
Temporal Changes in Sensitivity of Zymoseptoria tritici Field Populations to Different Fungicidal Modes of Action


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Area and Isolation of Z. tritici Field Populations
2.2. Fungicidal Active Ingredients
2.3. In Vitro Fungicide Sensitivity Testing
2.4. Data Analysis
2.5. Statistical Analysis
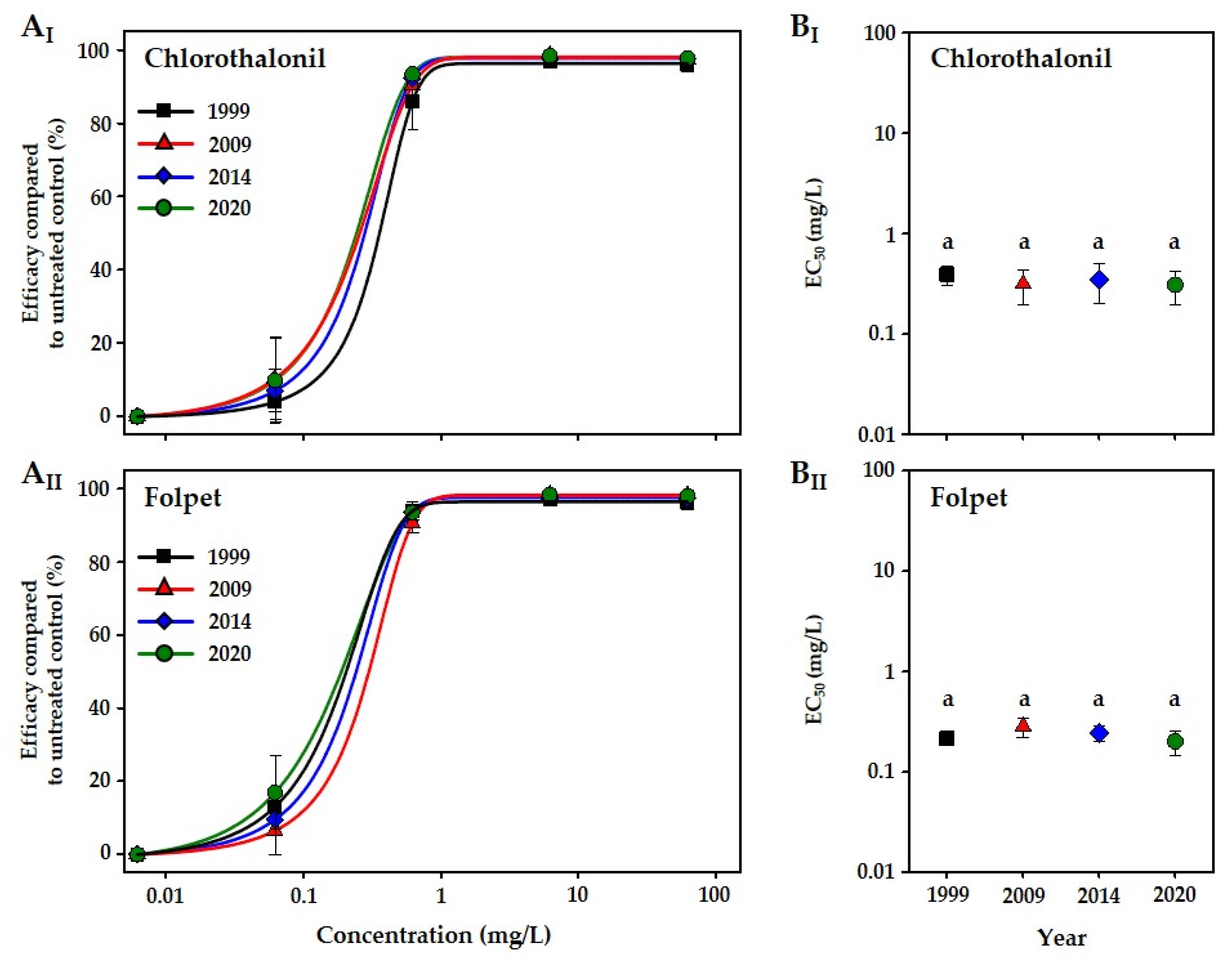
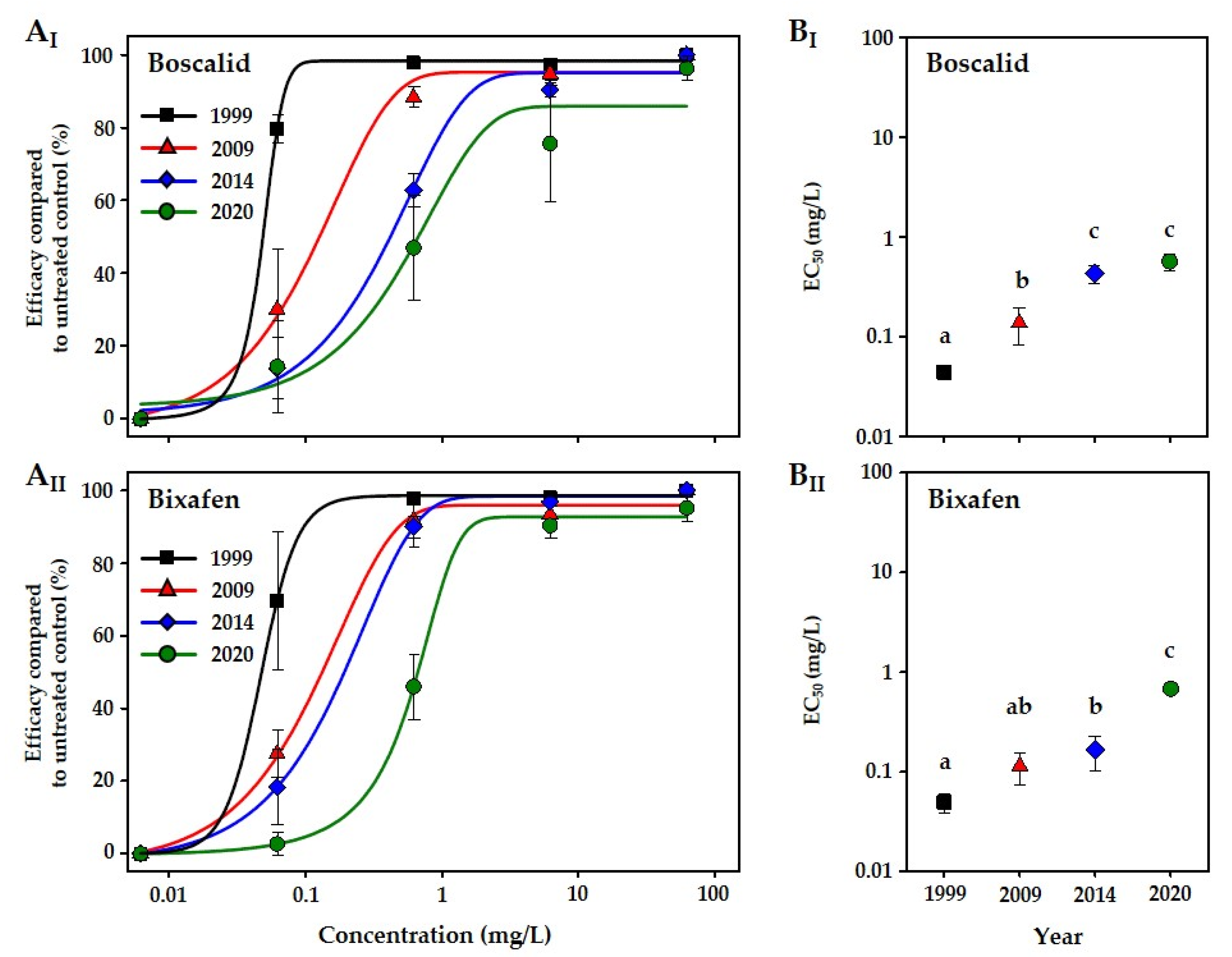
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Driscoll, A.; Kildea, S.; Doohan, F.; Spink, J.; Mullins, E. The wheat-Septoria conflict: A new front opening up? Trends Plant Sci. 2014, 19, 602–610. [Google Scholar] [CrossRef]
- Jørgensen, L.N.; Hovmøller, M.S.; Hansen, J.G.; Lassen, P.; Clark, B.; Bayles, R.; Rodemann, B.; Flath, K.; Jahn, M.; Goral, T.; et al. IPM strategies and their dilemmas including an introduction to www.eurowheat.org. J. Integr. Agric. 2014, 13, 265–281. [Google Scholar] [CrossRef]
- Fones, H.; Gurr, S. The impact of Septoria tritici blotch disease on wheat: An EU perspective. Fungal Genet. Biol. 2015, 79, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torriani, S.F.F.; Melichar, J.P.E.; Mills, C.; Pain, N.; Sierotzki, H.; Courbot, M. Zymoseptoria tritici: A major threat to wheat production, integrated approaches to control. Fungal Genet. Biol. 2015, 79, 8–12. [Google Scholar] [CrossRef]
- Schuh, W. Influence of tillage systems on disease intensity and spatial pattern of Septoria leaf blotch. Phytopathology 1990, 80, 1337. [Google Scholar] [CrossRef] [Green Version]
- Gladders, P.; Paveley, N.D.; Barrie, I.A.; Hardwick, N.V.; Hims, M.J.; Langton, S.; Taylor, M.C. Agronomic and meteorological factors affecting the severity of leaf blotch caused by Mycosphaerella graminicola in commercial wheat crops in England. Ann. Appl. Biol. 2001, 138, 301–311. [Google Scholar] [CrossRef]
- McDonald, B.A.; Mundt, C.C. How knowledge of pathogen population biology informs management of Septoria tritici blotch. Phytopathology 2016, 106, 948–955. [Google Scholar] [CrossRef] [Green Version]
