virus (CPMMV) isolate from eggplant in Jordan and evidence for biological and serological differences between CPMMV Isolates from leguminous and solanaceous hosts. J. Phytopathol. 2008, 146, 539–547. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Sapkota, R.; Lachman, O.; Pearlsman, M.; Antignus, Y. A new aubergine disease caused by a whitefly-borne strain of tomato mild mottle virus (TomMMoV). Plant Pathol. 2013, 62, 750–759. [Google Scholar] [CrossRef]
- Sadeghi, M.S.; Behjatnia, S.A.A.; Masumi, M.; Izadpanah, K. Characterisation of a strain of Potato virus Y causing eggplant mosaic in southern Iran. Australas Plant Pathol. 2008, 37, 79–86. [Google Scholar] [CrossRef]
- Tsai, W.S.; Abdourhamane, I.K.; Kenyon, L. First Report of Pepper veinal mottle virus associated with mosaic and mottle diseases of tomato and pepper in mali. Plant Dis. 2010, 94, 378. [Google Scholar] [CrossRef]
- Rakib, A.; Mustafa, A.A.; Adhab, A.; Ismail, K.A.H. Eggplant blister mottled virus (EBMV): A possible new potyvirus characterized from Iraq. J. Gen. Mol. Virol. 2011, 3, 49–52. [Google Scholar]
- Bagewadi, B.; Hossain, M.S.; Fayad, A.; Naidu, R.A. First report of cucumber mosaic virus from eggplant (S. melongena) in Bangladesh. Plant Dis. 2015, 99, 293. [Google Scholar] [CrossRef] [PubMed]
- Rui, P.H.; Jiang, L.; Li, S.; Jiang, X.Z.; Zhao, Q.Q.; Ying Feng, J.; Jiang, T. First report of broad bean wilt virus 2 infection in eggplant in China. J. Plant Pathol. 2020, 102, 543. [Google Scholar] [CrossRef] [Green Version]
- Sokhansanj, Y.; Rakhshandehroo, F.; Pourrahim, R. First report of tomato ringspot virus on eggplant in Iran. J. Plant Pathol. 2012, 94, S4.94. [Google Scholar] [CrossRef]
- Sastry, K.S.; Nayudu, M.V. Ringspot symptoms of eggplant, incited by Tobacco ring spot virus/Sintomi di maculatura anulare su Melanzana causati dal virus delta maculatura anulare del Tobacco. Phytopathol. Mediterr. 1976, 15, 60–62. [Google Scholar]
- Ozdemir, S.; Erilmez, S.; Payland, I.C. First report of Alfalfa mosaic virus in eggplant in Turkey. J. Plant Pathol. 2011, 93, S4.82. [Google Scholar]
- Blystad, D.A.; van der Vlugt, R.; Alfaro-Fernández, A.; del Carmen Córdoba, M.; Bese, G.; Hristova, D.; Pospieszny, H.; Mehle, N.; Ravnikar, M.; Tomassoli, L.; et al. Host range and symptomatology of Pepino mosaic virus strains occurring in Europe. Eur. J. Plant Pathol. 2015, 143, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Martelli, G.P.; Adams, M.J.; Kreuze, J.F.; Dolja, V.V. Family Flexiviridae: A case study in virion and genome plasticity. Ann. Rev. Phytopathol. 2007, 45, 73–100. [Google Scholar] [CrossRef] [Green Version]
- He, Y.H.; Huang, K.; Yao, L.Z.; Li, W.P.; Liu, C.M.; Chen, L.; Chen, H.R.; Chen, S.Y.; Wang, J.G. First report of tobacco mosaic virus infecting dutch eggplant (Solanum aculeatissimum) in China. Plant Dis. 2019, 103, 2973. [Google Scholar] [CrossRef]
- Nawaz, H.H.; Umer, M.; Bano, S.; Usmani, A.; Naseer, M. A Research review on tomato bushy stunt virus disease complex. J. Nat. Sci. Res. 2014, 4, 18–23. [Google Scholar]
- Kamberoglu, M.A.; Caliskan, A.F.; Alan, B. First report of tomato spotted wilt virus on eggplant in Turkey. J. Plant Pathol. 2009, 91, 231. [Google Scholar]
- Martelli, G.P. Bacilliform particles associated with mottled dwarf of eggplant (S. melongena L.). J. Gen. Virol. 1969, 5, 319–320. [Google Scholar] [CrossRef]
- Brioso, P.S.T.; Pimentel, J.P.; Louro, R.P.; Kitajima, E.W.; Oliveira, D.E. Andean potato mottle virus—Characterization of a strain naturally infecting eggplant (S. melongena). Fitopatol. Bras. 1993, 18, 526–533. [Google Scholar]
