
Changes in Bacterial and Fungal Soil Communities in Long-Term Organic Cropping Systems

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Description and Sampling
2.2. Soil Properties and Crop Yield
2.3. DNA Extraction, PCR, and Sequencing
2.4. Sequencing Data Processing
2.5. Fungal Pathogen Detection by qPCR
2.6. Statistical Analysis
3. Results
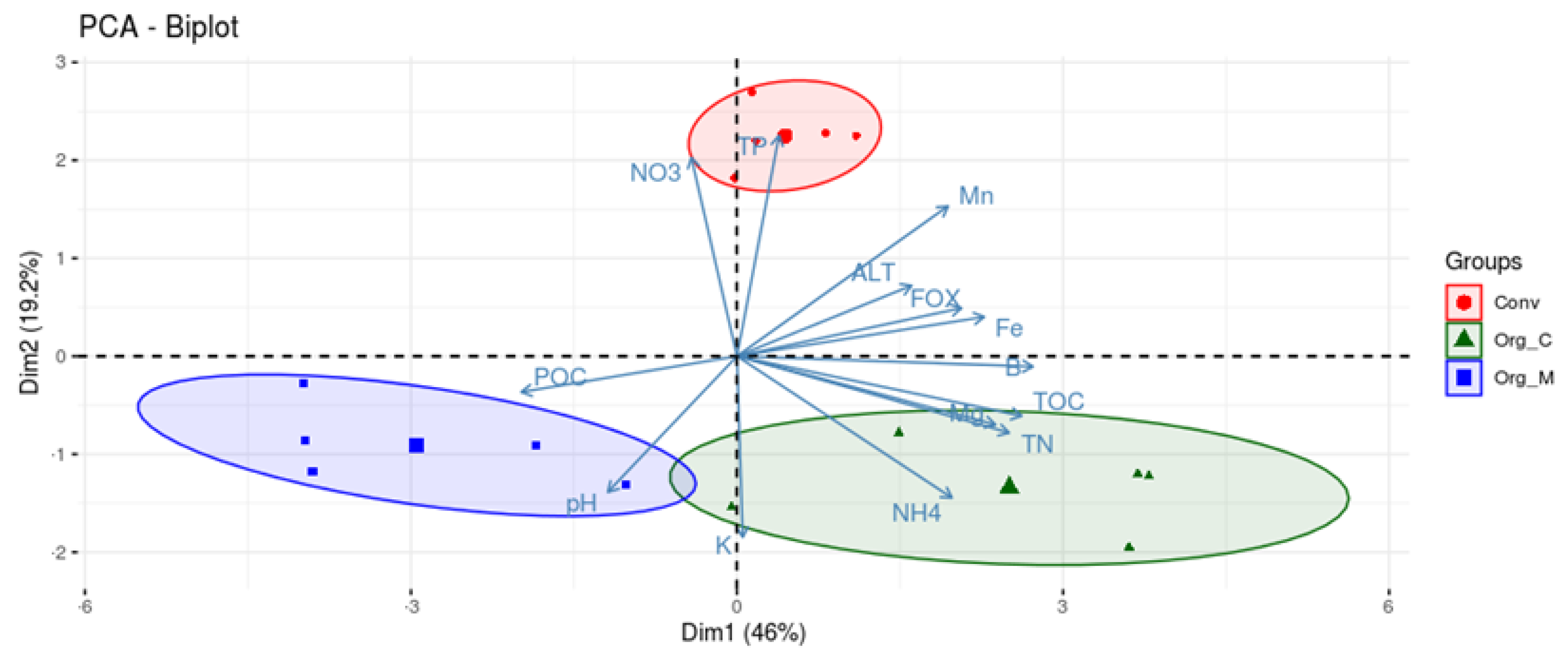
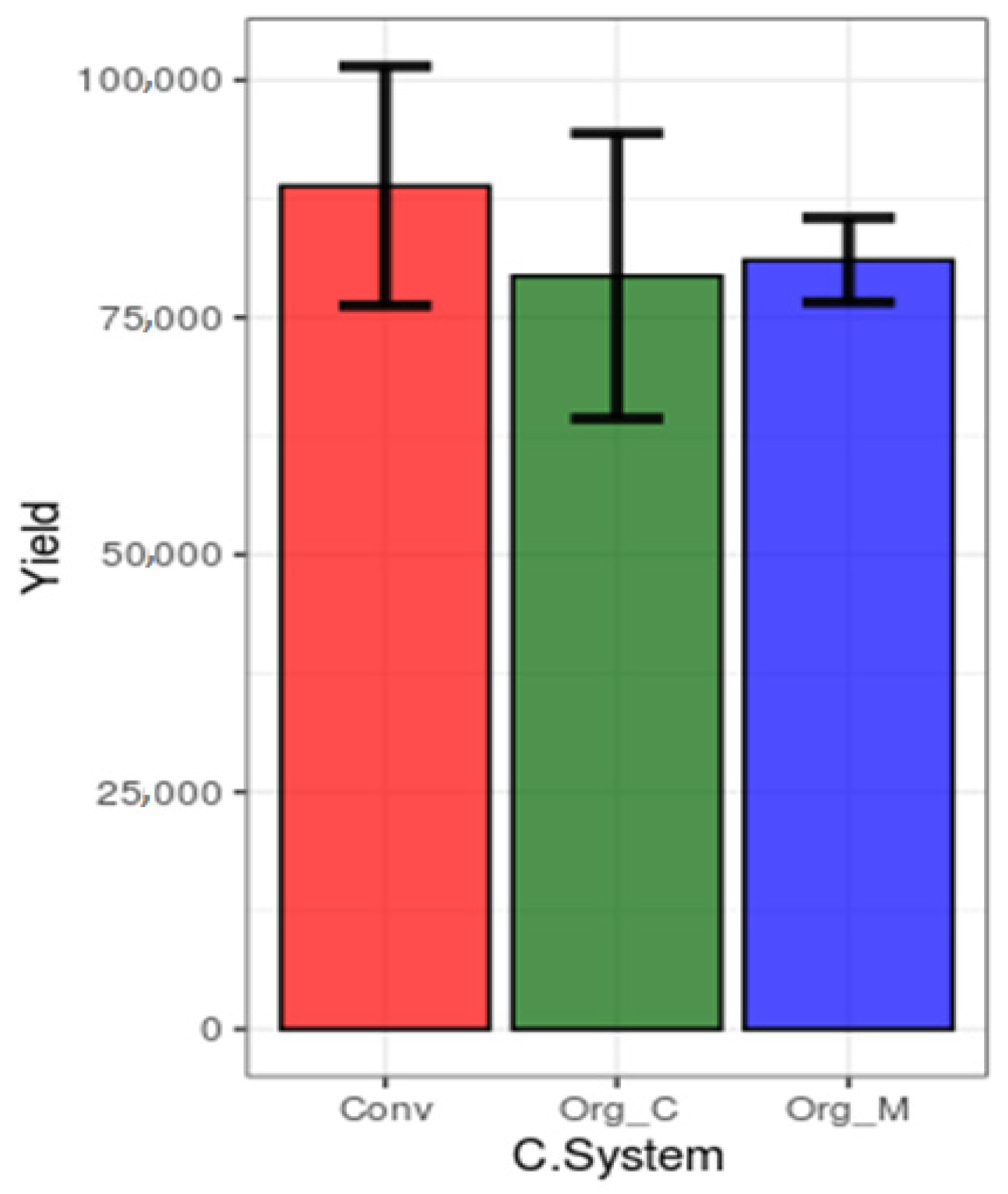
3.1. Effects of Different Cropping Systems on Soil Physico-Chemical Properties, Soil Pathogens, and Crop Yield
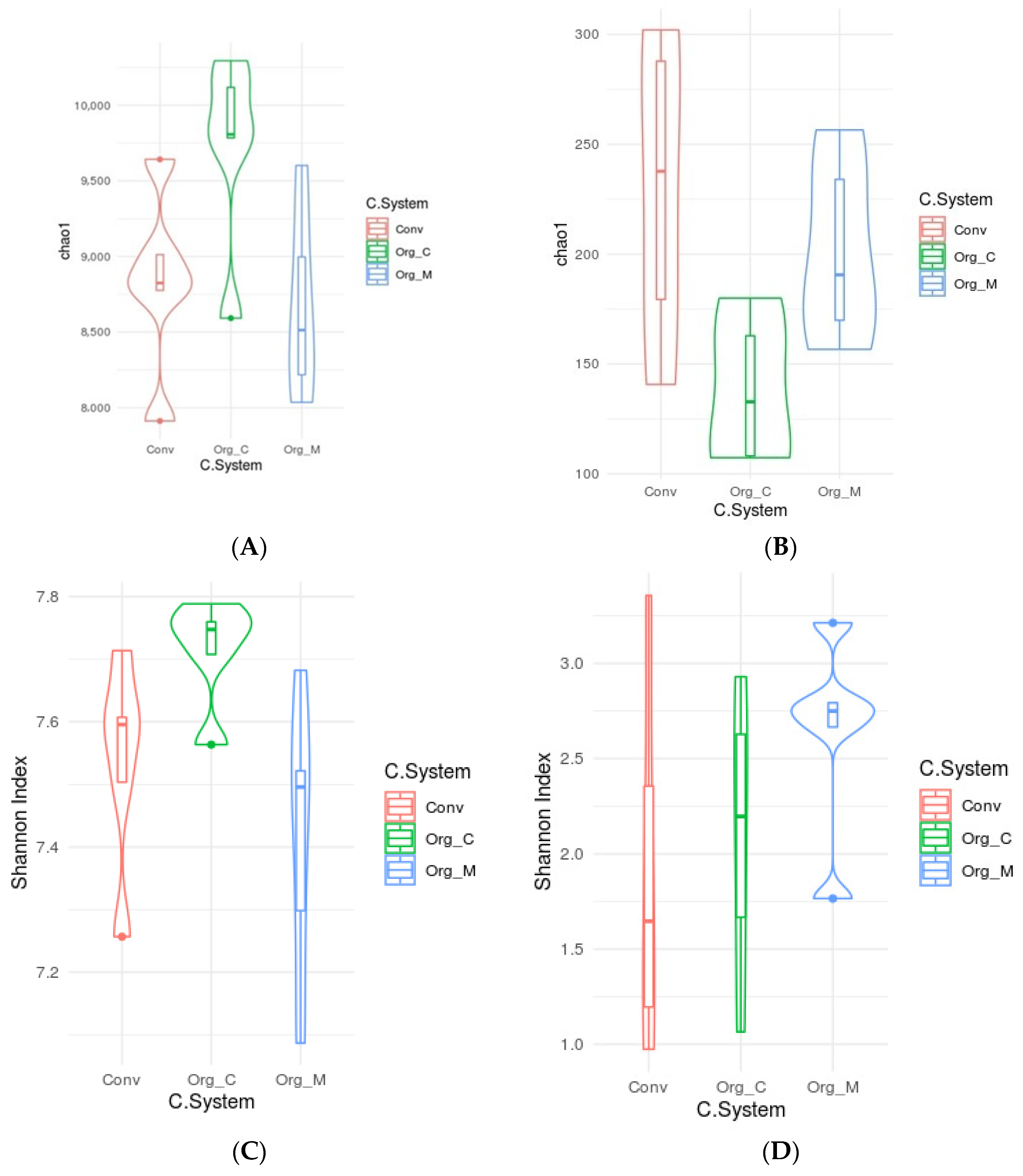
3.2. Effects of Different Cropping Systems on Soil Microbial Diversity
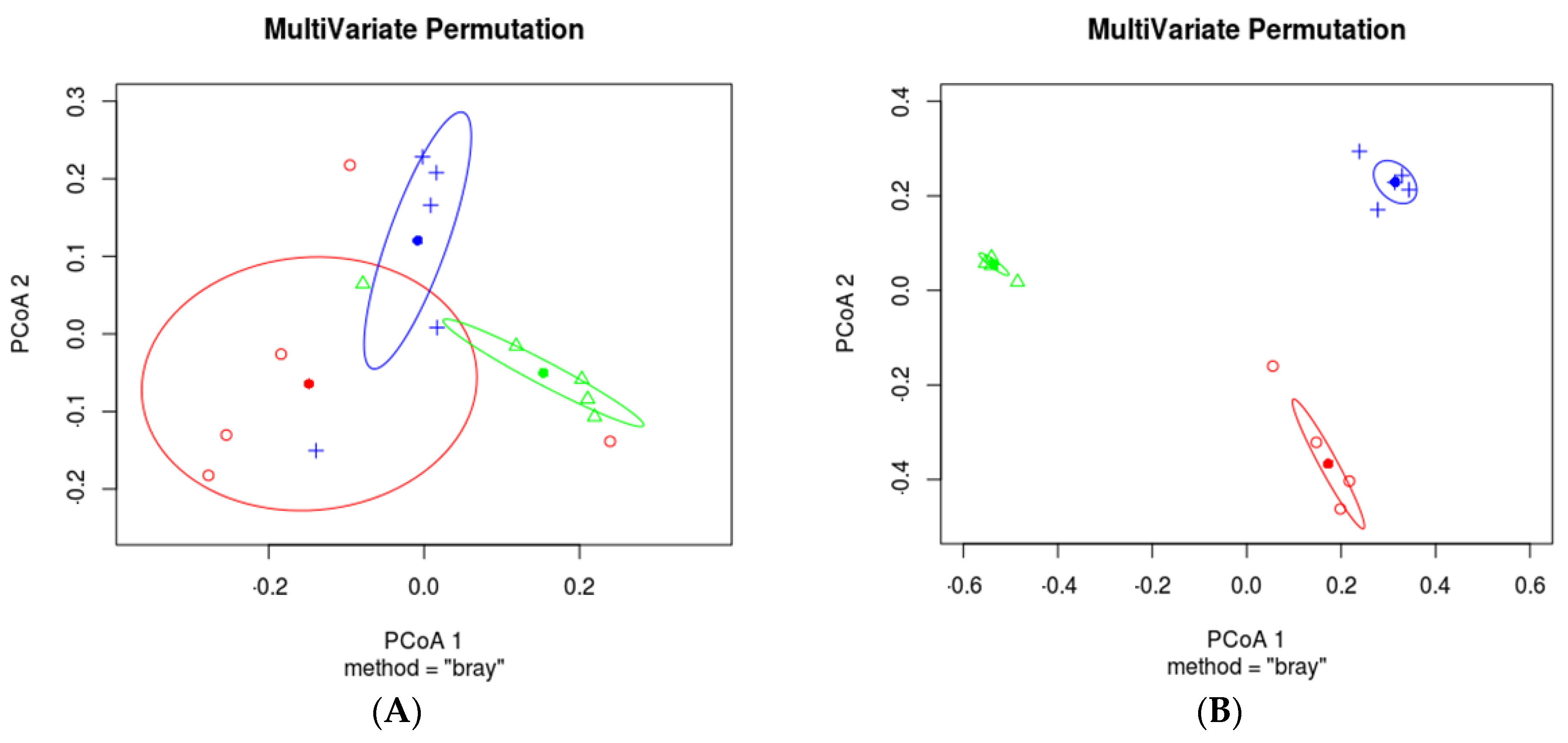
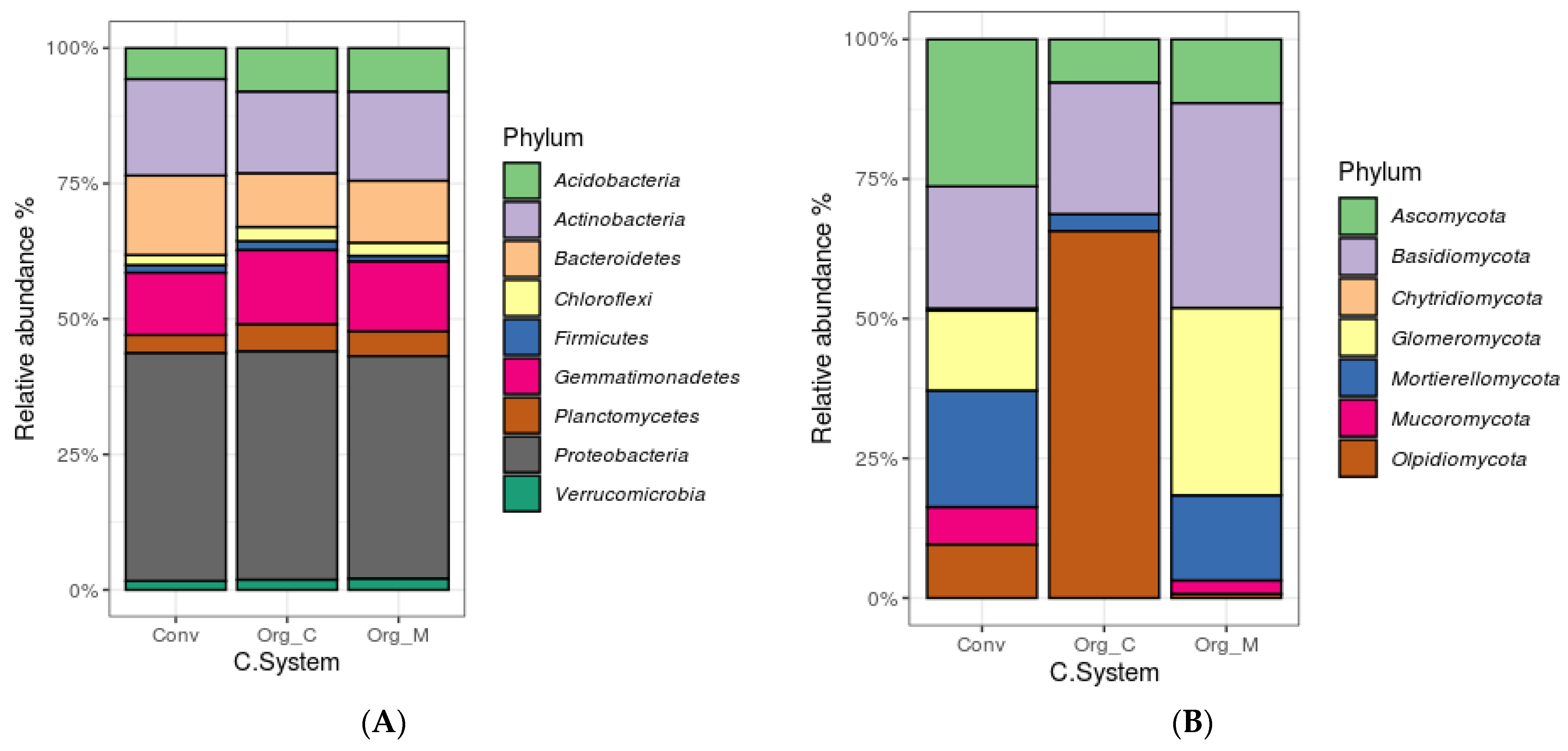
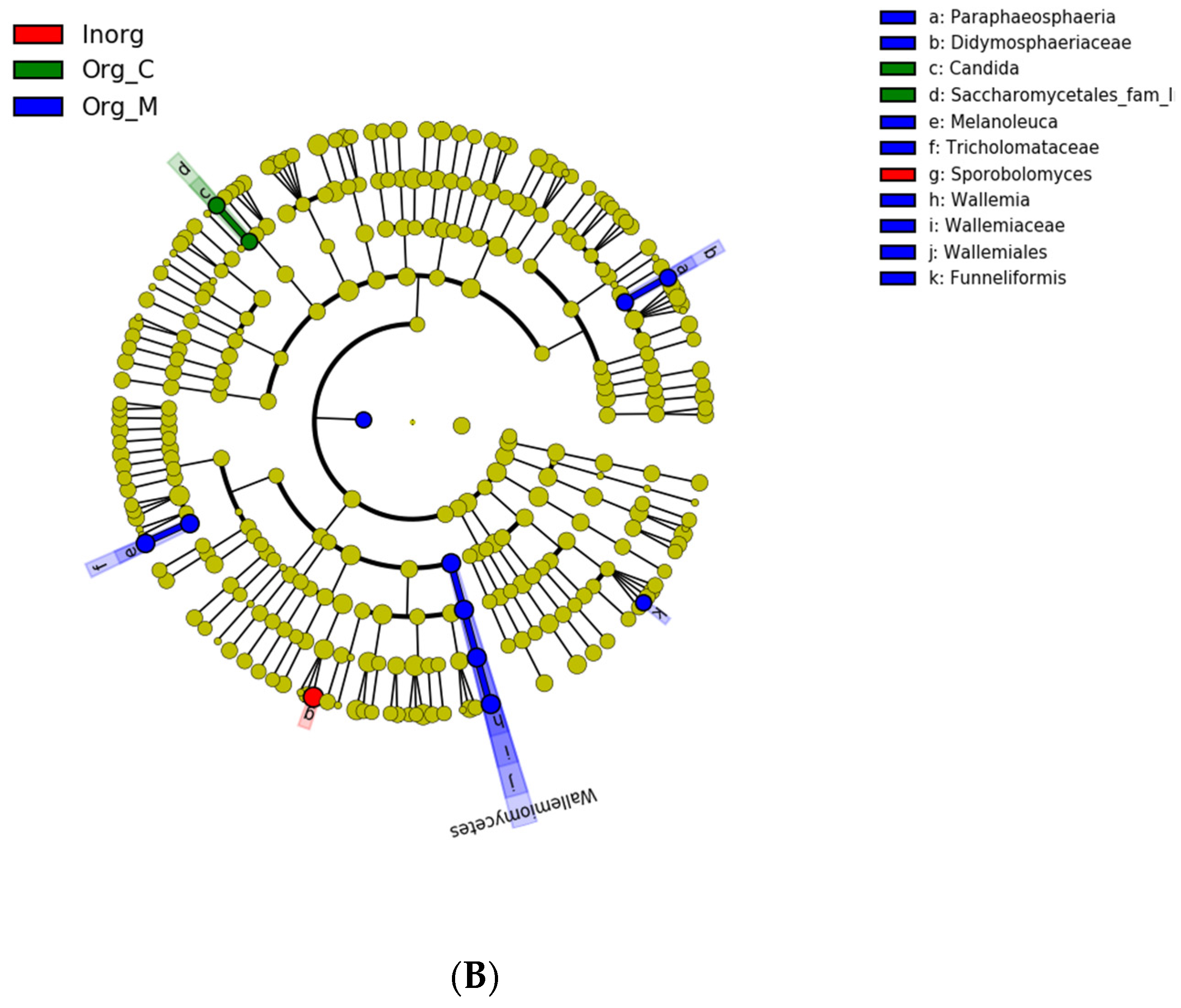
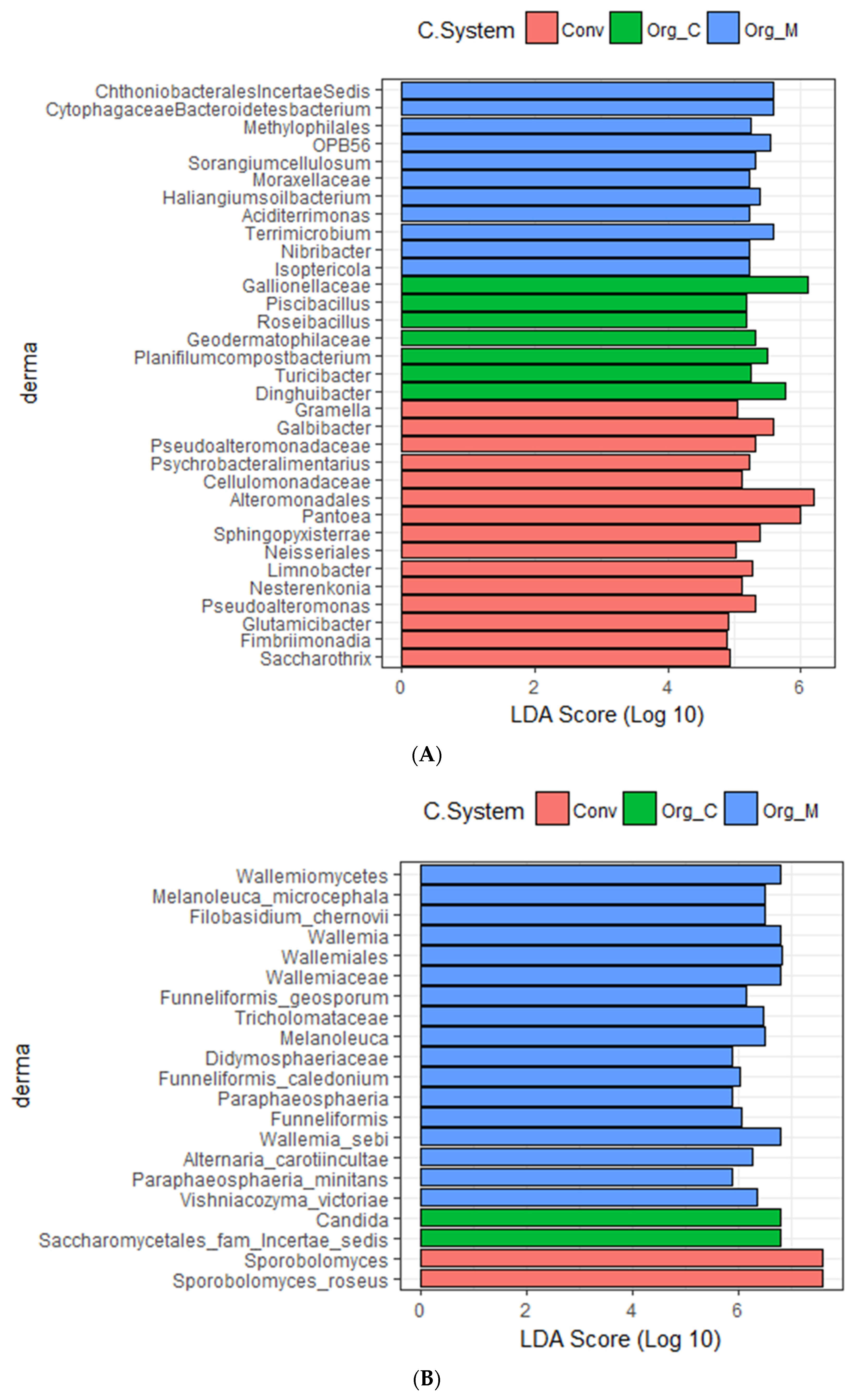
3.3. Effects of Different Cropping Systems on Bacterial and Fungal Community Composition
3.4. The Relationship between the Microbial Community and Soil Properties
4. Discussion
