
Effective Seed Yield and Flowering Synchrony of Parents of CIMMYT Three-Way-Cross Tropical Maize Hybrids

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Materials and Experimental Locations
2.2. Details of Experiments
2.3. Data Collection and Analysis
3. Results and Discussion
3.1. Parental Seed Management
3.1.1. Male Inbred Lines
3.1.2. Female Single Crosses
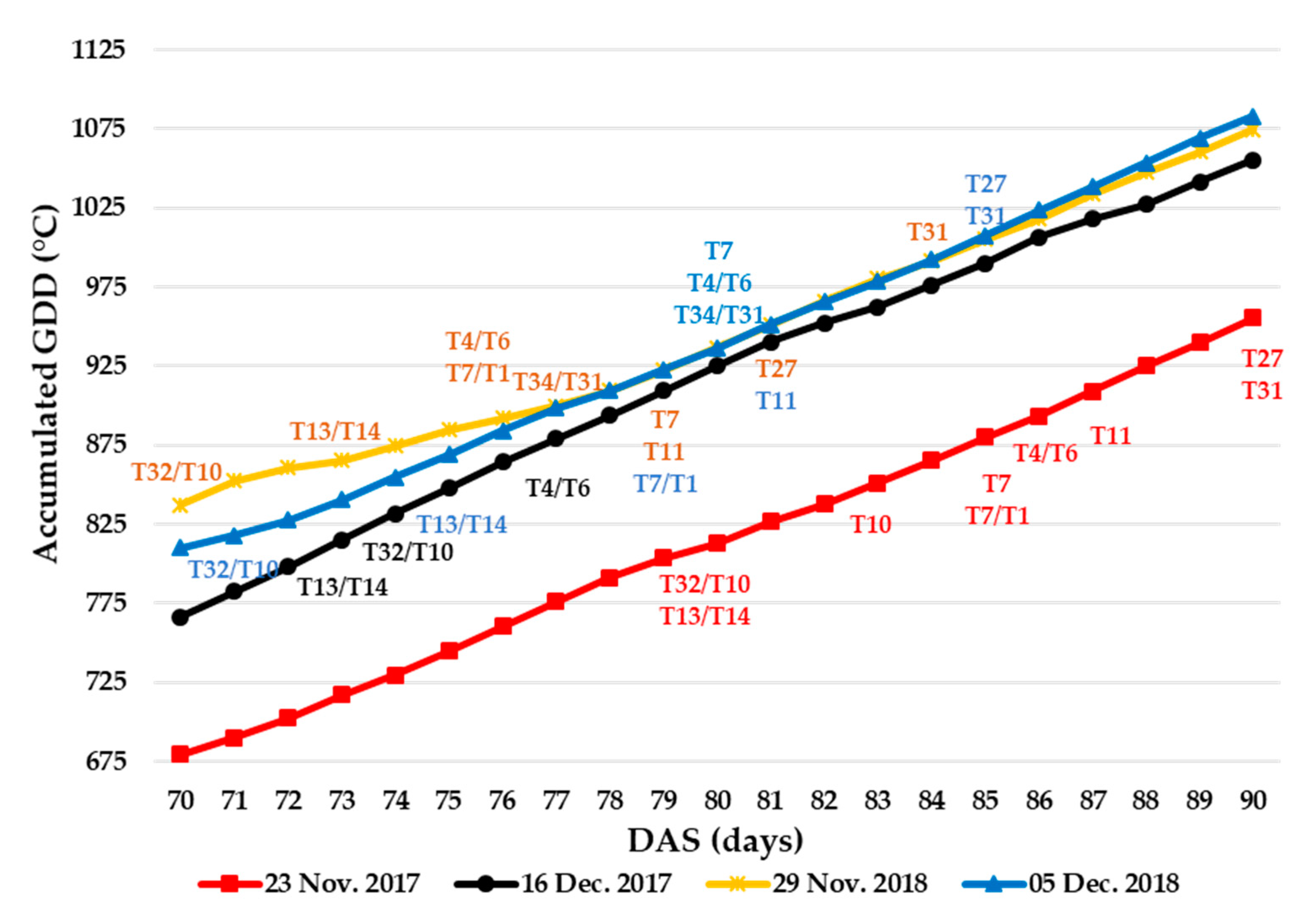
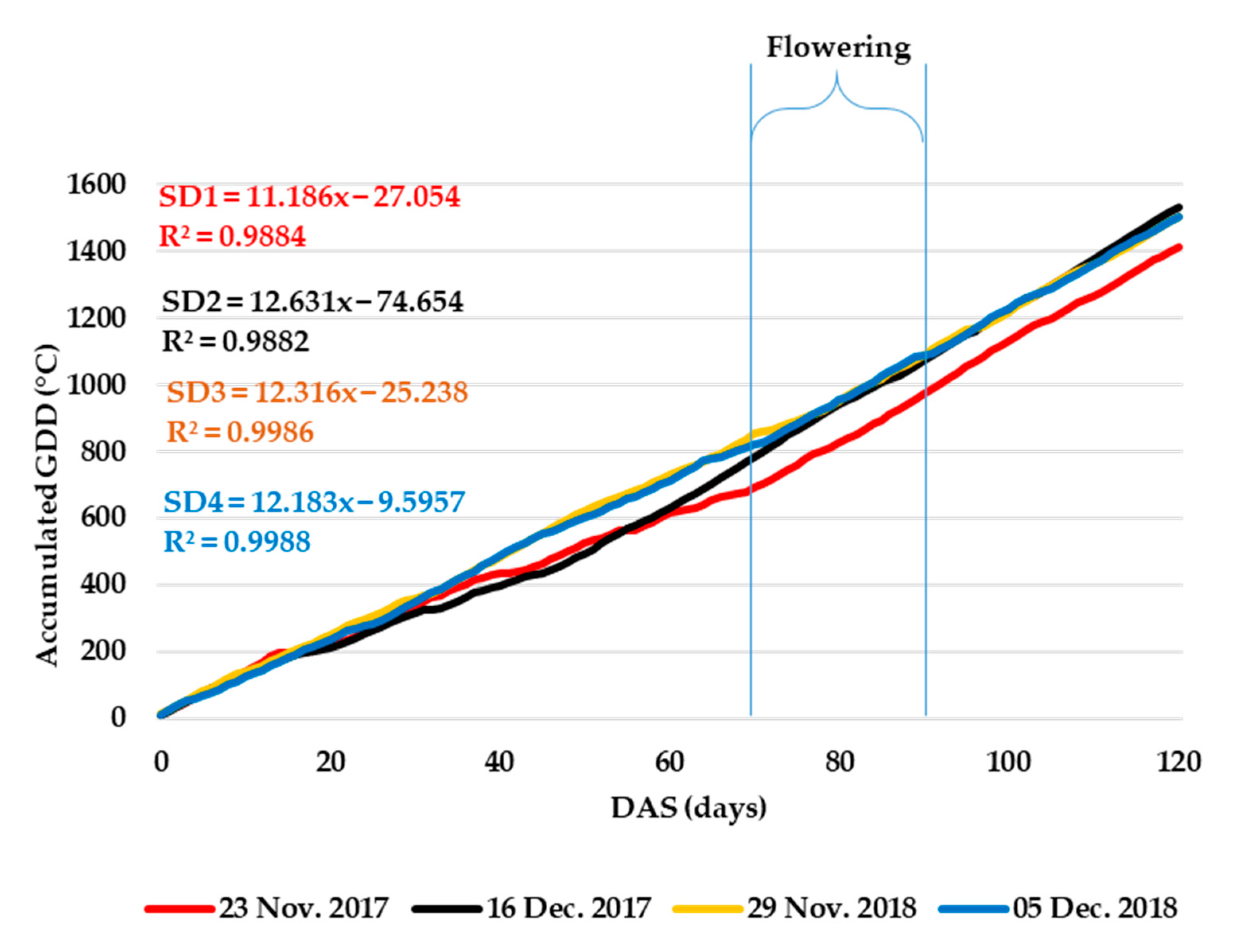
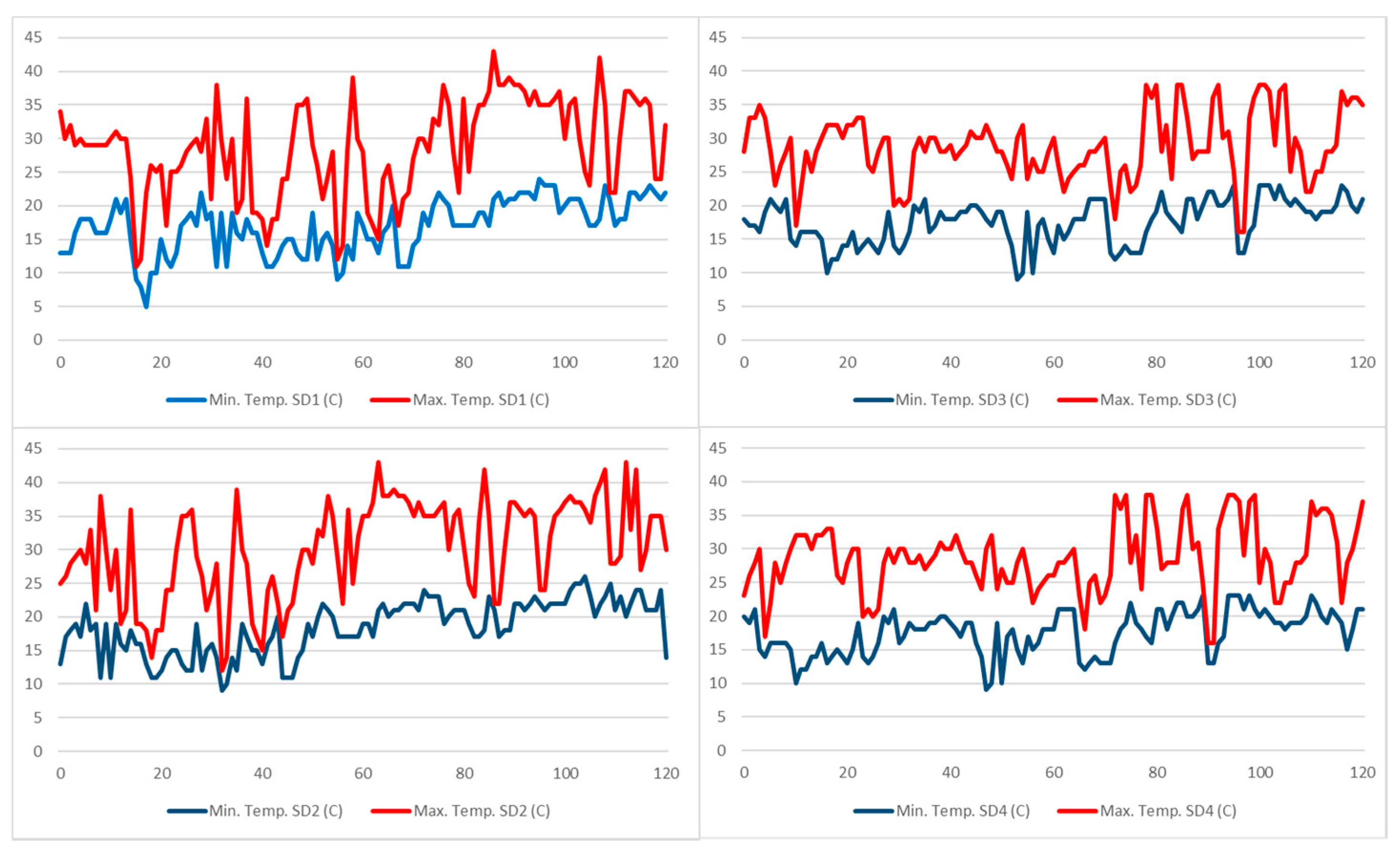
3.1.3. Relationship between Sowing Dates, Growing Degree Days, Floral Phenology, and Seed Yield
3.2. Hybrid Seed Production
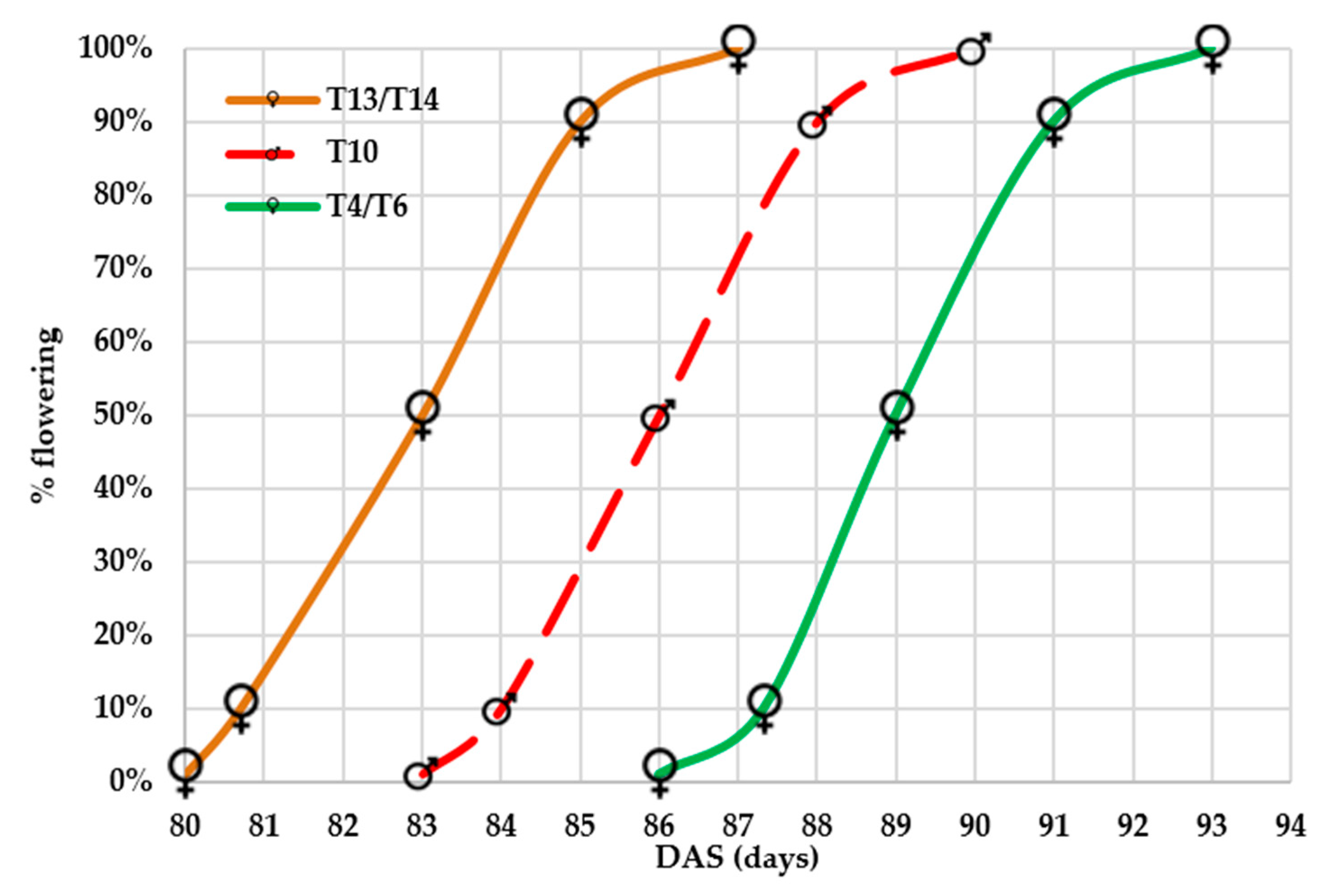
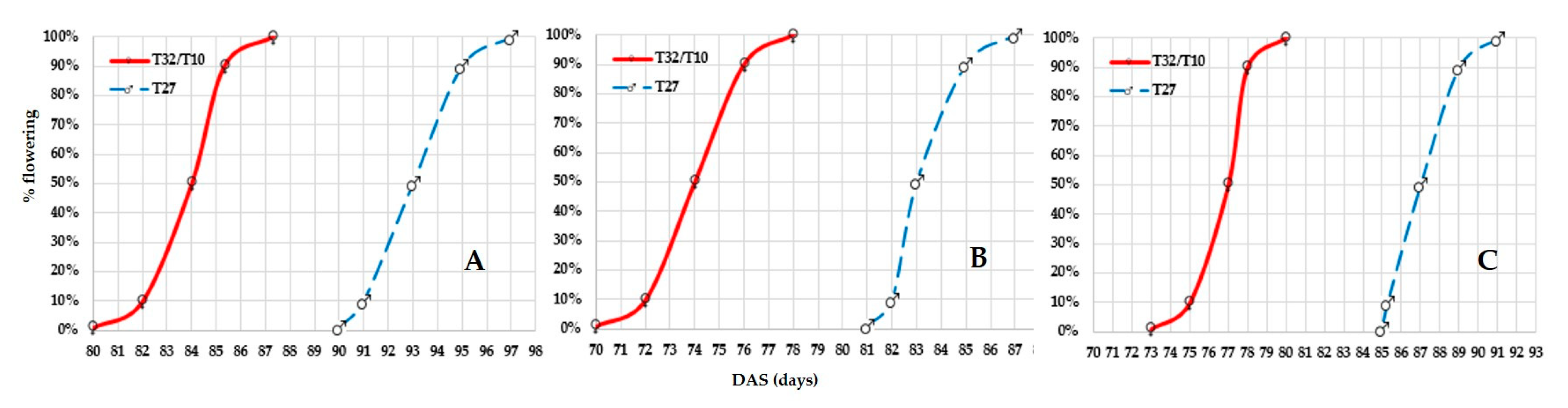
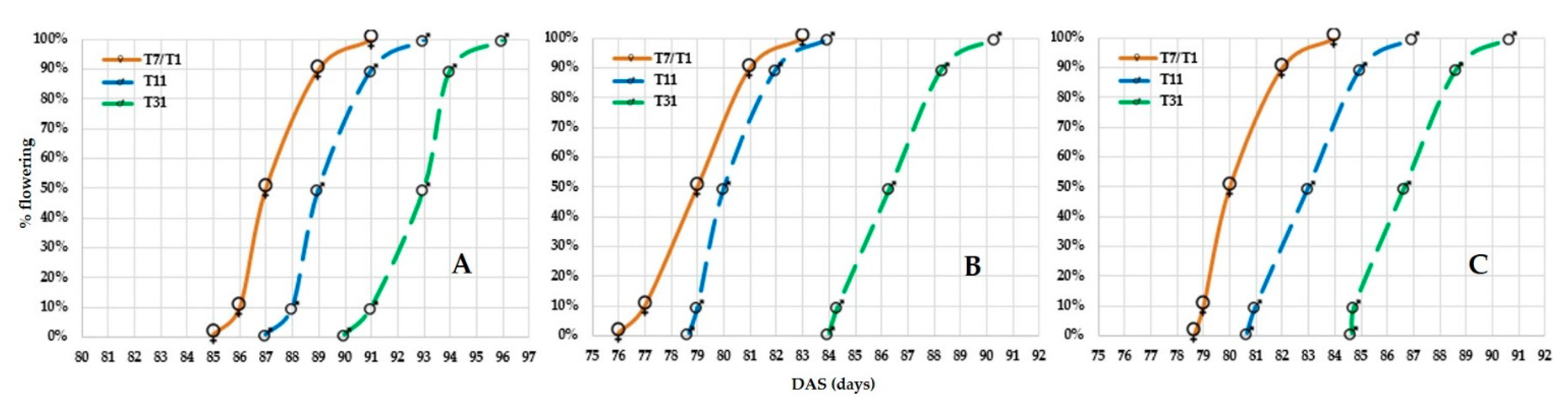
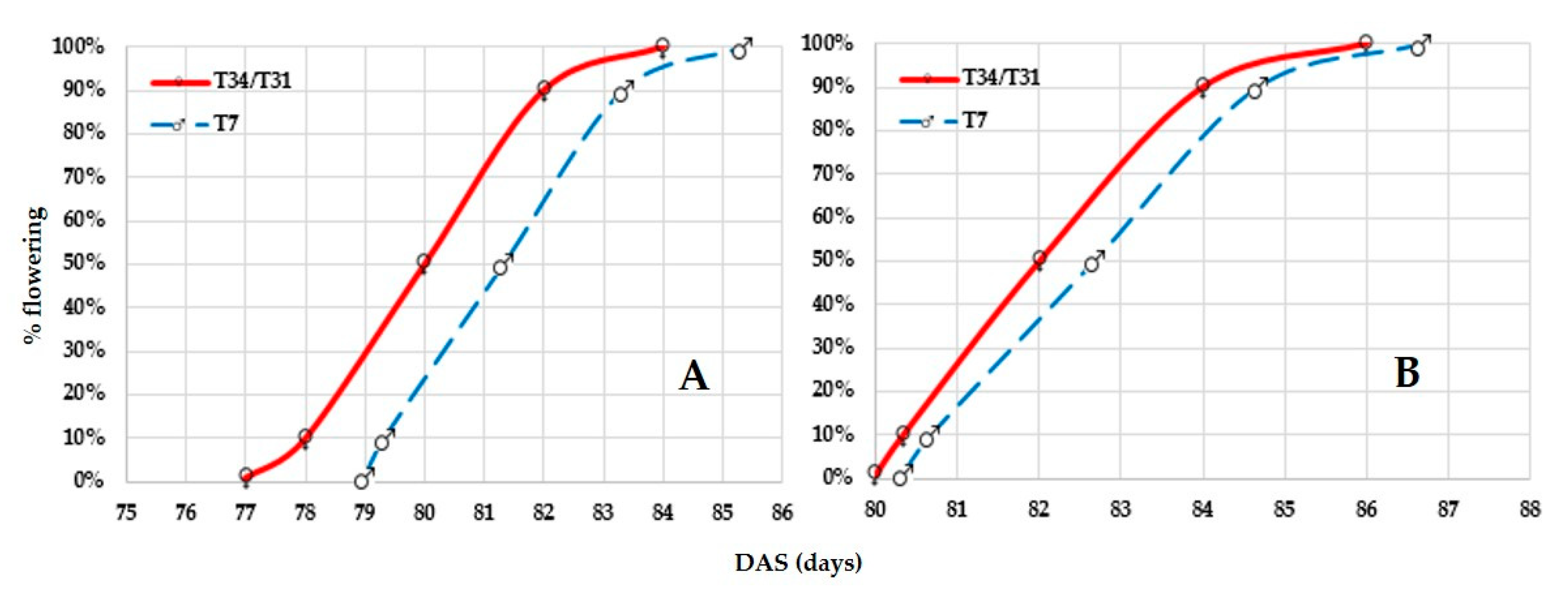
3.2.1. Floral Synchronization in Hybrid Seed Production
CLTHW13002 and CLTHW14003
CLTHW15001
CLTHY13002 and CLTHY15013
CLTHY15031
3.2.2. Effective Seed Yield (ESY) for FSC and Seed Yield (SY) for ML
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | DTA (days) | PP (days) | DT (cm) | SY (kg/ha) | |
|---|---|---|---|---|---|---|
| 0% | 90% | |||||
| ML | 4 | 24.23 ** 1.38 | 22.43 ** 1.18 | 1.07 ns | 1231 ns | 11,927,702.95 ** |
| CV (%) | 25.52 | 2.55 | 8.46 | |||
| Source | df | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|---|
| 0% | 90% | |||||
| Sowing dates (SD) | 2 | 203.25 ** | 118.11 ** | 11.86 ** | 10239.73 ** | 807,769.90 ns |