- Bankina, B.; Bimšteine, G.; Arhipova, I.; Kaņeps, J.; Stanka, T. Importance of agronomic practice on the control of wheat leaf diseases. Agriculture 2018, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Lucas, J.A.; Fraaije, B.A. QoI resistance in Mycosphaerella graminicola: What have we learned so far? In Modern Fungicides and Antifungal Compounds. In Proceedings of the 15th International Reinhardsbrunn Symposium, Friedrichroda, Germany, 6–10 May 2007; Dehne, H.W., Deising, H.B., Gisi, U., Kuck, K.H., Russel, P.E., Lyr, H., Eds.; Deutsche Phytomedizinische Gesellschaft: Braunschweig, Germany, 2008; Volume V. [Google Scholar]
- Gisi, U.; Sierotzki, H.; Cook, A.; McCaffery, A. Mechanisms influencing the evolution of resistance to Qo inhibitor fungicides. Pest Manag. Sci. 2002, 58, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Fraaije, B.A.; Cools, H.J.; Fountaine, J.; Lovell, D.J.; Motteram, J.; West, J.S.; Lucas, J.A. Role of ascospores in further spread of QoI-resistant cytochrome b alleles (G143A) in field populations of Mycosphaerella graminicola. Phytopathology 2005, 95, 933–941. [Google Scholar] [CrossRef] [Green Version]
- Fraaije, B.A.; Cools, H.J.; Kim, S.-H.; Motteram, J.; Clark, W.S.; Lucas, J.A. A novel substitution I381V in the sterol 14alpha-demethylase (CYP51) of Mycosphaerella graminicola is differentially selected by azole fungicides. Mol. Plant Pathol. 2007, 8, 245–254. [Google Scholar] [CrossRef]
- Torriani, S.F.; Brunner, P.C.; McDonald, B.A.; Sierotzki, H. QoI resistance emerged independently at least 4 times in European populations of Mycosphaerella graminicola. Pest Manag. Sci. 2009, 65, 155–162. [Google Scholar] [CrossRef]
- Cheval, P.; Siah, A.; Bomble, M.; Popper, A.D.; Reignault, P.; Halama, P. Evolution of QoI resistance of the wheat pathogen Zymoseptoria tritici in Northern France. Crop Prot. 2017, 92, 131–133. [Google Scholar] [CrossRef]
