
Genome-Wide Association Study Reveals Genetic Basis of Trace Elements Accumulation in Maize Kernels

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Trace Element Contents Detected in Maize Kernels and Soil
2.3. Phenotypic and Genotypes Analysis
2.4. Genome-Wide Association Analysis
2.5. Prediction of Candidate Genes
2.6. Statistical Analysis
3. Results
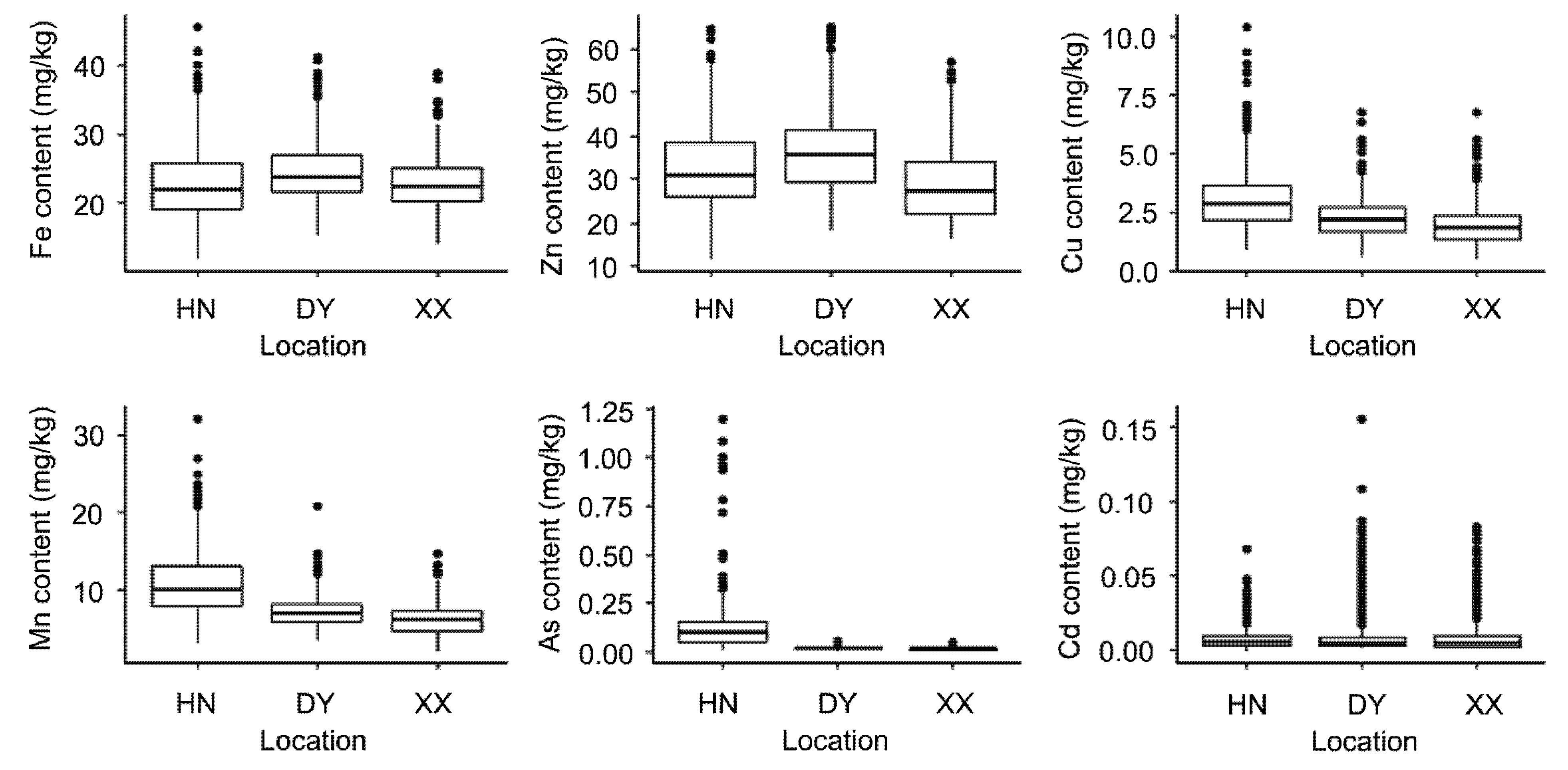
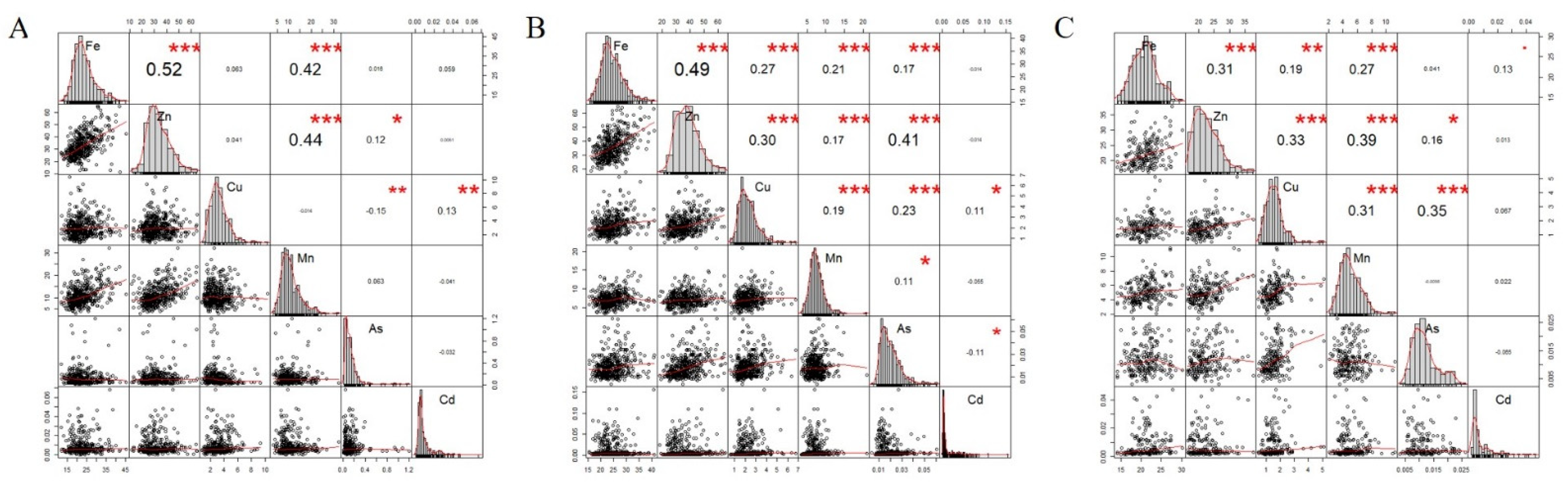
3.1. Phenotypic Analysis for Trace Elements’ Content of Maize Kernel in Different Locations
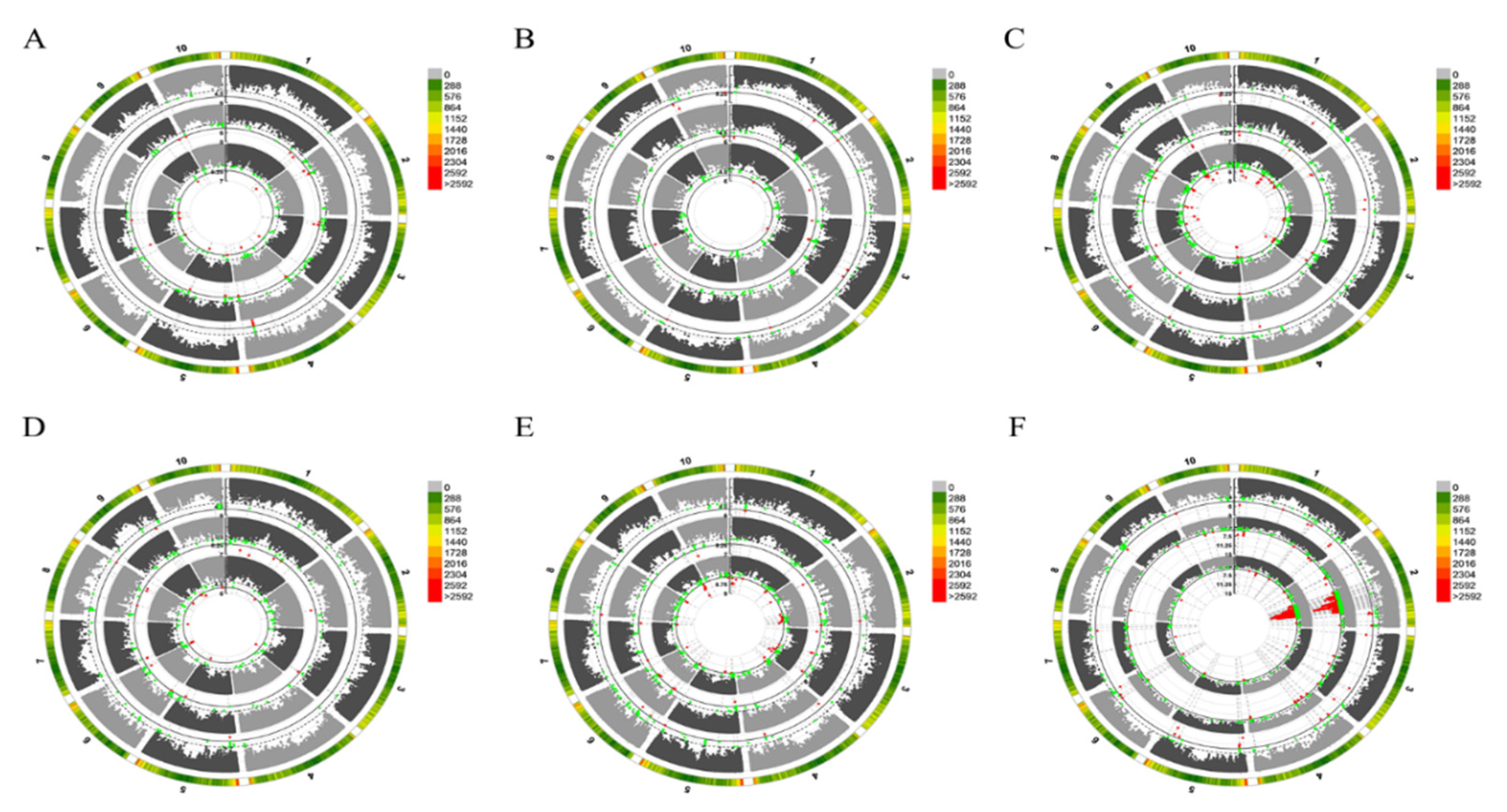
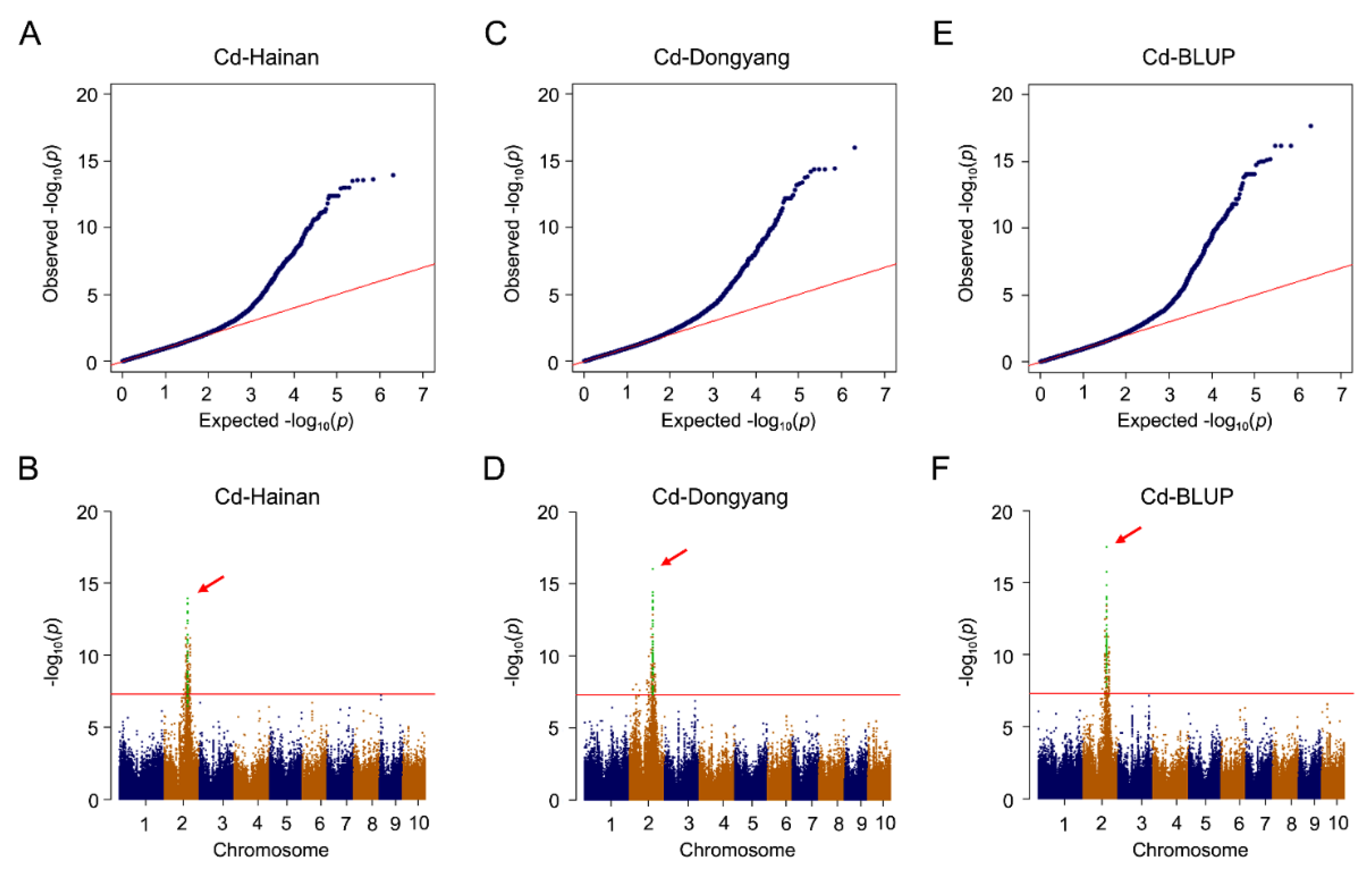
3.2. Genome-Wide Association Analysis of Trace Elements Accumulation in Kernels
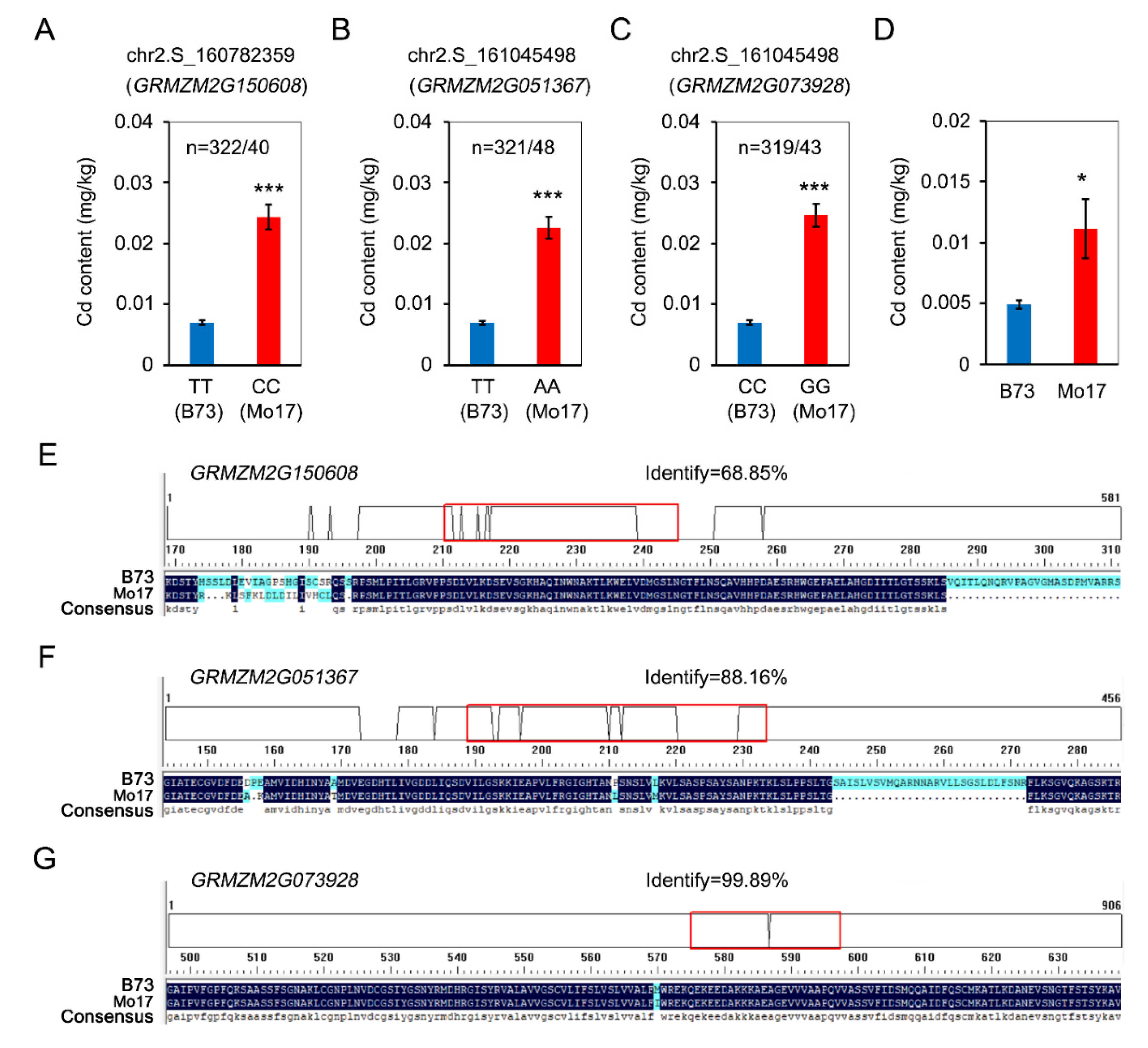
3.3. Identification of Causal Genes of Cd Accumulation
4. Discussion
4.1. Germplasm Resources for Cultivating New Maize Varieties with More Beneficial Trace Elements and Less Harmful Elements
4.2. Candidate Genes for Cultivating New High-Quality Maize Varieties with Higher Beneficial Elements and Less Harmful Elements
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zimmermann, M.B.; Hurrell, R.F. Improving iron, zinc and vitamin A nutrition through plant biotechnology. Curr. Opin. Biotechnol. 2002, 13, 142–145. [Google Scholar] [CrossRef]
- Godt, J.; Scheidig, F.; Grosse-Siestrup, C.; Esche, V.; Brandenburg, P.; Reich, A.; Groneberg, D.A. The toxicity of cadmium and resulting hazards for human health. J. Occup. Med. Toxicol. 2019, 1, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diepenbrock, C.H.; Gore, M.A. Closing the Divide between Human Nutrition and Plant Breeding. Crop Sci. 2015, 55, 1437–1448. [Google Scholar] [CrossRef]
- Wagner, G. Accumulation of Cadmium in Crop Plants And Its Consequences to Human Health. Adv. Agron. 1993, 51, 173–212. [Google Scholar]
- Feng, L.; Yan, H.; Dai, C.; Xu, W.; Gu, F.; Zhang, F.; Li, T.; Xian, J.; He, X.; Yu, Y.; et al. The systematic exploration of cadmium-accumulation characteristics of maize kernel in acidic soil with different pollution levels in China. Sci. Total Environ. 2020, 729, 138972. [Google Scholar] [CrossRef]