| FSC | 3 | 160.07 ** | 184.99 ** | 2.40 * | 598.10 ** | 14,291,233.74 ** |
| Plant densities (PD) | 2 | 2.58 ns | 20.53 ** | 8.69 ** | 305.01 ** | 2,868,422.79 ** |
| SD × FSC | 6 | 2.58 ns | 2.21 ns | 1.44 ns | 67.90 ns | 1,209,763.53 ** |
| PD × FSC | 6 | 0.77 ns | 0.44 ns | 1.57 ns | 73.77 ns | 1,037,679.49 * |
| SD × FSC × PD | 12 | 0.92 ns | 1.02 ns | 0.42 ns | 31.14 ns | 345,271.62 ns |
| CV (%) | 1.52 | 1.36 | 17.87 | 5.45 | 9.26 | |
| Source | df | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|---|
| 0% | 90% | |||||
| Sowing dates (SD) | 2 | 241.19 ** | 242.03 ** | 3.86 ns | 645.08 ** | 7,571,699.53 ** |
| FSC | 3 | 86.99 ** | 66.78 ** | 1.88 ns | 321.81 ** | 3,592,101.07 ** |
| SD × FSC | 6 | 1.49 ns | 1.36 ns | 0.27 ns | 43.56 ns | 439,581.49 ns |
| CV (%) | 1.53 | 1.00 | 23.1 | 5.55 | 7.68 | |
| DAS | 23 November 2017 | 16 December 2017 | 29 November 2018 | 5 December 2018 | ||||
|---|---|---|---|---|---|---|---|---|
| GDD day | GDD accu. | GDD day | GDD accu. | GDD day | GDD accu. | GDD day | GDD accu. | |
| 70 | 6.5 | 679.5 | 16.0 | 766.5 | 15.0 | 837.0 | 1.2 | 810.0 |
| 71 | 10.5 | 690.0 | 16.0 | 782.5 | 15.5 | 852.5 | 3.9 | 818.0 |
| 72 | 12.5 | 702.5 | 15.5 | 798.0 | 8.0 | 860.5 | 4.2 | 827.5 |
| 73 | 14.5 | 717.0 | 17.0 | 815.0 | 5.0 | 865.5 | 2.9 | 840.5 |
| 74 | 12.5 | 729.5 | 16.5 | 831.5 | 9.0 | 874.5 | 5.5 | 854.5 |
| 75 | 15.0 | 744.5 | 16.5 | 848.0 | 10.0 | 884.5 | 14.5 | 869.0 |