- Blake, J.J.; Gosling, P.; Fraaije, B.A.; Burnett, F.J.; Knight, S.M.; Kildea, S.; Paveley, N.D. Changes in field dose-response curves for demethylation inhibitor (DMI) and quinone outside inhibitor (QoI) fungicides against Zymoseptoria tritici, related to laboratory sensitivity phenotyping and genotyping assays. Pest Manag. Sci. 2018, 74, 302–313. [Google Scholar] [CrossRef]
- Jørgensen, L.N.; Matzen, N.; Heick, T.M.; Havis, N.; Holdgate, S.; Clark, B.; Blake, J.; Glazek, M.; Korbas, M.; Danielewicz, J.; et al. Decreasing azole sensitivity of Z. tritici in Europe contributes to reduced and varying field efficacy. J. Plant Dis. Prot. 2020. [CrossRef]
- Curvers, K.; Pycke, B.; Kyndt, T.; Haesaert, G.; Gheysen, G. Sensitivity towards DMI fungicides and haplotypic diversity of their CYP51 target in the Mycosphaerella graminicola population of Flanders. J. Plant Dis. Prot. 2014, 121, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Stammler, G.; Semar, M. Sensitivity of Mycosphaerella graminicola (anamorph: Septoria tritici) to DMI fungicides across Europe and impact on field performance. EPPO Bull. 2011, 41, 149–155. [Google Scholar] [CrossRef]
- Kildea, S.; Dooley, H.; Phelan, S.; Mehenni-Ciz, J.; Spink, J. Developing fungicide control programmes for blotch in Irish winter wheat crops. In Modern Fungicides and Antifungal Compounds, Proceedings of the 18th International Reinhardsbrunn Symposium, Friedrichroda, Germany, 24–28 April 2016; Deising, H.B., Fraaije, B., Mehl, A., Oerke, E.C., Sierotzki, H., Stammler, G., Eds.; Deutsche Phytomedizinische Gesellschaft: Braunschweig, Germany, 2017; Volume VIII. [Google Scholar]
- Strobel, D.; Bryson, R.; Roth, J.; Stammler, G. Field performance of DMI fungicides against Zymoseptoria tritici across Europe—Compromized by further sensitivity shift? In Modern Fungicides and Antifungal Compounds, Proceedings of the 18th International Reinhardsbrunn Symposium, Friedrichroda, Germany, 24–28 April 2016; Deising, H.B., Fraaije, B., Mehl, A., Oerke, E.C., Sierotzki, H., Stammler, G., Eds.; Deutsche Phytomedizinische Gesellschaft: Braunschweig, Germany, 2017; Volume VIII. [Google Scholar]
