
Allelic Variation of Soybean Maturity Genes E1–E4 in the Huang-Huai-Hai River Valley and the Northwest China

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Locations
2.2. Experimental Design and Data Collection
2.3. Genotyping of Maturity Loci
2.4. Statistical Analysis
3. Results
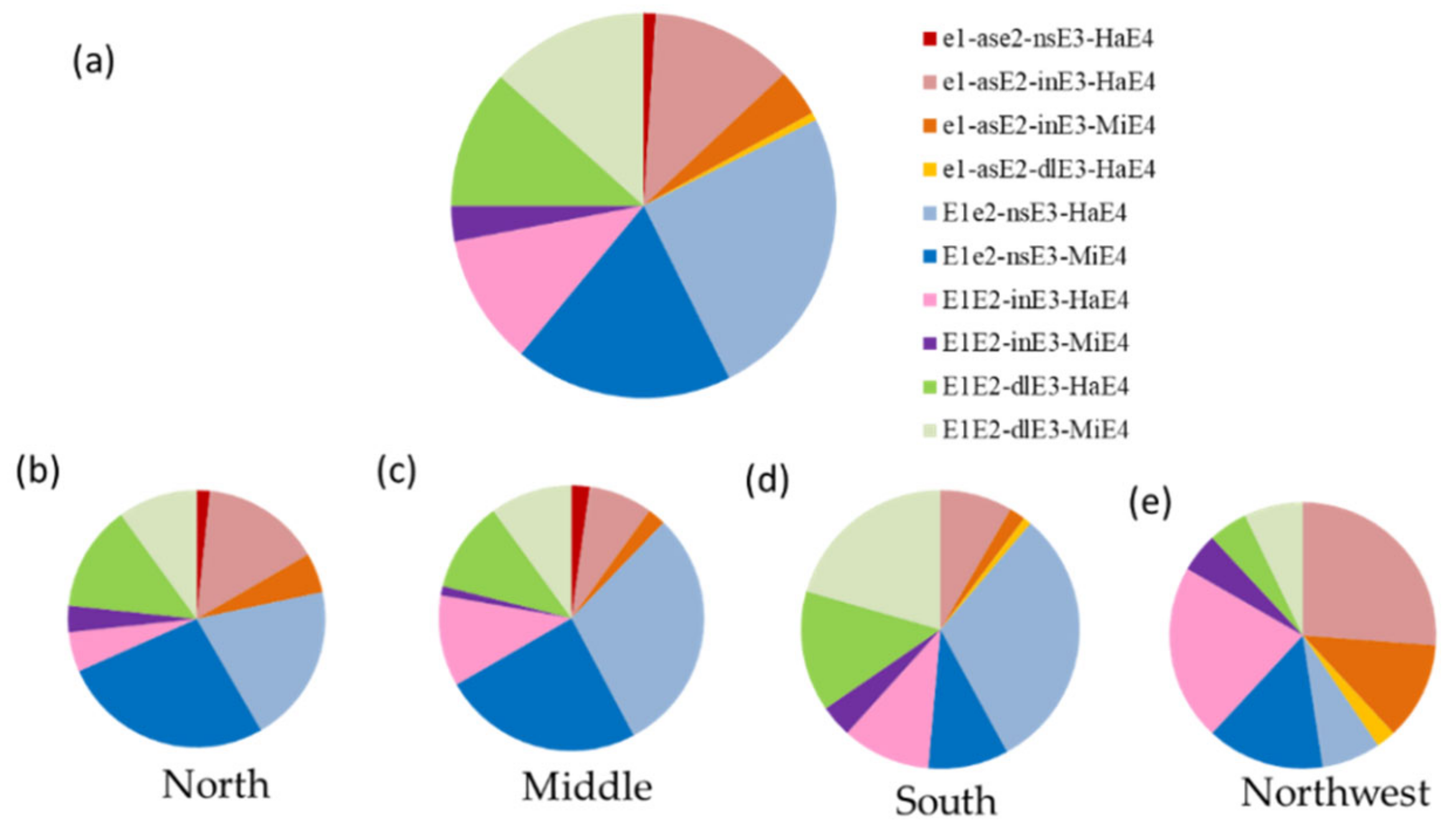
3.1. Genotyping of E1–E4 in the Huang-Huai-Hai River Valley and the Northwest China
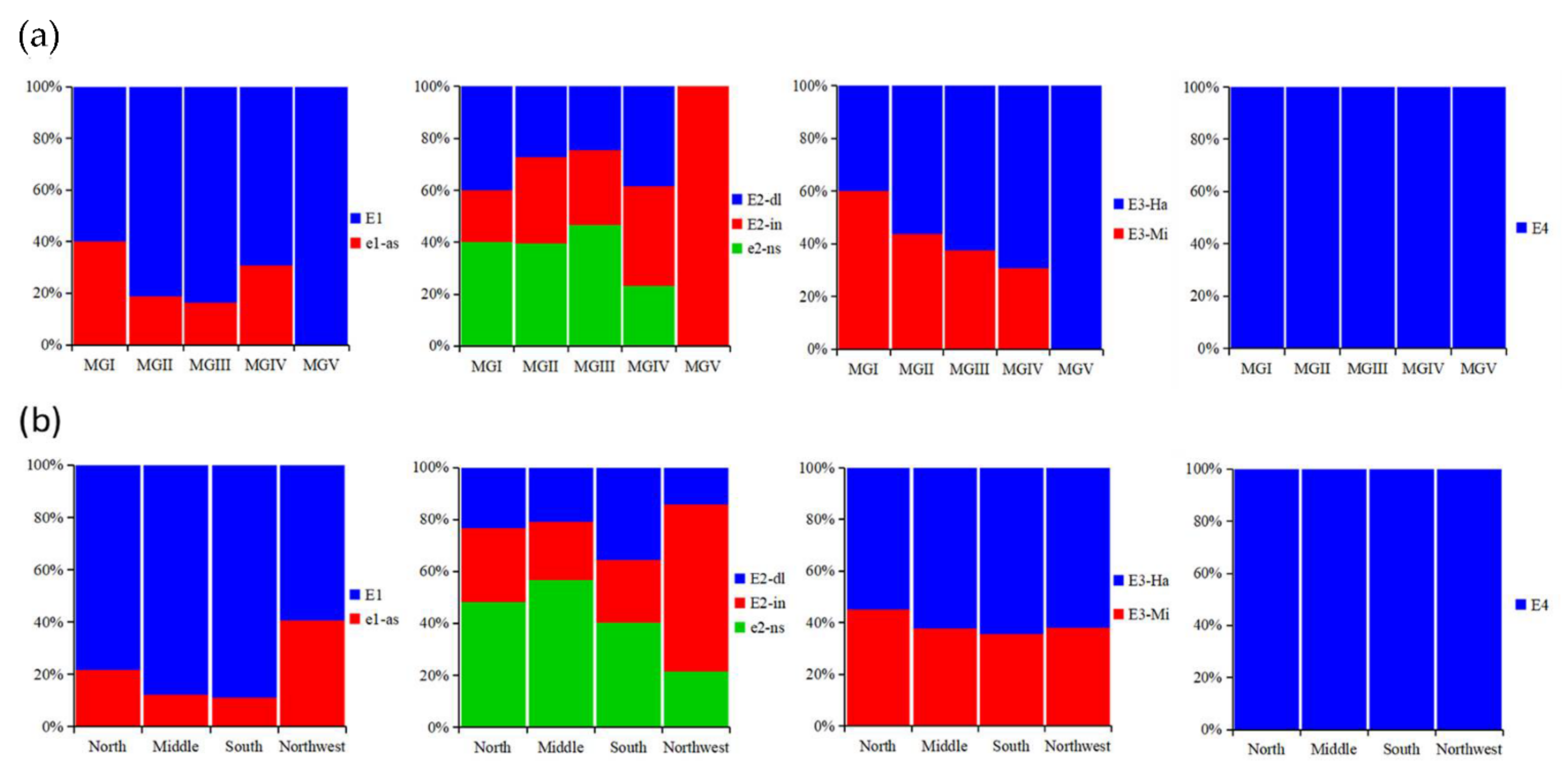
3.2. Distribution of E Genotypes in Different Maturity Groups and Sub-Regions
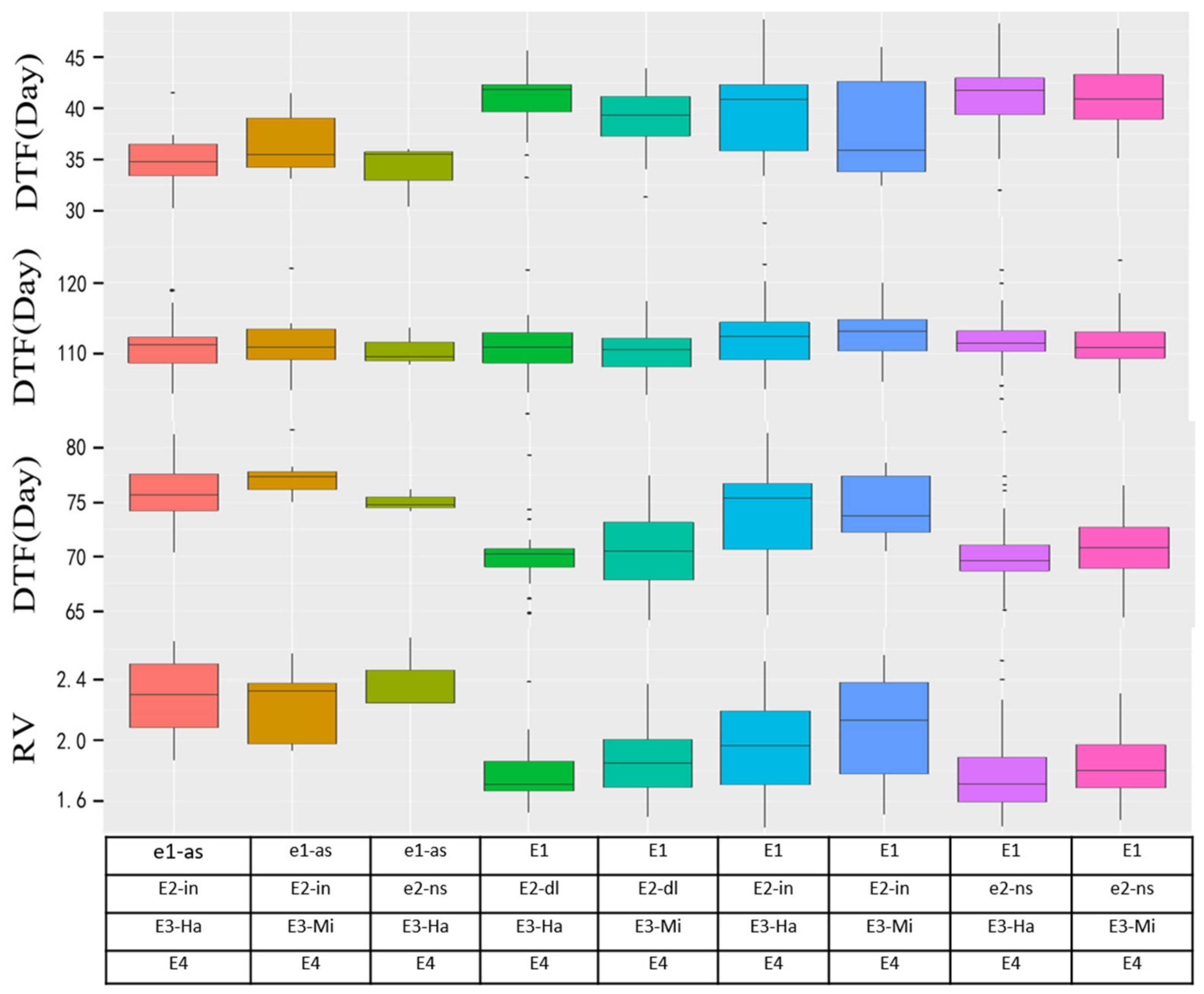
3.3. The Performance of Flowering and Maturity Time with Different E Allelic Combinations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Badole, S.L.; Bodhankar, S.L. Chapter 8—Glycine max (Soybean) Treatment for diabetes. In Bioactive Food as Dietary Interventions for Diabetes; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 77–82. [Google Scholar]
- Wilson, R.F. Soybean: Market driven research needs. In Genetics and Genomics of Soybean; Stacey, G., Ed.; Springer: New York, NY, USA, 2008; pp. 3–15. [Google Scholar]
- Langewisch, T.; Lenis, J.; Jiang, G.L.; Wang, D. The development and use of a molecular model for soybean maturity groups. BMC Plant Biol. 2017, 17, 91. [Google Scholar] [CrossRef]
- Bernard, R.L. Two major genes for time of flowering and maturity in soybeans. Crop Sci. 1971, 11, 242–244. [Google Scholar] [CrossRef]
- Buzzell, R.I. Inheritance of a soybean flowering response to fluorescentdaylength conditions. Can. J. Genet. Cytol. 1971, 13, 703–707. [Google Scholar] [CrossRef]
- Buzzell, R.I.; Voldeng, H.D. Inheritance of insensitivity to long daylength. Soyb. Genet. Newsl. 1980, 7, 26–29. [Google Scholar]