| 76 | 16.0 | 760.5 | 16.5 | 864.5 | 7.5 | 892.0 | 15.0 | 884.0 |
| 77 | 15.5 | 776.0 | 14.5 | 879.0 | 8.0 | 900.0 | 14.5 | 898.5 |
| 78 | 15.0 | 791.0 | 15.0 | 894.0 | 9.5 | 909.5 | 11.0 | 909.5 |
| 79 | 12.5 | 803.5 | 15.5 | 909.5 | 13.0 | 922.5 | 13.5 | 923.0 |
| 80 | 9.5 | 813.0 | 15.5 | 925.0 | 14.0 | 936.5 | 13.0 | 936.0 |
| 81 | 13.5 | 826.5 | 15.5 | 940.5 | 14.5 | 951.0 | 15.5 | 951.5 |
| 82 | 11.0 | 837.5 | 12.0 | 952.5 | 15.0 | 966.0 | 14.0 | 965.5 |
| 83 | 13.5 | 851.0 | 10.0 | 962.5 | 14.5 | 980.5 | 13.0 | 978.5 |
| 84 | 14.5 | 865.5 | 13.5 | 976.0 | 11.0 | 991.5 | 14.0 | 992.5 |
| 85 | 14.5 | 880.0 | 14.0 | 990.0 | 13.5 | 1005.0 | 15.0 | 1007.5 |
| 86 | 13.5 | 893.5 | 16.5 | 1006.5 | 13.0 | 1018.0 | 16.0 | 1023.5 |
| 87 | 15.5 | 909.0 | 11.5 | 1018.0 | 15.5 | 1033.5 | 15.0 | 1038.5 |
| 88 | 16.0 | 925.0 | 9.5 | 1027.5 | 14.0 | 1047.5 | 15.0 | 1053.5 |
| 89 | 15.0 | 940.0 | 14.0 | 1041.5 | 13.0 | 1060.5 | 15.5 | 1069.0 |
| 90 | 15.5 | 955.5 | 14.0 | 1055.5 | 14.0 | 1074.5 | 14.0 | 1083.0 |

References
- Prasanna, B.M. Maize in the developing world: Trends, challenges, and opportunities. Indonesian Center for Food Crops Research and Development (ICFORD), Indonesia. 2013. Available online: http://pangan.litbang.pertanian.go.id/files/IMC-PDF/03-Prasanna.pdf (accessed on 19 July 2020).
- Donnet, M.L.; López-Becerril, I.D.; Dominguez, C.; Arista-Cortés, J. Analysis of the sector structure and the public-private partnership of maize seed in Mexico. Agron. Mesoam. 2020, 31, 367–383. [Google Scholar] [CrossRef]