- Zhao, X.; Luo, L.; Cao, Y.; Liu, Y.; Li, Y.; Wu, W.; Lan, Y.; Jiang, Y.; Gao, S.; Zhang, Z.; et al. Genome-wide association analysis and QTL mapping reveal the genetic control of cadmium accumulation in maize leaf. BMC Genom. 2018, 19, 91. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, L.; Wang, W.; Li, T.; He, Z.; Yang, X. Current status of agricultural soil pollution by heavy metals in China: A meta-analysis. Sci. Total Environ. 2019, 651, 3034–3042. [Google Scholar] [CrossRef]
- Du, C.; Zhang, N.; Baokun, L.; Anqiang, C.; Yanting, M.; Wanli, H.; Bin, F.; Zhengxiang, Y.; Jun, C. Selection of varieties of Zea mays with low accumulation of heavy metals of arsenic, lead and cadmium. Southwest China J. Agric. Sci. 2017, 30, 5–10. [Google Scholar]
- Ohmori, Y.; Sotta, N.; Fujiwara, T. Identification of introgression lines of Oryza glaberrima Steud. with high mineral content in grains. Soil Sci. Plant Nutr. 2016, 62, 456–464. [Google Scholar] [CrossRef] [Green Version]
- Mahender, A.; Anandan, A.; Pradhan, S.K.; Pandit, E. Rice grain nutritional traits and their enhancement using relevant genes and QTLs through advanced approaches. SpringerPlus 2016, 5, 2086. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Pinson, S.R.; Tarpley, L.; Huang, X.Y.; Lahner, B.; Yakubova, E.; Baxter, I.; Guerinot, M.L.; Salt, D.E. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 2014, 127, 137–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Zeng, D.; Wang, B.; Qian, Q.; Zheng, S.; Ling, H.Q. Environmental effects on mineral accumulation in rice grains and identification of ecological specific QTLs. Environ. Geochem. Health 2013, 35, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Norton, G.J.; Duan, G.L.; Lei, M.; Zhu, Y.G.; Meharg, A.A.; Price, A.H. Identification of quantitative trait loci for rice grain element composition on an arsenic impacted soil: Influence of flowering time on genetic loci. Ann. Appl. Biol. 2012, 161, 46–56. [Google Scholar] [CrossRef]