- McBlain, B.A.; Bernard, R.L. A new gene affecting the time of flowering and matuirty in soybean. J. Hered. 1987, 78, 160–162. [Google Scholar] [CrossRef]
- Bonato, E.R.; Vello, N.A. E6, a dominant gene conditioning early flowering and maturity in soybeans. Genet. Mol. Biol. 1999, 22, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Cober, E.R.; Voldeng, H.D. A new soybean maturity and photoperiod-sensitivity locus linked to E1 and T. Crop Sci. 2001, 41, 698–701. [Google Scholar] [CrossRef]
- Kong, F.; Nan, H.; Cao, D.; Li, Y.; Wu, F.; Wang, J.; Lu, S.; Yuan, X.; Cober, E.R.; Abe, J.; et al. A new dominant gene E9 conditions early flowering and maturity in soybean. Crop Sci. 2014, 54, 2529–2535. [Google Scholar] [CrossRef]
- Xia, Z.; Watanabe, S.; Yamada, T.; Tsubokura, Y.; Nakashima, H.; Zhai, H.; Anai, T.; Sato, S.; Yamazaki, T.; Lü, S.; et al. Positional cloning and characterization reveal the molecular basis for soybean maturity locus E1 that regulates photoperiodic flowering. PNAS 2012, 109, 12852–12853. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Xia, Z.; Hideshima, R.; Tsubokura, Y.; Sato, S.; Yamanaka, N.; Takahashi, R.; Anai, T.; Tabata, S.; Kitamura, K.; et al. A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene is involved in soybean maturity and flowering. Genetics 2011, 188, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Hideshima, R.; Xia, Z.; Tsubokura, Y.; Sato, S.; Nakamoto, Y.; Yamanaka, N.; Takahashi, R.; Ishimoto, M.; Anai, T.; et al. Map-based cloning of the gene associated with the soybean maturity locus E3. Genetics 2009, 182, 1251–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Kanazawa, A.; Matsumura, H.; Takahashi, R.; Harada, K.; Abe, J. Genetic redundancy in soybean photoresponses associated with duplication