- Luna-Mena, B.; Hinojosa-Rodriguez, M.; Ayala-Garay, O.; Castillo-Gonzalez, F.; Mejía-Contreras, J. Perspectivas de desarrollo de la industria semillera en México. Rev. Fitotec. Mex. 2012, 35, 1–7. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0187-73802012000100003 (accessed on 19 July 2020).
- Córdova-Téllez, L.; Caballero-García, M.A.; Hernández-Nicolás, N.Y.; Ríos-Santos, E. Boletín Informativo de Producción de Semilla Calificada por el SNICS; SADER: Mexico City, Mexico, 2019; p. 96. [Google Scholar]
- Córdova-Téllez, L.; Caballero-García, M.A.; Hernández-Nicolás, N.Y.; Ríos-Santos, E. Boletín Informativo de Producción de Semilla Calificada por el SNICS. Ciclos agrícolas OI 2018–2019, PV 2019; SADER: Mexico City, Mexico, 2020; p. 89. [Google Scholar]
- Córdova-Téllez, L.; Caballero-García, M.A.; Hernández-Nicolás, N.Y.; Ríos-Santos, E.; Rebollar-Ávila, C. Boletín informativo de producción de semilla calificada por el SNICS. Ciclos agrícolas PV 2019, OI 2019–2020. Año 2, No. 1; SADER: Mexico City, Mexico, 2020; p. 64. [Google Scholar]
- Servicio de Información Agroalimentaria y Pesquera (SIAP). Estadística de Producción Agrícola de 2019. 2020. Available online: http://infosiap.siap.gob.mx/gobmx/datosAbiertos.php (accessed on 2 December 2020).