5. Practical Implications of This Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ITPS FAO. Food Agric. In Status of the World’s Soil Resources (SWSR) Main Report; Organ. United Nations Intergov. Tech. Panel Soils: Rome, Italy, 2015. [Google Scholar]
- Le Campion, A.; Oury, F.-X.; Heumez, E.; Rolland, B. Conventional versus organic farming systems: Dissecting comparisons to improve cereal organic breeding strategies. Org. Agric. 2020, 10, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Bonou-zin, R.D.C.; Allali, K.; Fadlaoui, A. Environmental Efficiency of Organic and Conventional Cotton in Benin. Sustainability 2019, 11, 3044. [Google Scholar] [CrossRef] [Green Version]
- Tully, K.L.; McAskill, C. Promoting soil health in organically managed systems: A review. Org. Agric. 2020, 10, 339–358. [Google Scholar] [CrossRef]
- Eurostat. Organic Farming Statistics. Available online: http://ec.europa.eu/eurostat/statistics-explained/index.php/Organic_farming_statistics (accessed on 15 January 2019).
- De Ponti, T.; Rijk, B.; Van Ittersum, M.K. The crop yield gap between organic and conventional agriculture. Agric. Syst. 2012, 108, 1–9. [Google Scholar] [CrossRef]
- Bonanomi, G.; De Filippis, F.; Cesarano, G.; La Storia, A.; Ercolini, D.; Scala, F. Organic farming induces changes in soil microbiota that affect agro-ecosystem functions. Soil Biol. Biochem. 2016, 103, 327–336. [Google Scholar] [CrossRef]
- Luo, G.; Li, L.; Friman, V.-P.; Guo, J.; Guo, S.; Shen, Q.; Ling, N. Organic amendments increase crop yields by improving microbe-mediated soil functioning of agroecosystems: A meta-analysis. Soil Biol. Biochem. 2018, 124, 105–115. [Google Scholar] [CrossRef]
- Dangi, S.R.; Bañuelos, G.; Buyer, J.S.; Hanson, B.; Gerik, J. Microbial community biomass and structure in saline and non-saline soils associated with salt-and boron-tolerant poplar clones grown for the phytoremediation of selenium. Int. J. Phytoremediat. 2018, 20, 129–137. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2017, 68, 12–26. [Google Scholar] [CrossRef]
- Schrama, M.; De Haan, J.J.; Kroonen, M.; Verstegen, H.; Van der Putten, W.H. Crop yield gap and stability in organic and conventional farming systems. Agric. Ecosyst. Environ. 2018, 256, 123–130. [Google Scholar] [CrossRef]