- Lu, K.; Li, L.; Zheng, X.; Zhang, Z.; Mou, T.; Hu, Z. Quantitative trait loci controlling Cu, Ca, Zn, Mn and Fe content in rice grains. J. Genet. 2008, 87, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Hindu, V.; Palacios-Rojas, N.; Babu, R.; Suwarno, W.B.; Rashid, Z.; Usha, R.; Saykhedkar, G.R.; Nair, S.K. Identification and validation of genomic regions influencing kernel zinc and iron in maize. Appl. Genet. 2018, 131, 1443–1457. [Google Scholar] [CrossRef] [Green Version]
- Jin, T.; Chen, J.; Zhu, L.; Zhao, Y.; Guo, J.; Huang, Y. Comparative mapping combined with homology-based cloning of the rice genome reveals candidate genes for grain zinc and iron concentration in maize. BMC Genet. 2015, 16, 17. [Google Scholar] [CrossRef] [Green Version]
- Benke, A.; Urbany, C.; Marsian, J.; Shi, R.; Wiren, N.; Stich, B. The genetic basis of natural variation for iron homeostasis in the maize IBM population. BMC Plant Biol. 2014, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Jin, T.; Zhou, J.; Chen, J.; Zhu, L.; Zhao, Y.; Huang, Y. The genetic architecture of zinc and iron content in maize grains as revealed by QTL mapping and meta-analysis. Breed. Sci. 2013, 63, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Cai, Y.; Liu, Z.; Wang, G.; Wang, J.; Guo, Y.; Wang, H. Identification of QTL for zinc and iron concentration in maize kernel and cob. Euphytica 2012, 187, 345–358. [Google Scholar] [CrossRef]
- Lung’aho, M.G.; Mwaniki, A.M.; Szalma, S.J.; Hart, J.J.; Rutzke, M.A.; Kochian, L.V.; Glahn, R.P.; Hoekenga, O.A. Genetic and physiological analysis of iron biofortification in maize kernels. PLoS ONE 2011, 6, e20429. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Huang, Y.; Liu, Z.; Chen, J.; Zhu, L.; Song, Z.; Zhao, Y. Genetic analysis and QTL mapping of zinc, iron, copper and manganese contents in maize seed. J. Plant Genet. Resour. 2010, 11, 593–595. [Google Scholar]
- Zdunic, Z.; Grljusic, S.; Ledencan, T.; Duvnjak, T.; Simic, D. Quantitative trait loci mapping of metal concentrations in leaves of the maize IBM population. Hereditas 2014, 151, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Ueno, D.; Yamaji, N.; Kono, I.; Chao, F.H.; Ando, T.; Yano, M.; Jian, F.M. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Zhang, Z.; Chai, X.; Wang, H.; Wang, Y. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [CrossRef]