of the phytochrome A gene. Genetics 2008, 180, 995–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Xu, Z.; Liu, B.; Kong, F.; Jun, A. Genetic variation in four maturity genes affects photoperiod insensitivity and PHYA-regulated post-flowering responses of soybean. BMC Plant Biol. 2013, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Nan, H.; Gao, Y.; Tang, L.; Yue, Y. Allelic combinations of soybean maturity loci E1, E2, E3 and E4 result in diversity of maturity and adaptation to different latitudes. PLoS ONE 2014, 9, e106042. [Google Scholar]
- Li, J.; Wang, X.; Song, W.; Huang, X.; Qiu, L. Genetic variation of maturity groups and four E genes in the Chinese soybean mini core collection. PLoS ONE 2017, 12, e0172106. [Google Scholar] [CrossRef]
- Liu, L.; Song, W.; Wang, L.; Sun, X.; Han, T. Allele combinations of maturity genes E1-E4 affect adaptation of soybean to diverse geographic regions and farming systems in China. PLoS ONE 2020, 15, e0235397. [Google Scholar] [CrossRef]
- Kurasch, A.K.; Hahn, V.; Leiser, W.L.; Vollmann, J.; Schori, A.; Bétrix, C.-A.; Mayr, B.; Winkler, J.; Mechtler, K.; Aper, J.; et al. Identification of mega-environments in Europe and effect of allelic variation at maturity E loci on adaptation of European soybean. Plant Cell Environ. 2017, 40, 765–778. [Google Scholar] [CrossRef]
- Wolfgang, G.; An, Y.C. Genetic separation of southern and northern soybean breeding programs in North America and their associated allelic variation at four maturity loci. Mol. Breeding. 