- Wagner, S.; Cattle, S.R.; Scholten, T. Soil-aggregate formation as influenced by clay content and organic-matter amendment. J. Plant Nutr. Soil Sci. 2007, 170, 173–180. [Google Scholar] [CrossRef]
- Chaudhry, V.; Rehman, A.; Mishra, A.; Chauhan, P.S.; Nautiyal, C.S. Changes in bacterial community structure of agricultural land due to long-term organic and chemical amendments. Microb. Ecol. 2012, 64, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Reilly, K.; Cullen, E.; Lola-Luz, T.; Stone, D.; Valverde, J.; Gaffney, M.; Brunton, N.; Grant, J.; Griffiths, B.S. Effect of organic, conventional and mixed cultivation practices on soil microbial community structure and nematode abundance in a cultivated onion crop. J. Sci. Food Agric. 2013, 93, 3700–3709. [Google Scholar] [CrossRef] [Green Version]
- Gonthier, D.J.; Ennis, K.K.; Farinas, S.; Hsieh, H.-Y.; Iverson, A.L.; Batáry, P.; Rudolphi, J.; Tscharntke, T.; Cardinale, B.J.; Perfecto, I. Biodiversity conservation in agriculture requires a multi-scale approach. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, L.W.; Sparling, B.; Tenuta, M.; Entz, M.H. Soil profile carbon and nutrient stocks under long-term conventional and organic crop and alfalfa-crop rotations and re-established grassland. Agric. Ecosyst. Environ. 2012, 158, 156–163. [Google Scholar] [CrossRef]
- Krishnaraj, P.U.; Sabale, S.N. Effect of organic and inorganic fertilization on soil bacterial diversity associated with sole crop (Pigeon pea) and crop rotation (Green gram-Sorghum). J. Pharmacogn. Phytochem. 2019, 8, 577–581. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; In World Soil Resources Reports; FAO: Rome, Italy, 2015. [Google Scholar]
- ISO HRN. Soil Quality—Determination of Dry Bulk Density; Croat Stand Institute: Zagreb, Croat, 2017. [Google Scholar]
- De Angelis, M. The Beginning of History: Value Struggles and Global Capital; Pluto Press: London, UK, 2007; ISBN 0745320368. [Google Scholar]
- ISO. Soil Quality: Determination of the Potential Cation Exchange Capacity and Exchangeable Cations Using Barium Chloride Solution Buffered at PH; ISO 13536:1995; Available online: https://www.iso.org/standard/22180.html (accessed on 5 January 2021).