- Huang, X.; Han, B. Natural variations and genome-wide association studies in crop plants. Annu. Rev. Plant Biol. 2014, 65, 531–551. [Google Scholar] [CrossRef]
- Yan, J.; Warburton, M.; Crouch, J. Association mapping for enhancing maize (Zea mays L.) genetic improvement. Crop Sci. 2011, 51, 433–449. [Google Scholar] [CrossRef]
- Chao, D.Y.; Chen, Y.; Chen, J.; Shi, S.; Chen, Z.; Wang, C.; Danku, J.M.; Zhao, F.J.; Salt, D.E.; Maloof, J.N. Genome-wide Association Mapping Identifies a New Arsenate Reductase Enzyme Critical for Limiting Arsenic Accumulation in Plants. PLoS Biol. 2014, 12, e1002009. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.Y.; Silva, A.; Baxter, I.; Huang, Y.S.; Nordborg, M.; Danku, J.; Lahner, B.; Yakubova, E.; Salt, D.E.; Bomblies, K. Genome-Wide Association Studies Identify Heavy Metal ATPase3 as the Primary Determinant of Natural Variation in Leaf Cadmium in Arabidopsis thaliana. PLoS Genet. 2012, 8, e1002923. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Pettersson, M.; Rönnegård, L.; Carlborg, Ö.; Barsh, G.S. Inheritance Beyond Plain Heritability: Variance-Controlling Genes in Arabidopsis thaliana. PLoS Genet. 2012, 8, e1002839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsberg, S.; Andreatta, M.E.; Huang, X.Y.; Danku, J.; Salt, D.; Carlborg, Ö. The multi-allelic genetic architecture of a variance-heterogeneity locus for molybdenum concentration in leaves acts as a source of unexplained additive genetic variance. PLoS Genet. 2015, 11, e1005648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, G.J.; Douglas, A.; Lahner, B.; Yakubova, E.; Price, A.H. Genome wide association mapping of grain arsenic, copper, molybdenum and zinc in rice (Oryza sativa L.) grown at four international field sites. PLoS ONE 2014, 9, e89685. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, Z.; Kakar, K.U.; Li, X.B.; Li, S.; Zhang, B.; Shou, H.-X.; Shu, Q.-Y. Genome-wide Association Mapping of Quantitative Trait Loci (QTLs) for Contents of Eight Elements in Brown Rice (Oryza sativa L.). J. Agric. Food Chem. 2015, 63, 8008–8016. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lu, K.; Zhao, F.J.; Xie, W.B.; Ramakrishna, P.; Wang, G.Y.; Du, Q.Q.; Liang, L.M.; Sun, C.J.; Zhao, H.; et al. Genome-Wide Association Studies Reveal the Genetic Basis of Ionomic Variation in Rice. Plant Cell 2018, 30, 2720–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Lu, Y.; Yang, X.; Huang, J.; Zhou, Y.; Ali, F.; Wen, W.; Liu, J.; Li, J.; Yan, J. Genome wide association studies using a new nonparametric model reveal the genetic architecture of 17 agronomic traits in an enlarged maize association panel. PLoS Genet. 2014, 10, e1004573. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.B.; Nyquist, W.E.; Cervantes-Martínez, C.T. Estimating and Interpreting Heritability for Plant Breeding: An Update; John Wiley&Sons, Inc.: Oxford, UK, 2010; pp. 9–112. [Google Scholar]
- Liu, H.J.; Luo, X.; Niu, L.Y.; Xiao, Y.J.; Chen, L.; Liu, J.; Wang, X.Q.; Jin, M.L.; Li, W.Q.; Zhang, Q.H.; et al. Distant eQTLs and Non-coding Sequences Play Critical Roles in Regulating Gene Expression and Quantitative Trait Variation in Maize. Mol. Plant 2017, 10, 414–426. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Cao, L.; Lin, C.; Gao, Y.; Sun, C.; Zhang, Z. Health risk assessment of trace elements exposure through the soil-plant (maize)-human contamination pathway near a petrochemical industry complex, Northeast China. Environ. Pollut. 2020, 263, 114414. [Google Scholar] [CrossRef]
- Lv, W.; Meng, Y. Effect of microelement fertilizers on maize in calcareous region. Guangxi Agric. Sci. 2006, 37, 419–421. [Google Scholar]
- Li, D.; Sun, Z.; Wang, Y.; Li, T.; Xue, S.; Wang, X. Effect of NPK and microelement fertilizers on nutrient accumulation, distribution and yield of maize on high-fertility soil. Soil Fertil. Sci. China 2009, 6, 32–36. [Google Scholar]
- Li, D.; Ma, W.; Wei, J.; Mao, Y.; Chen, Q. Magnesium promotes root growth and increases aluminum tolerance via modulation of nitric oxide production in Arabidopsis. Plant Soil 2019, 457, 83–95. [Google Scholar] [CrossRef]
- Mai, N.; To, H. A genome-wide association study reveals the quantitative trait locus and candidate genes that regulate phosphate efficiency in a Vietnamese rice collection. Physiol. Mol. Biol. Plants 2020, 26, 2267–2281. [Google Scholar]