2017, 37, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tsubokura, Y.; Watanabe, S.; Xia, Z.; Kanamori, H.; Yamagata, H.; Kaga, A.; Katayose, Y.; Abe, J.; Ishimoto, M.; Harada, K. Natural variation in the genes responsible for maturity loci E1, E2, E3 and E4 in soybean. Ann. Bot. 2014, 113, 429–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubokura, Y.; Matsumura, H.; Xu, M.; Liu, B.; Nakashima, H.; Anai, T.; Kong, F.; Yuan, X.; Kanamori, H.; Katayose, Y.; et al. Genetic variation in soybean at the maturity locus E4 is involved in adaptation to long days at high latitudes. Agron. J. 2013, 3, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Zhai, H.; Lü, S.; Wu, H.; Zhang, Y.; Zhang, X.; Yang, J.; Wang, Y.; Yang, G.; Qiu, H.; Cui, T.; et al. Diurnal expression pattern, allelic variation, and association analysis reveal functional features of the e1 gene in control of photoperiodic flowering in soybean. PLoS ONE 2015, 10, e0135909. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breeding. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Thomson, M.J. High-Throughput SNP genotyping to accelerate crop improvement. Plant Breed Biotech. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Sedivy, E.J.; Wu, F.; Hanzawa, Y. Soybean domestication: The origin, genetic architecture and molecular bases. N. Phytol. 2017, 214, 539–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cober, E.R.; Tanner, J.W.; Voldeng, H.D. Genetic control of photoperiod response in early-maturing, near-isogenic soybean lines. Crop Sci. 1996, 36, 601–605. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MG | E allele Combinations | No. of Variety |
|---|---|---|
| I | e1-as/E2-in/E3-Ha/E4 | 1 |
| e1-as/e2-ns/E3-Ha/E4 | 1 | |
| E1E2-dl/E3-Mi/E4 | 2 | |
| E1/e2-ns/E3-Mi/E4 | 1 | |
| II | e1-as/E2-in/E3-Ha/E4 | 7 |
| e1-as/e2-ns/E3-Ha/E4 | 1 | |
| E1/E2-dl/E3-Ha/E4 | 4 | |
| E1/E2-dl/E3-Mi/E4 | 8 | |
| E1/E2-in/E3-Ha/E4 | 3 | |
| E1/E2-in/E3-Mi/E4 | 2 | |