- Kildea, S.; Heick, T.M.; Grant, J.; Mehenni-Ciz, J.; Dooley, H. A combination of target-site alterations, overexpression and enhanced efflux activity contribute to reduced azole sensitivity present in the Irish Zymoseptoria tritici population. Eur. J. Plant Pathol. 2019, 154, 529–540. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; van Nistelrooy, J.G.M.; Kema, G.H.J.; de Waard, M.A. Multiple mechanisms account for variation in base-line sensitivity to azole fungicides in field isolates of Mycosphaerella graminicola. Pest Manag. Sci. 2003, 59, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Leroux, P.; Albertini, C.; Gautier, A.; Gredt, M.; Walker, A.S. Mutations in the CYP51 gene correlated with changes in sensitivity to sterol 14 alpha-demethylation inhibitors in field isolates of Mycosphaerella graminicola. Pest Manag. Sci. 2007, 63. [Google Scholar] [CrossRef]
- Leroux, P.; Walker, A.S. Multiple mechanisms account for resistance to sterol 14α-demethylation inhibitors in field isolates of Mycosphaerella graminicola. Pest Manag. Sci. 2011, 67. [Google Scholar] [CrossRef]
- Cools, H.J.; Bayon, C.; Atkins, S.; Lucas, J.A.; Fraaije, B.A. Overexpression of the sterol 14α-demethylase gene (MgCYP51) in Mycosphaerella graminicola isolates confers a novel azole fungicide sensitivity phenotype. Pest Manag. Sci. 2012, 68, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Cools, H.J.; Fraaije, B.A. Update on mechanisms of azole resistance in Mycosphaerella graminicola and implications for future control. Pest Manag. Sci. 2013, 69. [Google Scholar] [CrossRef]
- Omrane, S.; Sghyer, H.; Audéon, C.; Lanen, C.; Duplaix, C.; Walker, A.S.; Fillinger, S. Fungicide efflux and the MgMFS1 transporter contribute to the multidrug resistance phenotype in Zymoseptoria tritici field isolates. Environ. Microbiol. 2015, 17. [Google Scholar] [CrossRef]
- Rehfus, A.; Strobel, D.; Bryson, R.; Stammler, G. Mutations in sdh genes in field isolates of Zymoseptoria tritici and impact on the sensitivity to various succinate dehydrogenase inhibitors. Plant Pathol. 2018, 67, 175–180. [Google Scholar] [CrossRef]