- San Vicente, F.; Leyva, M.; Gordon, R.; Deras, H.; Cruz, O.; Brenes, G.; Pineda, A.; Velázquez, R.; Martinez, H.; Gomez, M.; et al. 64 Reunión del Programa Cooperativo Centroamericano para el Mejoramiento de Cultivos y Animales (PCCMCA). Memoria Técnica Mesa de Maíz. IICA Editorial DICTA, Honduras. 21 pp. 2019. Available online: http://apps.iica.int/pccmca/docs/Memoria%20T%C3%A9cnica/Memoria%20T%C3%A9cnica%20-%20Mesa%20de%20Ma%C3%ADz.pdf (accessed on 2 December 2020).
- Skaugen, T.; Tveito, O. Growing-season and degree-day scenario in Norway for 2021–2050. Clim. Res. 2004, 26, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Iannucci, A.; Terribile, M.; Martiniello, P. Effects of temperature and photoperiod on flowering time of forage legumes in a Mediterranean environment. Field Crop. Res. 2008, 106, 156–162. [Google Scholar] [CrossRef]
- Hou, P.; Liu, Y.; Xie, R.; Ming, B.; Ma, D.; Li, S.; Mei, X. Temporal and spatial variation in accumulated temperature requirements of maize. Field Crop. Res. 2014, 158, 55–64. [Google Scholar] [CrossRef]
- Sacks, W.; Kucharik, C. Crop management and phenology trends in the U.S. Corn Belt: Impacts on yields, evapotranspiration and energy balance. Agric. For. Meteorol. 2011, 151, 882–894. [Google Scholar] [CrossRef]
- SNICS. Catálogo Nacional de Variedades Vegetales (online). 2020. Available online: https://datastudio.google.com/u/0/reporting/5b7206ba-e190-48fe-9696-73523bfccf58/page/itBWB (accessed on 2 December 2020).