| E1/e2-ns/E3-Ha/E4 | 7 | |
| E1/e2-ns/E3-Mi/E4 | 10 | |
| III | e1-asE2-in/E3-Ha/E4 | 17 |
| e1-as/E2-in/E3-Mi/E4 | 9 | |
| e1-as/e2-ns/E3-Ha/E4 | 1 | |
| E1/E2-dl/E3-Ha/E4 | 27 | |
| MG | E allele combinations | No. of Variety |
| III | E1/E2-dl/E3-Mi/E4 | 27 |
| E1/E2-in/E3-Ha/E4 | 18 | |
| E1/E2-in/E3-Mi/E4 | 6 | |
| E1/e2-ns/E3-Ha/E4 | 54 | |
| E1/e2-ns/E3-Mi/E4 | 34 | |
| IV | e1-as/E2-in/E3-Ha/E4 | 1 |
| e1-as/E2-in/E3-Mi/E4 | 1 | |
| E1/E2-dl/E3-Ha/E4 | 2 | |
| E1/E2-in/E3-Ha/E4 | 1 | |
| E1/E2-in/E3-Mi/E4 | 2 | |
| E1/e2-ns/E3-Ha/E4 | 2 | |
| E1/e2-ns/E3-Mi/E4 | 1 | |
| V | E1/E2-in/E3-Ha/E4 | 1 |
| All Regions | |||||
|---|---|---|---|---|---|
| DF | DTF a | DTM a | RP a | R/V a | |
| E1 | 1 | 53.50 ***b | 358.1 *** | 655.1 ** | 425.71 *** |
| E2 | 2 | 29.24 *** | 313.6 *** | 241.6 *** | 61.17 *** |
| E3 | 1 | 10.87 *** | 31.3 *** | 6.0 *c | 0.29 |
| Loc | 42 | 144.13 *** | 227.7 *** | 86.1 *** | 58.30 *** |
| E1 × Loc | 42 | 3.26 *** | 2.6 *** | 2.6 *** | 5.74 *** |
| E2 × Loc | 84 | 2.78 *** | 3.1 *** | 2.0 *** | 1.85 *** |
| E3 × Loc | 42 | 0.99 | 1.0 | 1.0 | 0.83 |
| North Region | |||||
| E1 | 1 | 129.62 *** | 0.36 | 93.39 ** | 181.55 *** |
| E2 | 2 | 10.49 *** | 2.81 | 8.49 **c | 12.73 *** |
| E3 | 1 | 0.03 | 1.11 | 1.46 | 1.46 |
| Loc | 11 | 39.35 *** | 55.52 *** | 52.71 *** | 57.01 *** |
| E1 × Loc | 11 | 1.73 | 1.13 | 1.90* | 3.49 *** |
| E2 × Loc | 22 | 1.02 | 0.73 | 0.80 | 1.18 |
| E3 × Loc | 11 | 0.52 | 0.90 | 0.82 | 0.88 |
| Middle Region | |||||
| E1 | 1 | 241.55 *** | 11.6 *** | 143.73 ** | 278.22 *** |
| E2 | 2 | 76.05 *** | 15.1 *** | 80.19 *** | 81.46 *** |
| E3 | 1 | 0.56 | 0.9 | 0.01 | 0.58 |
| Loc | 12 | 46.68 *** | 87.6 *** | 120.49 *** | 64.35 *** |
| E1 × Loc | 12 | 2.84 *** | 0.9 | 1.95 * | 4.22 *** |