- Yamashita, M.; Fraaije, B. Non-target site SDHI resistance is present as standing genetic variation in field populations of Zymoseptoria tritici. Pest Manag. Sci. 2018, 74, 672–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierotzki, H.; Scalliet, G. A review of current knowledge of resistance aspects for the next-generation succinate dehydrogenase inhibitor fungicides. Phytopathology 2013, 103, 880–887. [Google Scholar] [CrossRef] [Green Version]
- Dooley, H.; Shaw, M.W.; Mehenni-Ciz, J.; Spink, J.; Kildea, S. Detection of Zymoseptoria tritici SDHI-insensitive field isolates carrying the SdhC-H152R and SdhD-R47W substitutions. Pest Manag. Sci. 2016, 72, 2203–2207. [Google Scholar] [CrossRef] [PubMed]
- Kirikyali, N.; Diez, P.; Hawkins, N.; Fraaije, B.A. Azole and SDHI sensitivity status of Zymoseptoria tritici field populations sampled in France, Germany and the UK during 2015. In Modern Fungicides and Antifungal Compounds, Proceedings of the 18th International Reinhardsbrunn Symposium, Friedrichroda, Germany, 24–28 April 2016; Deising, H.B., Fraaije, B., Mehl, A., Oerke, E.C., Sierotzki, H., Stammler, G., Eds.; Deutsche Phytomedizinische Gesellschaft: Braunschweig, Germany, 2017; Volume VIII. [Google Scholar]
- Hobbelen, P.H.F.; Paveley, N.D.; van den Bosch, F. Delaying selection for fungicide insensitivity by mixing fungicides at a low and high risk of resistance development: A modeling analysis. Phytopathology 2011, 101, 1224–1233. [Google Scholar] [CrossRef]
- Oliver, R.P.; Hewitt, H. Fungicides in Crop Protection; CABI: Wallingford, UK, 2014. [Google Scholar]
- Heick, T.M.; Justesen, A.F.; Jørgensen, L.N. Resistance of wheat pathogen Zymoseptoria tritici to DMI and QoI fungicides in the Nordic-Baltic region—A status. Eur. J. Plant Pathol. 2017, 149. [Google Scholar] [CrossRef]
- Deutscher Wetterdienst (DWD). Klimareport Schleswig-Holstein. Available online: https://www.dwd.de/DE/leistungen/klimareport_sh/download_report_2017.pdf?__blob=publicationFile&v=5 (accessed on 4 February 2021).