- Ruiz, R.S.; Hernández, R.V. Producción de Semilla de Maíz Caso CIR Pacífico Centro; Folleto Técnico Núm. 4; Campo Experimental Centro-Altos de Jalisco: Tepatitlán de Morelos, Mexico, 2017; p. 61. [Google Scholar]
- Gilmore, E.C.; Rogers, J.S. Heat units as a method of measuring maturity in corn. Agron. J. 1958, 50, 611–615. [Google Scholar] [CrossRef]
- Noriega-González, L.; Preciado-Ortiz, R.; Andrio-Enríquez, E.; Terrón México-Ibarra1, A.; Covarrubias-Prieto, J. Phenology, plant growth and floral synchrony of the parental lines of H-374C QPM maize hybrid. Rev. Mex. Cienc. Agríc. 2011, 2, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Westgate, M.; Lisazo, J.; Batchelor, W. Quantitative relationship between pollen-shed density and grain yield in maize. Crop. Sci. 2003, 43, 934–942. [Google Scholar] [CrossRef]
- MacRobert, J.F.; Setimela, P.S.; Gethi, J.; Worku, M. Maize Hybrid. Seed Production Manual; International Maize and Wheat Improvement Center (CIMMYT): Mexico City, Mexico, 2014; pp. 1–26. [Google Scholar]
- Jiang, W.; Thapa, S.; Jessup, K.; Hao, B.; Hou, X.; Marek, T.; Becker, J.; Bell, J.; Xue, Q. Corn response to later than traditional planting dates in the Texas high plains. Crop. Sci. 2020, 60, 1004–1020. [Google Scholar] [CrossRef]
- Arellano-Vázquez, J.; Virgen-Vargas, J.; Martínez-Rojas, I. Gacela H72: Early maize hybrid to rainfed and irrigated areas of Central Highlands of Mexico. Rev. Mex. Cienc. Agríc. 2018, 9, 1303–1310. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, P.H.; Zaman-Allah, M.; Trachsel, S.; Seetharam, K.; Cairns, J.E.; Vinayan, M.T. Phenotyping for Abiotic Stress Tolerance in Maize Heat Stress: A Field Manual; CIMMYT: Mexico City, Mexico, 2016. [Google Scholar]
- Schlenker, W.; Roberts, M. Nonlinear temperature effects indicate severe damages to U.S. crop yields under climate change. Proc. Natl. Acad. Sci. USA 2009, 106, 15594–15598. [Google Scholar] [CrossRef] [Green Version]
- Cicchino, M.; Rattalino-Edreira, J.; Uribelarrea, M.; Otegui, M. Heat stress in field-grown maize: Response of physiological determinants of grain yield. Crop. Sci. 2010, 50, 1438–1448. [Google Scholar] [CrossRef]
- Sánchez, B.; Rasmussen, A.; Porter, J. Temperature and the growth and development of maize and rice: A review. Glob. Chang. Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef]
- Rodriguez, P. Aspectos climatológicos relacionados con la producción comercial de maíz. In El maíz en Venezuela; Fontana, H., González, C., Eds.; Fundación Polar: Caracas, Venezuela, 2000; pp. 61–67. [Google Scholar]
- Benacchio, S. Algunos Aspectos Agroecológicos Importantes en la Producción Maicera de Venezuela; FONAIAP: Caracas, Venezuela, 1983; p. 29. [Google Scholar]
- Peters, D.; Pendleton, J.; Hageman, R.; Brown, C. Effect of night air temperature on grain yield of corn, wheat, and soybeans. Agron. J. 1971, 63, 809. [Google Scholar] [CrossRef]
- Nelimor, C.; Badu-Apraku, B.; Tetteh, A.; Garcia-Oliveira, A.; N’guetta, A. Assessing the potential of extra-early maturing landraces for improving tolerance to drought, heat, and both combined stresses in maize. Agronomy 2020, 10, 318. [Google Scholar] [CrossRef] [Green Version]
- Major, D.; Brown, D.; Bootsma, A.; Dupuis, G.; Fairey, N.; Grant, E.; Green, D.; Hamilton, R.; Langille, J.; Sonmor, G.; et al. An evaluation of the corn heat unit system for the short-season growing regions across Canada. Can. J. Plant. Sci. 1983, 63, 121–130. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, R.; Hou, P.; Li, S.; Zhang, H.; Ming, B.; Long, H.; Liang, S. Phenological responses of maize to changes in environment when grown at different latitudes in China. Field Crop. Res. 2013, 144, 192–199. [Google Scholar] [CrossRef]
- Totis, L. Requerimientos agroclimáticos del cultivo de maíz. In: Bases para el manejo del cultivo de maíz. INTA. 2012. Available online: https://inta.gob.ar/sites/default/files/inta_bases_para_el_manejo_de_maiz_reglon_100-2_2.pdf (accessed on 23 July 2020).
- Wang, Y.; Tao, H.; Tian, B.; Sheng, D.; Xu, C.; Zhou, H.; Huang, S.; Wang, P. Flowering dynamics, pollen, and pistil contribution to grain yield in response to high temperature during maize flowering. Environ. Exp. Bot. 2019, 158, 80–88. [Google Scholar] [CrossRef]
- Baum, M.; Archontoulis, S.; Licht, M. Planting date, hybrid maturity, and weather effects on maize yield and crop stage. Agron. J. 2019, 3, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Kumudini, S.; Andrade, F.; Boote, K.; Brown, G.; Dzotsi, K.; Edmeades, G.; Gocken, T.; Goodwin, M.; Halter, A.; Hammer, G.; et al. Predicting maize phenology: Intercomparison of functions for developmental response to temperature. Agron. J. 2014, 106, 2087–2097. [Google Scholar] [CrossRef] [Green Version]
- Bolaños, J.; Edmeades, G. Eight cycles of selection for drought tolerance in lowland tropical maize. I. Responses in grain yield, biomass and radiation utilization. Field Crop. Res. 1993, 31, 233–252. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. Eight cycles of selection for drought tolerance in lowland tropical maize. II. Responses in reproductive behavior. Field Crop. Res. 1993, 31, 253–272. [Google Scholar] [CrossRef]
- Espinosa-Calderón, A.; Sierra-Macías, M.; Gómez-Montiel, N. Seed technology and production of improved maize varieties from INIFAP without PRONASE. Agron. Mesoam. 2003, 14, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Montiel, N.; González-Camarillo, M.; Cantú-Almaguer, M.; Sierra-Macías, M.; Coutiño-Estrada, B.; Manjarrez-Salgado, M. ‘H-563’, tropical maize hybrid tolerant to tar spot disease complex. Rev. Fitotec. Mex. 2013, 36, 81–83. [Google Scholar] [CrossRef]
- Virgen-Vargas, J.; Zepeda-Bautista, R.; Ávila-Perches, M.; Espinosa-Calderón, A.; Arellano-Vázquez, J.; Gámez-Vázquez, A. Seed production lines of maize: Population density and interaction. Agron. Mesoam. 2014, 25, 323–335. [Google Scholar] [CrossRef]
- Gómez-Montiel, N.; Cantú-Almaguer, M.; Vázquez-Carrillo, G.; Hernández-Galeno, C.; Espinosa-Calderón, A.; Sierra-Macías, M.; Coutiño-Estrada, B.; Aragón-Cuevas, F.; Trujillo-Campos, A. H-568 maize hybrid: New option for high productivity areas of the low tropic of Mexico. Rev. Mex. Cienc. Agríc. 2017, 8, 1213–1218. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Bravo, A.; Vidal-Martínez, V.; Bojórquez-Serrano, J.; García-Paredes, D. Respuesta del maíz al impacto ambiental ocurrido en las etapas de floración y ciclo vegetativo. Rev. Mex. Cienc. Agríc. 2014, 10, 2035–2045. [Google Scholar] [CrossRef] [Green Version]
- Arisnabarreta, S.; Solari, F. Hybrid maize seed production yield associations with inbred line performance in multienvironment trials. Crop. Sci. 2017, 57, 3203–3216. [Google Scholar] [CrossRef]
- Carcova, J.; Uribelarrea, M.; Borras, L.; Otegui, M.E.; Westgate, M.E. Synchronous pollination within and between ears improves kernel set in maize. Crop. Sci. 2000, 40, 1056–1061. [Google Scholar] [CrossRef]
- Sierra-Macías, M.; Palafox-Caballero, A.; Rodríguez-Montalvo, F.; Espinosa-Calderón, A.; Gómez-Montiel, N.; Caballero-Hernández, F.; Barrón-Freyre, S.; Zambada-Martínez, A.; Vázquez-Carrillo, G. H-520, three way maize hybrid for the humid tropic of Mexico. Agric. Téc. Méx. 2008, 34, 119–122. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0568-25172008000100015&lng=es&nrm=iso (accessed on 4 December 2020).