- Chong, K.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2010, 181, 71–78. [Google Scholar]
- Wu, D.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Ma, J.F. The HvNramp5 transporter mediates uptake of cadmium and manganese, but not iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Xu, W.; Xie, J.; Gao, Y.; He, Z. Variation of a major facilitator superfamily gene contributes to differential cadmium accumulation between rice subspecies. Nat. Commun. 2019, 10, 2562. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.; Luo, M.; Zhang, Y.; Guo, H.; Li, J.; Song, W.; Zhang, R.; Feng, Z.; Kong, M.; Li, H.; et al. Natural variations in the P-type ATPase heavy metal transporter gene ZmHMA3 control cadmium accumulation in maize grains. J. Exp. Bot. 2021, 72, 6230–6246. [Google Scholar] [CrossRef]
- Cao, Y.; Zhao, X.; Liu, Y.; Wang, Y.; Wu, W.; Jiang, Y.; Liao, C.; Xu, X.; Gao, S.; Shen, Y.; et al. Genome-wide identification of ZmHMAs and association of natural variation in ZmHMA2 and ZmHMA3 with leaf cadmium accumulation in maize. PeerJ 2019, 7, e7877. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.W.; Tan, L.M.; Zhang, C.J.; Hou, X.M.; Cai, X.W.; Chen, S.; He, X.J. FHA2 is a plant-specific ISWI subunit responsible for stamen development and plant fertility. J. Integr. Plant Biol. 2020, 62, 1703–1716. [Google Scholar] [CrossRef]
- Ahn, E.R.; Cho, H.K.; Pai, H.S. The forkhead-associated domain 2 (FHA2) in Arabidopsis plays a role in plant fertility by regulating stamen development. Planta 2013, 237, 1015–1023. [Google Scholar] [CrossRef]
- Cheng, Q.; Li, Y.; Jian, G.; Wang, W.; Zhang, H.; Yu, L.; Ping, W. OsDGL1, a homolog of an oligosaccharyltransferase complex subunit, is involved in N-glycosylation and root development in rice. Plant Cell Physiol. 2013, 54, 129–137. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fe (mg/kg) | Zn (mg/kg) | Cu (mg/kg) | Mn (mg/kg) | As (mg/kg) | Cd (mg/kg) | |
|---|---|---|---|---|---|---|
| HN | 3706.79 ± 301.22 b | 29.96 ± 3.49 b | 8.16 ± 1.61 b | 125.11 ± 7.55 c | 5.2 ± 0.70 c | 0.02 ± 0.01 c |