- Verreet, J.A.; Klink, H.; Hoffmann, G.M. Regional monitoring for disease prediction and optimization of plant protection measuares: The IPM Wheat Model. Plant Dis. 2000, 84, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henze, M.; Beyer, M.; Klink, H.; Verreet, J.-A. Characterizing Meteorological Scenarios Favorable for Septoria tritici Infections in Wheat and Estimation of Latent Periods. Plant Dis. 2007, 91, 1445–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statistisches Amt für Hamburg und Schleswig-Holstein. Die Bodennutzung in Schleswig-Holstein 2017. Available online: http://epub.sub.uni-hamburg.de/epub/volltexte/2018/78253/pdf/C_I_1_j_17_SH_e.pdf (accessed on 4 February 2021).
- Statistisches Amt für Hamburg und Schleswig-Holstein. Getreideernte in Schleswig-Holstein 2020. Available online: https://www.statistik-nord.de/fileadmin/Dokumente/Presseinformationen/SI20_116.pdf (accessed on 4 February 2021).
- Beyer, M.; Kiesner, F.; Verreet, J.-A.; Klink, H. Fungicide sensitivity of Septoria tritici field isolates is affected by an interaction between fungicidal mode of action and time. J. Plant Pathol. 2011, 93, 7–13. [Google Scholar]
- Dachbrodt-Saaydeh, S.; Sellmann, J.; Strassemeyer, J.; Schwarz, J.; Klocke, B.; Krengel, S.; Kehlenbeck, H. Netz Vergleichsbetriebe Pflanzenschutz Zwei-Jahresbericht 2015 und 2016—Analyse der Ergebnisse der Jahre 2007 bis 2016. Berichte aus dem Julius Kühn-Institut 2018, 194. [Google Scholar] [CrossRef]
- Bundessortenamt. Beschreibende Sortenliste 2008. Available online: https://www.bundessortenamt.de/bsa/media/Files/BSL/bsl_getreide_2008.pdf (accessed on 10 September 2020).
- Birr, T.; Verreet, J.-A.; Klink, H. Prediction of deoxynivalenol and zearalenone in winter wheat grain in a maize-free crop rotation based on cultivar susceptibility and meteorological factors. J. Plant Dis. Prot. 2019, 126, 13–27. [Google Scholar] [CrossRef]
- Birr, T.; Hasler, M.; Verreet, J.-A.; Klink, H. Composition and predominance of Fusarium species causing Fusarium head blight in winter wheat grain depending on cultivar susceptibility and meteorological factors. Microorganisms 2020, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, Austria). Available online: https://www.R-project.org/ (accessed on 22 June 2020).
- Laird, N.M.; Ware, J.H. Random-effects models for longitudinal data. Biometrics 1982, 38, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Bretz, F.; Hothorn, T.; Westfall, P.H. Multiple Comparisons Using R; Chapman & Hall/CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Russell, P.E. A century of fungicide evolution. J. Agric. Sci. 2005, 143, 11–25. [Google Scholar] [CrossRef]
- Van den Bosch, F.; Paveley, N.; van den Berg, F.; Hobbelen, P.; Oliver, R. Mixtures as a fungicide resistance management tactic. Phytopathology 2014, 104, 1264–1273. [Google Scholar] [CrossRef]
- Fraaije, B.A.; Bayon, C.; Atkins, S.; Cools, H.J.; Lucas, J.A.; Fraaije, M.W. Risk assessment studies on succinate dehydrogenase inhibitors, the new weapons in the battle to control Septoria leaf blotch in wheat. Mol. Plant Pathol. 2012, 13, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.J.; Hims, M.J.; Vaughan, T.B. Effects of fungicide spray timing on winter wheat disease control. Plant Pathol. 1999, 48, 33–50. [Google Scholar] [CrossRef]