- Chassaigne-Ricciulli, A.A.; Mendoza-Onofre, L.E.; Córdova-Téllez, L.; Carballo-Carballo, A.; San Vicente-García, F.M.; Dhliwayo, T. Development of seed production technology of CIMMYT tropical single cross maize hybrids. Agriculture 2020, 10, 259. [Google Scholar] [CrossRef]
- McCoy, A.; Roth, M.; Shay, R.; Noel, Z.; Jayawardana, M.; Longley, R.; Bonito, G.; Chilvers, M. Identification of fungal communities within the tar spot complex of corn in Michigan via next-generation sequencing. Phytobiomes. J. 2019, 3, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]






| Female Single Crosses | Male Inbred Line | Pedigree | TWH | Year of Release |
|---|---|---|---|---|
| T13/T14 | T10 | T13/T14//T10 | CLTHW13002 | 2013 |
| T4/T6 | T10 | T4/T6//T10 | CLTHW14003 | 2014 |
| T32/T10 | T27 | T32/T10//T27 | CLTHW15001 | 2016 |
| T7/T1 | T11 | T7/T1//T11 | CLTHY13002 | 2013 |
| T7/T1 | T31 | T7/T1//T31 | CLTHY15013 | 2016 |
| T34/T31 | T7 | T34/T31//T7 | CLTHY15031 | 2016 |
| Parents | Group 1 | Group 2 | Group 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2018A | 2019A | 2018A | 2019A | 2018A | 2019A | ||||
| Exp. 1 | Exp. 2 | Exp. 3 | Exp. 4 | Exp. 5 | Exp. 6 | Exp. 7 | Exp. 8 | Exp. 9 | |
| SD1 | SD3 | SD4 | SD2 | SD3 | SD4 | SD1 | SD3 | SD4 | |
| T7 | X | X | X | ||||||
| T10 | X | ||||||||
| T11 | X | X | X | ||||||
| T27 | X | X | X | ||||||
| T31 | X | X | X | ||||||
| T13/T14 | X | X | X | X | X | X | |||
| T32/T10 | X | X | X | X | X | X | |||
| T4/T6 | X | X | X | X | X | X | |||
| T7/T1 | X | X | X | X | X | X | |||
| T34/T31 | X | X | X | X | |||||
| Source | df | DTA (days) | PP (days) | DT (cm) | SY (kg/ha) | |||
|---|---|---|---|---|---|---|---|---|
| 0% | 10% | 50% | 90% | |||||
| SD | 2 | 173.58 ** | 214.08 ** | 205.44 ** | 213.53 ** | 2.09 ns | 320.58 ** | 2,698,813.36 ** |
| ML | 3 | 45.73 ** | 47.33 ** | 53.29 ** | 45.29 ** | 1.76 ns | 2944.89 ** | 8,199,353.07 ** |
| SD × ML | 6 | 2.29 ns | 2.31 ns | 2.93 ns | 3.01 ns | 0.60 ns | 260.25 ** | 1,263,451.10 ** |
| CV (%) | 1.45 | 1.42 | 1.45 | 1.35 | 19.07 | 3.28 | 7.54 | |
| Concept and (n) | DTA (days) | PP (days) | DT (cm) | SY (kg/ha) | |||
|---|---|---|---|---|---|---|---|
| 0% | 10% | 50% | 90% | ||||
| Sowing Dates | |||||||
| SD1: Exp. 1 (12) | 88 a | 89 a | 91 a | 93 a | 4.0 | 214 a | 4704 b |
| SD3: Exp. 2 (12) | 81 c | 81 c | 83 c | 85 c | 4.4 | 204 b | 5605 a |
| SD4: Exp. 3 (12) | 83 b | 83 b | 85 b | 87 b | 4.8 | 209 ab | 4899 b |
| Male lines | |||||||
| T31 (9) | 86 a | 87 a | 89 a | 90 a | 4.3 | 201 b | 6193 a |
| T27 (9) | 85 a | 86 a | 88 a | 90 a | 4.4 | 236 a | 5282 b |
| T11 (9) | 82 b | 83 b | 84 b | 86 b | 3.9 | 199 b | 4918 b |
| T7 (9) | 82 b | 82 b | 84 b | 86 b | 5.0 | 199 b | 3885 c |
| ML and (n) | DTA (days) | PP (days) | DT (cm) | SY (kg/ha) | |
|---|---|---|---|---|---|
| 0% | 90% | ||||
| T27 (3) | 90 a | 93 a | 4.7 | 229 | 4303 c |
| T31 (3) | 90 a | 93 a | 4.7 | 216 | 6786 b |
| T11 (3) | 87 ab | 89 b | 4.0 | 208 | 4672 c |
| T7 (3) | 85 bc | 88 bc | 6.0 | 203 | 3057 d |
| T10 (3) | 83 c | 86 c | 4.7 | 204 | 8001 a |
| Source | df | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|---|
| 0% | 90% | |||||
| Sowing dates (SD) | 1 | 435.60 ** | 273.88 ** | 18.68 ** | 448.90 ** | 67,623.21 ns |
| FSC | 4 | 118.46 ** | 111.04 ** | 0.88 ns | 3887.54 ** | 9,741,608.46 ** |
| Plant densities (PD) | 2 | 5.28 * | 25.43 ** | 8.04 ** | 698.63 ** | 4,443,690.0 ** |
| SD × FSC | 4 | 4.77 * | 1.35 ns | 1.84 * | 26.37 ns | 1,330,460.24 ** |
| PD × FSC | 8 | 1.10 ns | 0.99 ns | 0.67 ns | 67.19 ns | 860,668.61 ** |
| SD × PD × FSC | 8 | 0.73 ns | 0.27 ns | 0.40 ns | 24.12 ns | 395,147.11 ns |
| CV (%) | 1.68 | 1.25 | 16.93 | 6.45 | 8.08 | |
| Source | df | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|---|
| 0% | 90% | |||||
| Sowing dates (SD) | 1 | 70.53 ** | 30.00 ** | 8.53 ** | 24.30 ns | 76,104.03 ns |