| DY | 14760.52 ± 182.45 a | 65.17 ± 2.41 a | 24.74 ± 0.19 a | 146.70 ± 2.89 b | 19.4 ± 0.35 a | 0.31 ± 0.01 a |
| XX | 33428.20 ± 562.40 b | 70.27 ± 4.15 a | 24.51 ± 0.12 a | 614.41 ± 12.09 a | 11.88 ± 0.37 b | 0.26 ± 0.01 b |
| Trait | Location | Heritability | Mean a | Range | CV (%) |
|---|---|---|---|---|---|
| Fe | HN | 0.98 | 22.91 a | 11.74–45.47 | 5.41 |
| DY | 0.95 | 24.54 b | 15.32–41.04 | 5.74 | |
| XX | 0.89 | 21.16 a | 14.09–38.89 | 7.58 | |
| Zn | HN | 0.98 | 32.66 b | 11.55–64.79 | 5.24 |
| DY | 0.98 | 36.38 c | 18.08–64.98 | 4.67 | |
| XX | 0.94 | 22.31 a | 16.33–57.01 | 6.47 | |
| Cu | HN | 0.87 | 3.08 c | 0.83–10.40 | 25.93 |
| DY | 0.94 | 2.29 b | 0.61–6.78 | 14.32 | |
| XX | 0.94 | 1.58 a | 0.45–6.78 | 16.18 | |
| Mn | HN | 0.99 | 11.00 c | 3.06–32.19 | 4.68 |
| DY | 0.99 | 7.23 b | 3.37–20.93 | 3.75 | |
| XX | 0.99 | 5.08 a | 2.09–14.76 | 4.23 | |
| As | HN | 0.95 | 0.13 b | 0.10–1.20 | 37.13 |
| DY | 0.96 | 0.02 a | 0.0037–0.061 | 14.55 | |
| XX | 0.98 | 0.01 a | 0.0031–0.05 | 7.55 | |
| Cd | HN | 0.87 | 8.59 × 103 a | 0–0.068 | 61.71 |
| DY | 0.98 | 1.21 × 102 b | 0.0012–0.16 | 28.34 | |
| XX | 0.99 | 7.20 × 103 ab | 0.001–0.083 | 12.52 |
| SNP | Gene ID | Associated Trace Element | Annotation |
|---|---|---|---|
| chr2.s_121584444 | GRMZM2G076905 | Zn Cd | uncharacterized |
| chr3.s_28354214 | GRMZM2G375197 | Zn Fe | uncharacterized |
| chr4.s_38571269 | GRMZM2G123257 | Cd Fe | S-adenosyl-L-methionine-dependent methyltransferase superfamily protein |
| chr4.s_38571275 | GRMZM2G123257 | Cd Fe | S-adenosyl-L-methionine-dependent methyltransferase superfamily protein |
| chr4.s_38571287 | GRMZM2G123257 | Cd Fe | S-adenosyl-L-methionine-dependent methyltransferase superfamily protein |
| chr4.s_38571537 | GRMZM2G123257 | Cd Fe | S-adenosyl-L-methionine-dependent methyltransferase superfamily protein |
| chr4.s_240465668 | GRMZM2G384293 | Zn Fe | embryo defective 14 |
| chr4.s_240465753 | GRMZM2G384293 | Zn Fe | embryo defective 14 |
| chr5.s_164728663 | GRMZM2G018820 | Mn Fe | glycerophosphoryl phosphodiesterase |