- Van den Bosch, F.; Oliver, R.; van den Berg, F.; Paveley, N. Governing principles can guide fungicide-resistance management tactics. Annu. Rev. Phytopathol. 2014, 52, 175–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commision. Commission Implementing Regulation (EU) 2019/677 of 29 April 2019 concerning the non-renewal of the approval of the active substance chlorothalonil, in accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council concerning the placing of plant protection products on the market, and amending Commission Implementing Regulation (EU) No 40/2011. Off. J. Eur. Union 2019, L114, 15–17. [Google Scholar]
- European Commision. Commission Implementing Regulation (EU) 2020/2087 of 14 December 2020 concerning the non-renewal of the approval of the active substance mancozeb, in accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council concerning the placing of plant protection products on the market, and amending the Annex to Commission Implementing Regulation (EU) No 540/2011. Off. J. Eur. Union 2020, L423, 50–52. [Google Scholar]
- Mäe, A.; Fillinger, S.; Sooväli, P.; Heick, T.M. Fungicide sensitivity shifting of Zymoseptoria tritici in the Finnish-Baltic region and a novel insertion in the MFS1 promoter. Front. Plant Sci. 2020, 11, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huf, A.; Rehfus, A.; Lorenz, K.H.; Bryson, R.; Voegele, R.T.; Stammler, G. Proposal for a new nomenclature for CYP51 haplotypes in Zymoseptoria tritici and analysis of their distribution in Europe. Plant Pathol. 2018, 67, 1706–1712. [Google Scholar] [CrossRef]
- Dooley, H.; Shaw, M.W.; Spink, J.; Kildea, S. Effect of azole fungicide mixtures, alternations and dose on azole sensitivity in the wheat pathogen Zymoseptoria tritici. Plant Pathol. 2016, 65, 124–136. [Google Scholar] [CrossRef]
- European Commission. Commission Implementing Regulation (EU) 2018/1865 of 28 November 2018 concerning the non-renewal of approval of the active substance propiconazole, in accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council concerning the placing of plant protection products on the market, and amending Commission Implementing Regulation (EU) No 540/2011. Off. J. Eur. Union 2018, L304, 6–9. [Google Scholar]
- European Commission. Commission Implementing Regulation (EU) 2019/168 of 31 January 2019 amending Implementing Regulation (EU) No 540/2011 as regards the extension of the approval periods of the active substances abamectin, Bacillus subtilis (Cohn 1872) Strain QST 713, Bacillus thuringiensis subsp. Aizawai, Bacillus thuringiensis subsp. israeliensis, Bacillus thuringiensis subsp. kurstaki, Beauveria bassiana, benfluralin, clodinafop, clopyralid, Cydia pomonella Granulovirus (CpGV), cyprodinil, dichlorprop-P, epoxiconazole, fenpyroximate, fluazinam, flutolanil, fosetyl, Lecanicillium muscarium, mepanipyrim, mepiquat, Metarhizium anisopliae var. Anisopliae, metconazole, metrafenone, Phlebiopsis gigantea, pirimicarb, Pseudomonas chlororaphis strain: MA 342, pyrimethanil, Pythium oligandrum, rimsulfuron, spinosad, Streptomyces K61, thiacloprid, tolclofos-methyl, Trichoderma asperellum, Trichoderma atroviride, Trichoderma gamsii, Trichoderma harzianum, triclopyr, trinexapac, triticonazole, Verticillium albo-atrum and ziram. Off. J. Eur. Union 2019, L33, 1–4. [Google Scholar]
- Sykes, E.M.; Sackett, K.E.; Severns, P.M.; Mundt, C.C. Sensitivity variation and cross-resistance of Zymoseptoria tritici to azole fungicides in North America. Eur. J. Plant Pathol. 2017. [Google Scholar] [CrossRef]