| FSC | 4 | 53.67 ** | 41.92 ** | 1.25 ns | 1402.88 ** | 1,962,139.80 ** |
| SD × FSC | 4 | 0.37 ns | 0.58 ns | 0.80 ns | 27.05 ns | 430,160.37 ns |
| CV (%) | 1.41 | 1.02 | 21.27 | 6.69 | 7.96 | |
| Concept and (n) | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|
| 0% | 90% | ||||
| Sowing dates | |||||
| SD 3: Exp. 5 (45) | 74 b | 79 b | 5.3 a | 125 a | 6533 |
| SD 4: Exp. 6 (45) | 78 a | 83 a | 4.4 b | 121 b | 6478 |
| Planting densities | |||||
| 82,667 pt/ha (30) | 76 b | 80 c | 4.4 b | 125 a | 6687 a |
| 93,333 pt/ha (30) | 76 ab | 81 b | 4.7 b | 126 a | 6767 a |
| 101,333 pt/ha (30) | 76 a | 82 a | 5.4 a | 117 b | 6064 b |
| Female single crosses | |||||
| T4/T6 (18) | 78 a | 83 a | 4.9 | 124 b | 6103 b |
| T7/T1 (18) | 77 a | 82 a | 5.0 | 136 a | 6094 b |
| T34/T31 (18) | 78 a | 83 a | 4.6 | 98 c | 5761 b |
| T13/T14 (18) | 74 b | 78 b | 4.7 | 127 b | 7103 a |
| T32/T10 (18) | 73 b | 78 b | 5.1 | 130 ab | 7466 a |
| Concept and (n) | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|
| 0% | 90% | ||||
| Sowing dates | |||||
| SD3: Exp. 8 (15) | 74 b | 80 b | 5.2 a | 123 | 6274 |
| SD4: Exp. 9 (15) | 78 a | 82 a | 4.1 b | 122 | 6374 |
| Female single crosses | |||||
| T4/T6 (6) | 78 a | 83 a | 4.7 | 128 ab | 6228 abc |
| T7/T1 (6) | 78 a | 82 a | 4.0 | 137 a | 6012 bc |
| T34/T31 (6) | 79 a | 83 a | 4.5 | 98 c | 5591 c |
| T13/T14 (6) | 74 b | 79 b | 5.0 | 119 b | 6820 ab |
| T32/T10 (6) | 72 c | 77 c | 5.2 | 131 ab | 6970 a |
| Concept and (n) | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|
| 0% | 90% | ||||
| Sowing dates | |||||
| SD2: Exp. 4 (36) | 75 b | 80 b | 4.8 b | 158 a | 6433 |
| SD3: Exp. 5 (36) | 73 c | 79 c | 5.5 a | 131 b | 6680 |
| SD4: Exp. 6 (36) | 78 a | 82 a | 4.4 b | 127 b | 6703 |
| Planting densities | |||||
| 82,667 pt/ha (30) | 75 | 80 b | 4.4 b | 140 a | 6826 a |
| 93,333 pt/ha (30) | 76 | 81 a | 5.0 a | 141 a | 6703 a |
| 101,333 pt/ha (30) | 76 | 81 a | 5.3 a | 136 b | 6287 b |
| Female single crosses | |||||
| T4/T6 (27) | 78 a | 83 a | 5.0 ab | 135 b | 5935 b |
| T7/T1 (27) | 77 a | 82 a | 5.1 a | 145 a | 6019 b |
| T13/T14 (27) | 74 b | 78 b | 4.4 b | 135 b | 7188 a |
| T32/T10 (27) | 73 b | 78 b | 5.0 ab | 141 a | 7280 a |
| Concept and (n) | DTS (days) | SP (days) | DS (cm) | ESY (kg/ha) | |
|---|---|---|---|---|---|
| 0% | 90% | ||||
| Sowing dates | |||||
| SD1: Exp. 7 (12) | 83 a | 88 a | 5.3 | 141 a | 7864 a |
| SD3: Exp. 5 (12) | 74 c | 79 c | 4.2 | 129 b | 6376 b |
| SD4: Exp. 6 (12) | 77 b | 81 b | 5.0 | 128 b | 6639 b |
| Female single crosses | |||||
| T4/T6 (9) | 81 a | 86 a | 4.7 | 131 ab | 6511 b |
| T7/T1 (9) | 80 a | 84 b | 4.2 | 140 a | 6326 b |
| T13/T14 (9) | 76 b | 81 c | 5.1 | 126 b | 7435 a |
| T32/T10 (9) | 75 b | 80 d | 5.2 | 134 ab | 7567 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chassaigne-Ricciulli, A.A.; Mendoza-Onofre, L.E.; Córdova-Téllez, L.; Carballo-Carballo, A.; San Vicente-García, F.M.; Dhliwayo, T. Effective Seed Yield and Flowering Synchrony of Parents of CIMMYT Three-Way-Cross Tropical Maize Hybrids. Agriculture 2021, 11, 161. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020161
Chassaigne-Ricciulli AA, Mendoza-Onofre LE, Córdova-Téllez L, Carballo-Carballo A, San Vicente-García FM, Dhliwayo T. Effective Seed Yield and Flowering Synchrony of Parents of CIMMYT Three-Way-Cross Tropical Maize Hybrids. Agriculture. 2021; 11(2):161. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020161
Chicago/Turabian StyleChassaigne-Ricciulli, Alberto A., Leopoldo E. Mendoza-Onofre, Leobigildo Córdova-Téllez, Aquiles Carballo-Carballo, Félix M. San Vicente-García, and Thanda Dhliwayo. 2021. "Effective Seed Yield and Flowering Synchrony of Parents of CIMMYT Three-Way-Cross Tropical Maize Hybrids" Agriculture 11, no. 2: 161. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020161