- Briand, J.P.; Bouley, J.P.; Witz, J. Self-assembly of eggplant mosaic virus protein. Virology 1997, 76, 664–669. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Pearlsman, M.; Lachman, O.; Antignus, Y. Characterization of a new strain of eggplant mottled crinkle virus (EMCV) infecting eggplants in Israel. Phytoparasitica 2009, 37, 477–483. [Google Scholar] [CrossRef]
- Ravanbod, E.; Rakhshandehroo, F.; Golnaraghi, A. Survey of potato virus X in vegetable fields of Iran. J. Plant Pathol. 2018, 100, 137. [Google Scholar] [CrossRef] [Green Version]
- Hahm, Y.; Slack, S.A.; Slattery, R.J. Reinfection of potato seed stocks with potato virus S and potato virus X in Wisconsin. Am. Potato J. 1981, 58, 117–125. [Google Scholar] [CrossRef]
- Syller, J. Facilitative and antagonistic interactions between plant viruses in mixed infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef]
- Nie, X.; Singh, M. Response of potato, tobacco and Physalis floridana plants to mixed infection with PVX, PVYNTN and PVY strains. Can. J. Plant Pathol. 2013, 35, 390–401. [Google Scholar] [CrossRef]
- Liang, Z.; Dickison, V.; Singh, M.; Xiong, X.; Nie, X. Studies of tomato plants in response to infections with PVX and different PVY isolates reveal a remarkable PVX-PVYNTN synergism and diverse expression profiles of genes involved in different pathways. Eur. J. Plant Pathol. 2016, 144, 55–71. [Google Scholar] [CrossRef]
- Skryabin, K.G.; Kraev, A.S.; Morozov, S.Y.; Rozanov, M.; Chernov, B.K.; Lukasheva, L.I.; Atabekovet, J.G. The nucleotide sequence of potato virus X RNA. Nucleic Acids Res. 1988, 16, 10929–10930. [Google Scholar] [CrossRef]
- Huisman, M.J.; Linthorst, H.J.; Bol, J.F.; Cornelissen, J.C. The complete nucleotide sequence of potato virus X and its homologies at the amino acid level with various plus-stranded RNA viruses. J. Gen. Virol. 1988, 69, 1789–1798. [Google Scholar] [CrossRef]
- Orman, B.E.; Celnik, R.M.; Mandel, A.M.; Torres, H.N.; Mentaberry, A.N. Complete cDNA sequence of a South American isolate of potato virus X. Virus Res. 1990, 16, 293–305. [Google Scholar] [CrossRef]
- Scholthof, K.G.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of microplate method of enzyme-linked immunosorbent assay for detection of plant viruses. J. Gen.Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P.; Russo, M. Use of thin sectionning for the visualization and identification of plant viruses. J. Virol. Methods 1984, 8, 143–224. [Google Scholar]
- Nie, X.; Singh, R.P. A novel usage of random primers for multiplex RT-PCR detection of virus and viroid in aphids, leaves, and tubers. J. Virol. Methods 2001, 91, 37–49. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Daunay, M.C.; Caranta, C.; Pédron, F.; Deulvot, C.; Dussault, M.; Moretti, A.; Jullian, E.; Ruffel, S.; Moquet, F.; Majde, M.; et al. Heredity of Tomato mosaic virus (ToMV) and Potato virus Y (PVY) resistance in eggplant (S. melongena L.). In Proceedings of the XIIth Meeting on Genetics and Breeding of Capsicum and Eggplant, Noordwijkerhout, The Netherlands, 17–19 May 2004; pp. 154–160. [Google Scholar]
- Colak-Ates, A.; Fidan, H.; Ozarslandan, A.; Ata, A. Determination of the resistance of certain eggplant lines against Fusarium wilt, potato Y potyvirus and root-knot nematode using molecular and classic methods. Fresenius Environ. Bull. 2018, 27, 7446–7453. [Google Scholar]