- Schmeling, B.V.; Kulka, M. Systemic fungicidal activity of 1,4-oxathiin derivatives. Science 1966, 152, 659–660. [Google Scholar] [CrossRef]
- Avenot, H.F.; Michailides, T.J. Progress in understanding molecular mechanisms and evolution of resistance to succinate dehydrogenase inhibiting (SDHI) fungicides in phytopathogenic fungi. Crop Prot. 2010, 29, 643–651. [Google Scholar] [CrossRef]
- Rehfus, A.; Miessner, S.; Achenbach, J.; Strobel, D.; Bryson, R.; Stammler, G. Emergence of succinate dehydrogenase inhibitor resistance of Pyrenophora teres in Europe. Pest Manag. Sci. 2016, 72. [Google Scholar] [CrossRef]
- Skinner, W.; Bailey, A.; Renwick, A.; Keon, J.; Gurr, S.; Hargreaves, J. A single amino-acid substitution in the iron-sulphur protein subunit of succinate dehydrogenase determines resistance to carboxin in Mycosphaerella graminicola. Curr. Genet. 1998, 34, 393–398. [Google Scholar] [CrossRef]
- Stammler, G.; Glättli, A.; Koch, A.; Schlehuber, S. Mutations in the target protein conferring resistance to SDHI fungicides. In Modern Fungicides and Antifungal Compounds, Proceedings of the 16th International Reinhardsbrunn Symposium, Friedrichroda, Germany, 25–29 April 2010; Dehne, H.W., Deising, H.B., Gisi, U., Kuck, K.H., Russel, P.E., Lyr, H.G., Eds.; Deutsche Phytomedizinische Gesellschaft: Braunschweig, Germany, 2011; Volume VI. [Google Scholar]
- Scalliet, G.; Bowler, J.; Luksch, T.; Kirchhofer-Allan, L.; Steinhauer, D.; Ward, K.; Niklaus, M.; Verras, A.; Csukai, M.; Daina, A.; et al. Mutagenesis and functional studies with succinate dehydrogenase inhibitors in the wheat pathogen Mycosphaerella graminicola. PLoS ONE 2012, 7, e35429. [Google Scholar] [CrossRef] [Green Version]
- Garnault, M.; Duplaix, C.; Leroux, P.; Couleaud, G.; Carpentier, F.; David, O.; Walker, A.-S. Spatiotemporal dynamics of fungicide resistance in the wheat pathogen Zymoseptoria tritici in France. Pest Manag. Sci. 2019, 75, 1794–1807. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fungicides | Effect | df | F | p |
|---|---|---|---|---|
| All a | Fungicide (F) | 7 | 234.034 | <0.0001 |
| Year (Y) | 3 | 451.174 | <0.0001 | |
| F × Y | 21 | 50.110 | <0.0001 | |
| Multi-site inhibitors b | Fungicide (F) | 1 | 21.568 | <0.0001 |
| Year (Y) | 3 | 1.814 | 0.1627 | |
| F × Y | 3 | 1.310 | 0.2865 | |
| DMIs–Triazoles c | Fungicide (F) | 2 | 4.026 | 0.0233 |
| Year (Y) | 3 | 148.165 | <0.0001 | |
| F × Y | 6 | 14.291 | <0.0001 | |
| DMIs–Imidazoles d | Year | 3 | 12.147 | <0.0001 |
| SDHIs e | Fungicide (F) | 1 | 265.212 | <0.0001 |
| Year (Y) | 3 | 599.195 | <0.0001 | |
| F × Y | 3 | 10.906 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birr, T.; Hasler, M.; Verreet, J.-A.; Klink, H. Temporal Changes in Sensitivity of Zymoseptoria tritici Field Populations to Different Fungicidal Modes of Action. Agriculture 2021, 11, 269. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030269
Birr T, Hasler M, Verreet J-A, Klink H. Temporal Changes in Sensitivity of Zymoseptoria tritici Field Populations to Different Fungicidal Modes of Action. Agriculture. 2021; 11(3):269. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030269
Chicago/Turabian StyleBirr, Tim, Mario Hasler, Joseph-Alexander Verreet, and Holger Klink. 2021. "Temporal Changes in Sensitivity of Zymoseptoria tritici Field Populations to Different Fungicidal Modes of Action" Agriculture 11, no. 3: 269. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030269