| chr5.s_164728686 | GRMZM2G018820 | Mn Fe | glycerophosphoryldiester phosphodiesterase |
| chr7.s_4960091 | GRMZM2G039922 | Zn Fe | uncharacterized |
| chr10.s_110087144 | GRMZM2G034206 | Zn Fe | Beta-13-N-Acetylglucosaminyltransferase family protein |
| Gene ID | Chr | Start | End | Annotation |
|---|---|---|---|---|
| GRMZM2G150608 | Chr2 | 160781682 | 160789474 | fha7—FHA-transcription factor 7 |
| GRMZM2G150754 | Chr2 | 160797324 | 160805739 | (MED33A, RFR1) REF4-related 1 |
| GRMZM2G171370 | Chr2 | 160956979 | 160963086 | bzip86—bZIP-transcription factor 86 |
| GRMZM2G100005 | Chr2 | 161016548 | 161017635 | Ribosomal protein S4 (RPS4A) family protein |
| GRMZM2G051367 | Chr2 | 161045370 | 161050327 | (DGL1) dolichyl-diphosphooligosaccharide-protein glycosyltransferase |
| GRMZM2G018241 | Chr2 | 161123950 | 161130108 | cesa9—cellulose synthase9 |
| GRMZM2G319836 | Chr2 | 161128260 | 161129517 | cesa9—cellulose synthase9 |
| GRMZM2G150404 | Chr2 | 161135362 | 161138301 | cesa9—cellulose synthase9 |
| GRMZM2G452739 | Chr2 | 161136453 | 161137710 | cesa9—cellulose synthase9 |
| GRMZM2G165133 | Chr2 | 161197745 | 161199034 | (FQR1) flavodoxin-like quinone reductase 1 |
| GRMZM2G073928 | Chr2 | 161273580 | 161276954 | Leucine-rich repeat protein kinase family protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, F.; Wang, N.; Bao, F.; Zhao, G.; Jing, L.; Wang, G.; Han, Q.; Hao, Z.; Chen, B. Genome-Wide Association Study Reveals Genetic Basis of Trace Elements Accumulation in Maize Kernels. Agriculture 2022, 12, 262. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020262
Zhao F, Wang N, Bao F, Zhao G, Jing L, Wang G, Han Q, Hao Z, Chen B. Genome-Wide Association Study Reveals Genetic Basis of Trace Elements Accumulation in Maize Kernels. Agriculture. 2022; 12(2):262. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020262
Chicago/Turabian StyleZhao, Fucheng, Nan Wang, Fei Bao, Guangwu Zhao, Liquan Jing, Guiyue Wang, Qinghui Han, Zhuanfang Hao, and Bin Chen. 2022. "Genome-Wide Association Study Reveals Genetic Basis of Trace Elements Accumulation in Maize Kernels" Agriculture 12, no. 2: 262. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020262