- Van Genuchten, M.T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef] [Green Version]
- UGT. UGT Products. Soil Science; UGT: Müncheberg, Germany, 2018; Available online: http://www.ugt-online.de/en/products/soil-science/ (accessed on 10 March 2017).
- Cambardella, C.A.; Elliott, E.T. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Chen, H.; Hou, R.; Gong, Y.; Li, H.; Fan, M.; Kuzyakov, Y. Effects of 11 years of conservation tillage on soil organic matter fractions in wheat monoculture in Loess Plateau of China. Soil Tillage Res. 2009, 106, 85–94. [Google Scholar] [CrossRef]
- Keeney, D.R.; Nelson, D.W. Nitrogen—Inorganic forms. Methods Soil Anal. Part 2 Chem. Microbiol. Prop. 1983, 9, 643–698. [Google Scholar]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; U.S. Department of Agriculture: Washington, DC, USA, 1954.
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2000; ISBN 1420039903. [Google Scholar]
- Reichman, S.M. The Responses of Plants to Metal Toxicity: A Review Forusing on Copper, Manganese & Zinc; Australian Minerals & Energy Environment Foundation Melbourne: Melbourne, Australia, 2002; ISBN 187620513X.
- Porta Casanellas, J.; López Acevedo Requerin, M.; Rodríguez Ochoa, R. Técnicas y Experimentos en Edafología; Romanyá/Valls: Capellades, Spain, 1986; ISBN 846004341X. [Google Scholar]
- Anastassiades, M.; Lehotay, S.J.; Štajnbaher, D.; Schenck, F.J. Fast and easy multiresidue method employing acetonitrile extraction/partitioning and “dispersive solid-phase extraction” for the determination of pesticide residues in produce. J. AOAC Int. 2003, 86, 412–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taskin, B.; Gozen, A.G.; Duran, M. Selective quantification of viable Escherichia coli bacteria in biosolids by quantitative PCR with propidium monoazide modification. Appl. Environ. Microbiol. 2011, 77, 4329–4335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.P.; Peay, K.G. Sequence depth, not PCR replication, improves ecological inference from next generation DNA sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Bragg, L.; Stone, G.; Imelfort, M.; Hugenholtz, P.; Tyson, G.W. Fast, accurate error-correction of amplicon pyrosequences using Acacia. Nat. Methods 2012, 9, 425. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comeau, A.M.; Douglas, G.M.; Langille, M.G.I. Microbiome helper: A custom and streamlined workflow for microbiome research. MSystems 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing (Version 3.5.3); R Core Team: Boston, MA, USA, 2019. [Google Scholar]
- Ogle, D.H. FSA: Fisheries Stock Analysis. R Packag. Version 0.8.30. Available online: https://github.com/droglenc/FSA (accessed on 5 January 2019).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 35095. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. R Package Version 2020, 1, 337–354. [Google Scholar]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 1.17-4. Available online: http//CRAN.R-project.org/package=vegan2010 (accessed on 14 June 2019).
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; Van Den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Jannoura, R.; Joergensen, R.G.; Bruns, C. Organic fertilizer effects on growth, crop yield, and soil microbial biomass indices in sole and intercropped peas and oats under organic farming conditions. Eur. J. Agron. 2014, 52, 259–270. [Google Scholar] [CrossRef]
- Cai, A.; Xu, M.; Wang, B.; Zhang, W.; Liang, G.; Hou, E.; Luo, Y. Manure acts as a better fertilizer for increasing crop yields than synthetic fertilizer does by improving soil fertility. Soil Tillage Res. 2019, 189, 168–175. [Google Scholar] [CrossRef]
- Irshad, M.; Eneji, A.E.; Hussain, Z.; Ashraf, M. Chemical characterization of fresh and composted livestock manures. J. Soil Sci. Plant Nutr. 2013, 13, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Adeleke, R.; Nwangburuka, C.; Oboirien, B. Origins, roles and fate of organic acids in soils: A review. S. Afr. J. Bot. 2017, 108, 393–406. [Google Scholar] [CrossRef]
- Ge, G.; Li, Z.; Fan, F.; Chu, G.; Hou, Z.; Liang, Y. Soil biological activity and their seasonal variations in response to long-term application of organic and inorganic fertilizers. Plant Soil 2010, 326, 31. [Google Scholar] [CrossRef]
- Li, J.; Cooper, J.M.; Li, Y.; Yang, X.; Zhao, B. Soil microbial community structure and function are significantly affected by long-term organic and mineral fertilization regimes in the North China Plain. Appl. Soil Ecol. 2015, 96, 75–87. [Google Scholar] [CrossRef]
- Gu, S.; Gruau, G.; Dupas, R.; Rumpel, C.; Creme, A.; Fovet, O.; Gascuel-Odoux, C.; Jeanneau, L.; Humbert, G.; Petitjean, P. Release of dissolved phosphorus from riparian wetlands: Evidence for complex interactions among hydroclimate variability, topography and soil properties. Sci. Total Environ. 2017, 598, 421–431. [Google Scholar] [CrossRef]
- Yang, H.; Meng, Y.; Feng, J.; Li, Y.; Zhai, S.; Liu, J. Direct and indirect effects of long-term ditch-buried straw return on soil bacterial community in a rice–wheat rotation system. Land Degrad. Dev. 2020, 31, 851–867. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Q.; Chen, Y.; Dai, Q.; Hu, J. Dynamic change in enzyme activity and bacterial community with long-term rice cultivation in mudflats. Curr. Microbiol. 2019, 76, 361–369. [Google Scholar] [CrossRef]