- Pappi, P.G.; Chaintoutis, S.C.; Dovas, C.I.; Efthimiou, K.E.; Katis, N.I. Development of one-tube real-time qRT-PCR and evaluation of RNA extraction methods for the detection of eggplant mottled dwarf virus in different species. J. Virol. Methods 2015, 212, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Babaie, G.; Kouhi-Habibi, M.; Massah, A.; Dizadji, A.; Izadinejad, L.; Simon, A. Complete genome sequence and genome analysis of eggplant mottled dwarf virus-Iranian isolate. J. Phytopathol. 2015, 163, 331–341. [Google Scholar] [CrossRef]
- Babaie, G.; Izadpanah, K. Vector transmission of eggplant mottled dwarf virus in Iran. J. Phytopathol. 2003, 151, 679–682. [Google Scholar] [CrossRef]
- Dietzgen, R.G.; Mann, K.S.; Johnson, K.N. Plant virus-insect vector interactions: Current and potential future research directions. Viruses 2016, 8, 303. [Google Scholar] [CrossRef]
- Zechmann, B.; Zellnig, G. Rapid diagnosis of plant virus diseases by transmission electron microscopy. J. Virol. Methods 2009, 162, 163–169. [Google Scholar] [CrossRef]
- Richert-Pöggeler, K.R.; Franzke, K.; Hipp, K.; Kleespies, R.G. Electron microscopy methods for virus diagnosis and high resolution analysis of viruses. Front. Microbiol. 2019, 9, 3255. [Google Scholar] [CrossRef]
- Roossinck, M.J.; García-Arenal, F. Ecosystem simplification, biodiversity loss and plant virus emergence. Curr. Opin. Virol. 2015, 10, 56–62. [Google Scholar] [CrossRef]
- De Bokx, J.A. Potato virus Y. In Compendium of Potato Diseases; Hooker, W.J., Ed.; American Phytopathological Society: St. Paul, MN, USA, 1986; pp. 70–71. [Google Scholar]
- Rochow, W.F.; Ross, A.F. Virus multiplication in plants doubly infected by potato viruses X and Y. Virology 1955, 1, 10–27. [Google Scholar] [CrossRef]
- Vance, V.B. Replication of Potato virus X RNA is altered in coinfections with Potato virus Y. Virology 1991, 182, 486–494. [Google Scholar] [PubMed]
- Gonzalez-Jara, P.; Tenllado, F.; Martinez-Garcia, B.; Atencio, F.A.; Barajas, D.; Vargas, M.; Diaz-Ruiz, J.; Diaz-Ruiz, J.R. Host-dependent differences during synergistic infection by Potyviruses with Potato virus X. Mol. Plant Pathol. 2004, 5, 29–35. [Google Scholar] [CrossRef]
- Hameed, A.; Iqbal, Z.; Asad, S.; Mansoor, S. Detection of multiple Potato viruses in the field suggests synergistic interactions among Potato viruses in Pakistan. Plant Pathol. J. 2014, 30, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Cockerham, G. Genetical studies on resistance to potato viruses X and Y. Heredity 1970, 25, 309–348. [Google Scholar] [CrossRef]
- Goulden, M.G.; Köhm, B.A.; Santa Cruz, S.; Kavanagh, T.A.; Baulcombe, D.C. A feature of the coat protein of potato virus X affects both induced virus resistance in potato and viral fitness. Virology 1993, 197, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Santa Cruz, S.; Baulcombe, D. Analysis of potato virus X coat protein genes in relation to resistance conferred by the genes Nx, Nb and Rx1 of potato. J. Gen. Virol. 1995, 76, 2057–2061. [Google Scholar] [CrossRef]
- Querci, M.; Baulcombe, D.C.; Goldbach, R.W.; Salazar, L.F. Analysis of the resistance-breaking determinants of potato virus X (PVX) strain HB on different potato genotypes expressing extreme resistance to PVX. Phytopathology 1995, 85, 1003–1010. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Species | Natural Transmission | Reference | |
|---|---|---|---|---|---|
| Geminiviridae | Begomovirus | Tomato yellow leaf curl virus | TYLCV | Whiteflies | [11] |
| Closteroviridae | Crinivirus | Tomato chlorosis virus | ToCV | Whiteflies | [11] |
| Betaflexiviridae | Carlavirus | Cowpea mild mottle virus | CPMMV | Whiteflies | [12] |