| E2 × Loc | 24 | 1.25 | 2.3 *** | 1.58 * | 1.53 |
| E3 × Loc | 12 | 0.32 | 0.7 | 0.46 | 0.52 |
| South Region | |||||
| E1 | 1 | 276.50 *** | 0.7 | 142.38 *** | 378.75 *** |
| E2 | 2 | 16.55 *** | 3.8 * | 18.70 *** | 36.51 *** |
| E3 | 1 | 45.46 *** | 16.0 *** | 0.93 | 24.33 *** |
| Loc | 11 | 10.05 *** | 80.1 *** | 43.18 *** | 7.16 *** |
| E1 × Loc | 11 | 2.12 *d | 0.6 | 0.91 | 3.36 *** |
| E2 × Loc | 22 | 0.60 | 0.6 | 0.95 | 0.94 |
| E3 × Loc | 11 | 0.66 | 0.5 | 0.69 | 0.57 |
| Northwest Region | |||||
| E1 | 1 | 39.62 *** | 12.02 *** | 1.66 | 17.31 *** |
| E2 | 2 | 16.38 *** | 8.32 *** | 5.09 ** | 8.78 *** |
| E3 | 1 | 0.20 | 1.44 | 0.91 | 0.007 |
| Loc | 11 | 25.06 *** | 37.98 *** | 33.00 *** | 23.84 *** |
| E1 × Loc | 11 | 1.32 | 0.87 | 1.37 | 1.46 |
| E2 × Loc | 22 | 1.47 | 1.15 | 1.64 * | 1.64 * |
| E3 × Loc | 11 | 0.64 | 6.67 | 1.02 | 0.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wu, T.; Wen, H.; Song, W.; Xu, C.; Han, T.; Sun, S.; Wu, C. Allelic Variation of Soybean Maturity Genes E1–E4 in the Huang-Huai-Hai River Valley and the Northwest China. Agriculture 2021, 11, 478. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060478
Zhang X, Wu T, Wen H, Song W, Xu C, Han T, Sun S, Wu C. Allelic Variation of Soybean Maturity Genes E1–E4 in the Huang-Huai-Hai River Valley and the Northwest China. Agriculture. 2021; 11(6):478. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060478
Chicago/Turabian StyleZhang, Xinyue, Tingting Wu, Huiwen Wen, Wenwen Song, Cailong Xu, Tianfu Han, Shi Sun, and Cunxiang Wu. 2021. "Allelic Variation of Soybean Maturity Genes E1–E4 in the Huang-Huai-Hai River Valley and the Northwest China" Agriculture 11, no. 6: 478. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060478