- Liu, X.; Rashti, M.R.; Dougall, A.; Esfandbod, M.; Van Zwieten, L.; Chen, C. Subsoil application of compost improved sugarcane yield through enhanced supply and cycling of soil labile organic carbon and nitrogen in an acidic soil at tropical Australia. Soil Tillage Res. 2018, 180, 73–81. [Google Scholar] [CrossRef]
- Vera, A.; Moreno, J.L.; García, C.; Morais, D.; Bastida, F. Boron in soil: The impacts on the biomass, composition and activity of the soil microbial community. Sci. Total Environ. 2019, 685, 564–573. [Google Scholar] [CrossRef]
- Vera, A.; Moreno, J.L.; Siles, J.A.; López-Mondejar, R.; Zhou, Y.; Li, Y.; García, C.; Nicolás, E.; Bastida, F. Interactive impacts of boron and organic amendments in plant-soil microbial relationships. J. Hazard. Mater. 2021, 408, 124939. [Google Scholar] [CrossRef]
- Buckley, D.H.; Schmidt, T.M. The structure of microbial communities in soil and the lasting impact of cultivation. Microb. Ecol. 2001, 42, 11–21. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnström, J.; Weibull, A. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Jackson, C.R.; Vallaire, S.C. Microbial activity and decomposition of fine particulate organic matter in a Louisiana cypress swamp. J. N. Am. Benthol. Soc. 2007, 26, 743–753. [Google Scholar] [CrossRef]
- Kremen, C.; Miles, A. Ecosystem services in biologically diversified versus conventional farming systems: Benefits, externalities, and trade-offs. Ecol. Soc. 2012, 17, 40. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef] [Green Version]
- Allison, V.J.; Miller, R.M.; Jastrow, J.D.; Matamala, R.; Zak, D.R. Changes in soil microbial community structure in a tallgrass prairie chronosequence. Soil Sci. Soc. Am. J. 2005, 69, 1412–1421. [Google Scholar] [CrossRef] [Green Version]
- Lupatini, M.; Korthals, G.W.; de Hollander, M.; Janssens, T.K.S.; Kuramae, E.E. Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 2017, 7, 2064. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Espíndola, I.P.; Ferrara-Guerrero, M.J.; Luna-Guido, M.L.; Ramírez-Villanueva, D.A.; León-Lorenzana, D.; Arit, S.; Gómez-Acata, S.; González-Terreros, E.; Ramírez-Barajas, B.; Navarro-Noya, Y.E. The Bacterial Community Structure and Microbial Activity in a Traditional Organic Milpa Farming System Under Different Soil Moisture Conditions. Front. Microbiol. 2018, 9, 2737. [Google Scholar] [CrossRef]
- Bastida, F.; Hernández, T.; Albaladejo, J.; García, C. Phylogenetic and functional changes in the microbial community of long-term restored soils under semiarid climate. Soil Biol. Biochem. 2013, 65, 12–21. [Google Scholar] [CrossRef]
- Štursová, M.; Žifčáková, L.; Leigh, M.B.; Burgess, R.; Baldrian, P. Cellulose utilization in forest litter and soil: Identification of bacterial and fungal decomposers. FEMS Microbiol. Ecol. 2012, 80, 735–746. [Google Scholar] [CrossRef]
- Tang, H.; Li, C.; Xiao, X.; Shi, L.; Cheng, K.; Wen, L.; Li, W. Effects of short-term manure nitrogen input on soil microbial community structure and diversity in a double-cropping paddy field of southern China. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: London, UK, 2010; ISBN 0080559344. [Google Scholar]
- Zhu, H.; Liu, X.; Feng, Z.; Yao, Q. Promoting Crop Growth With Symbiotic Microbes in Agro-Ecosystems in Climate Change Era. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 31–41. [Google Scholar]
- Santísima-Trinidad, A.B.L.; del Mar Montiel-Rozas, M.; Diéz-Rojo, M.Á.; Pascual, J.A.; Ros, M. Impact of foliar fungicides on target and non-target soil microbial communities in cucumber crops. Ecotoxicol. Environ. Saf. 2018, 166, 78–85. [Google Scholar] [CrossRef]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2016, 2, 242. [Google Scholar] [CrossRef]
- Lian, T.; Ma, Q.; Shi, Q.; Cai, Z.; Zhang, Y.; Cheng, Y.; Nian, H. High aluminum stress drives different rhizosphere soil enzyme activities and bacterial community structure between aluminum-tolerant and aluminum-sensitive soybean genotypes. Plant Soil 2019, 440, 409–425. [Google Scholar] [CrossRef]
- Stagnari, F.; Perpetuini, G.; Tofalo, R.; Campanelli, G.; Leteo, F.; Della Vella, U.; Schirone, M.; Suzzi, G.; Pisante, M. Long-term impact of farm management and crops on soil microorganisms assessed by combined DGGE and PLFA analyses. Front. Microbiol. 2014, 5, 644. [Google Scholar] [CrossRef] [Green Version]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef] [Green Version]
- Offret, C.; Desriac, F.; Le Chevalier, P.; Mounier, J.; Jégou, C.; Fleury, Y. Spotlight on antimicrobial metabolites from the marine bacteria Pseudoalteromonas: Chemodiversity and ecological significance. Mar. Drugs 2016, 14, 129. [Google Scholar] [CrossRef] [Green Version]
- Chroňáková, A.; Schloter-Hai, B.; Radl, V.; Endesfelder, D.; Quince, C.; Elhottová, D.; Šimek, M.; Schloter, M. Response of archaeal and bacterial soil communities to changes associated with outdoor cattle overwintering. PLoS ONE 2015, 10, e0135627. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Saveliev, A.; Grigoryeva, T.; Boulygina, E.; Selivanovskaya, S. Fungal and bacterial successions in the process of co-composting of organic wastes as revealed by 454 pyrosequencing. PLoS ONE 2017, 12, e0186051. [Google Scholar]