| Potyviridae | Ipomovirus | Tomato mild mottle virus | TomMMoV | Whiteflies | [13] |
| Potyviridae | Potyvirus | Potato virus Y | PVY | Aphids | [14] |
| Potyviridae | Potyvirus | Pepper veinal mottle virus | PVMV | Aphids | [15] |
| Potyviridae | Potyvirus | Eggplant blister mottled virus | EBMV | Aphids | [16] |
| Bromoviridae | Cucumovirus | Cucumber mosaic virus | CMV | Aphids | [17] |
| Secoviridae | Fabavirus | Broad bean wilt virus | BBWV | Aphids | [18] |
| Secoviridae | Nepovirus | Tomato ringspot virus | ToRSV | Nematodes | [19] |
| Secoviridae | Nepovirus | Tobacco ringspot virus | TRSV | Nematodes | [20] |
| Bromoviridae | Alfamovirus | Alfalfa mosaic virus | AMV | Aphids/seeds | [21] |
| Alphaflexiviridae | Potexvirus | Pepino mosaic virus | PepMV | Contact/seeds | [22] |
| Alphaflexiviridae | Potexvirus | Potato virus X | PVX | Contact/seeds | [23] |
| Virgaviridae | Tobamovirus | Tobacco mosaic virus | TMV | Contact/seeds | [24] |
| Tombusviridae | Tombusvirus | Tomato bushy stunt virus | TBSV | Seed | [25] |
| Tospoviridae | Orthotospovirus | Tomato spotted wilt orthotospovirus | TSWV | Thrips | [26] |
| Rhabdoviridae | Alphanucleorhabdovirus | Eggplant mottled dwarf alphanucleorhabdovirus | EMDV | Leafhoppers | [27] |
| Secoviridae | Comovirus | Andean potato mottle virus | APMoV | Beetles/contact | [28] |
| Tymoviridae | Tymovirus | Eggplant mosaic virus | EMV | Beetles | [29] |
| Tombusviridae | Tombusvirus | Eggplant mottled crinkle virus | EMCV | Unknown | [30] |
| ELISA-Positive Samples | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | No. Fields | Mean Virus Incidence (%visual) | No. Tested | No. Detected | EMDV | PepMoV | TSWV | AMV | CMV | PVY | ELISA Positive (%) |
| Hermel | 3 | 35 | 50 | 9 | 2 | 0 | 0 | 0 | 0 | 7 | 18% |
| Qaa | 2 | 30 | 40 | 10 | 2 | 0 | 0 | 0 | 0 | 8 | 25% |
| Labweh | 1 | 20 | 20 | 3 | 0 | 0 | 0 | 0 | 0 | 3 | 15% |
| Kab Elias | 1 | 10 | 20 | 3 | 1 | 0 | 0 | 0 | 0 | 2 | 15% |
| Mansoura | 1 | 10 | 20 | 5 | 0 | 0 | 0 | 0 | 2 | 3 | 25% |
| Ammiq | 1 | 10 | 20 | 3 | 0 | 0 | 0 | 0 | 0 | 3 | 15% |
| Total | 9 | 170 | 33 | 5 | 0 | 0 | 0 | 2 | 26 | ||
| (19.4%) | (2.9%) | (1.2%) | (15.3%) | ||||||||
| Region | Location | No. Tested | No. PVX+ | RT-PCR Detection |
|---|---|---|---|---|
| Northern Bekaa | Hermel | 25 | 15 | 60% |
| Qaa | 20 | 9 | 45% | |
| Labweh | 10 | 6 | 60% | |
| Central Bekaa | Kab Elias | 10 | 0 | 0.0 |
| Western Bekaa | Mansoura | 10 | 0 | 0.0 |
| Ammiq | 10 | 0 | 0.0 | |
| Total | 85 | 30 | 35.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou Kubaa, R.; Choueiri, E.; De Stradis, A.; Jreijiri, F.; Saponari, M.; Cillo, F. Occurrence and Distribution of Major Viruses Infecting Eggplant in Lebanon and Molecular Characterization of a Local Potato Virus X Isolate. Agriculture 2021, 11, 126. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020126
Abou Kubaa R, Choueiri E, De Stradis A, Jreijiri F, Saponari M, Cillo F. Occurrence and Distribution of Major Viruses Infecting Eggplant in Lebanon and Molecular Characterization of a Local Potato Virus X Isolate. Agriculture. 2021; 11(2):126. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020126
Chicago/Turabian StyleAbou Kubaa, Raied, Elia Choueiri, Angelo De Stradis, Fouad Jreijiri, Maria Saponari, and Fabrizio Cillo. 2021. "Occurrence and Distribution of Major Viruses Infecting Eggplant in Lebanon and Molecular Characterization of a Local Potato Virus X Isolate" Agriculture 11, no. 2: 126. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020126