- Rosenberg, E.; DeLong, E.F.; Lory, S.; Stackebrandt, E.; Thompson, F. The Prokaryotes: Actinobacteria; Springer: New York, NY, USA, 2014; ISBN 364230138X. [Google Scholar]
- Kundim, B.A.; Itou, Y.; Sakagami, Y.; Fudou, R.; Iizuka, T.; Yamanaka, S.; Ojika, M. New haliangicin isomers, potent antifungal metabolites produced by a marine myxobacterium. J. Antibiot. 2003, 56, 630–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Zhao, S.; Liu, Q.; Wang, H.; Wang, Y. Premliminary investigation of RNase activity and antifungal activity of {\sl Wallemia sebi}. Wei Sheng Wu Xue Tong Bao 2006, 33, 30–33. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivation System | Conv | Org_C | Org_M |
|---|---|---|---|
| Geographical coordinates | 37°48′18.5″ N, 0°51′49.2″ W | 37°51′39.3″ N, 0°54′03.3″ W | 37°49′30.2″ N, 0°52′28.4″ W |
| Crop 2017–2018 season | Brassica oleracea var. sabellica | ||
| Harvest | Manual on 20–25 February 2018. Crop residues were incorporated in the soil | ||
| Crops grown in previous years | Apium graveolens/Cucumils melo (2016/2017) Lactuca sativa/Brassica oleracea var. Italica (2015/2016) Apium graveolens/Cucumils melo (2014/2015) Brassica oleracea var. Italica/Capsicum annum (2013/2014) Foeniculum vulgare/Cucurbita moschata (2012/2013) | ||
| Organic amendments (amount per year) | 15,000 kg ha−1 sheep manure | 10,000 kg ha−1 sheep compost; compost tea * | 15,000 kg ha−1 sheep |
| Fertilizers (amount per year) | 15 kg ha−1 ENTEC solub 21 (ammonium sulfate with inhibition of nitrification); 10 L ha−1 phosphoric acid; 15 kg ha−1 calcium nitrate; 10 L ha−1 nitric acid | 10 L ha−1 EcoZen NPK 2-2-7 (aminoacids); 10 L ha−1 Sunfol veg agri 12% (aminoacids) | |
| Pesticides | Linuron; Indoxacarb 30%; Cypermethrin;Lambda cihalotrin 10%; Imidacloprid 20%; Spinosad; Azadirachtin 3.2%; Emamectin 0.85%; Clortalonil 50%; Difenoconazol 25%; Azoxystrobin 2.5%; Propamocarb 52%; Ciflufenamid | No application of chemical pesticides | |
| Soil Properties | Cropping System | Anova | Kruskal–Wallis | ||
|---|---|---|---|---|---|
| Conv | Org_C | Org_M | |||
| pH | 8.39 ± 0.17 b | 8.47 ± 0.14 ab | 8.70 ± 0.10 a | * | - |
| EC (dS m−1) | 0.54 ± 0.15 | 0.52 ± 0.13 | 0.38 ± 0.04 | - | ns |
| TOC (g kg−1) | 11.49 ± 0.28 ab | 15.64 ± 3.37 a | 9.01 ± 3.49 b | ** | - |
| TN (g kg−1) | 1.13 ± 0.19 b | 1.59 ± 0.34 a | 0.93 ± 0.24 b | ** | - |
| POC (g kg−1) | 2.67 ± 0.72 ab | 2.20 ± 0.55 b | 4.03 ± 1.40 a | * | - |
| NH4+ (mg kg−1) | 0.10 ± 0.23 b | 1.33 ± 0.15 b | 0.00 ± 0.00 a | - | ** |
| NO3− (mg kg−1) | 53.04 ± 28.27 a | 11.86 ± 7.10 ab | 27.00 ± 13.57 b | - | * |
| Bulk density (kg dm−3) | 1.24 ± 0.06 | 1.27 ± 0.07 | 1.34 ± 0.09 | ns | - |
| SWW (cm3 cm−3) | 0.12 ± 0.01 | 0.12 ± 0.02 | 0.12 ± 0.03 | ns | - |
| SWFC (cm3 cm−3) | 0.22 ± 0.01 | 0.25 ± 0.03 | 0.22 ± 0.02 | ns | - |
| CEC (cmol kg−1) | 14.82 ± 0.86 | 17.47 ± 4.13 | 12.76 ± 2.20 | - | ns |
| CaCO3 (%) | 44.65 ± 2.71 | 45.54 ± 7.57 | 47.03 ± 1.92 | - | ns |
| FMA (cm3 cm3−1) | 0.17 ± 0.03 | 0.20 ± 0.06 | 0.19 ± 0.03 | - | ns |
| Ca (cmol kg−1) | 8.44 ± 0.83 | 10.03 ± 2.40 | 7.19 ± 1.49 | - | ns |
| Mg (cmol kg−1) | 3.54 ± 0.11 ab | 4.39 ± 1.09 a | 3.13 ± 0.54 b | * | - |
| K (cmol kg−1) | 0.62 ± 0.15 b | 0.85 ± 0.17 a | 0.78 ± 0.06 ab | * | - |
| Na (cmol kg−1) | 2.12 ± 0.32 | 2.19 ± 0.86 | 1.64 ± 0.23 | ns | - |
| P (mg kg−1) | 20.15 ± 5.24 | 14.65 ± 7.71 | 14.33 ± 7.48 | ns | - |
| Cu (mg kg−1) | 2.17 ± 0.74 | 3.17 ± 0.81 | 2.19 ± 0.75 | ns | - |
| Zn (mg kg−1) | 4.75 ± 2.99 | 5.46 ± 1.65 | 4.48 ± 0.91 | ns | - |
| Fe (mg kg−1) | 6.19 ± 2.91 a | 6.97 ± 2.69 b | 2.99 ± 1.24 a | - | * |
| Mn (mg kg−1) | 9.47 ± 1.03 a | 7.91 ± 2.37 a | 4.66 ± 0.36 b | *** | - |
| B (mg kg−1) | 1.68 ± 0.11 a | 1.94 ± 0.28 a | 1.31 ± 0.17 b | ** | - |
| TP (ng g−1) | 232.00 ± 146.54 a | 13.36 ± 9.11 b | 6.08 ± 5.21 b | - | ** |
| ALT (log copy ITS g−1 soil) | 4.24 ± 0.32 a | 4.33 ± 0.49 a | 2.05 ± 1.88 b | - | ** |
| RSO (log copy ITS g−1 soil) | 0.67 ± 1.51 | 2.72 ± 1.61 | 2.02 ± 1.86 | - | ns |
| FOX (log copy ITS g−1 soil) | 3.08 ± 1.73 a | 3.30 ± 0.44 a | 0.63 ± 1.41 ab | - | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuartero, J.; Özbolat, O.; Sánchez-Navarro, V.; Egea-Cortines, M.; Zornoza, R.; Canfora, L.; Orrù, L.; Pascual, J.A.; Vivo, J.-M.; Ros, M. Changes in Bacterial and Fungal Soil Communities in Long-Term Organic Cropping Systems. Agriculture 2021, 11, 445. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11050445
Cuartero J, Özbolat O, Sánchez-Navarro V, Egea-Cortines M, Zornoza R, Canfora L, Orrù L, Pascual JA, Vivo J-M, Ros M. Changes in Bacterial and Fungal Soil Communities in Long-Term Organic Cropping Systems. Agriculture. 2021; 11(5):445. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11050445
Chicago/Turabian StyleCuartero, Jessica, Onurcan Özbolat, Virginia Sánchez-Navarro, Marcos Egea-Cortines, Raúl Zornoza, Loredana Canfora, Luigi Orrù, Jose Antonio Pascual, Juana-María Vivo, and Margarita Ros. 2021. "Changes in Bacterial and Fungal Soil Communities in Long-Term Organic Cropping Systems" Agriculture 11, no. 5: 445. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11050445