
Gene Locus Mapping and Candidate Gene Screening for Branched Spike and Its Associated Traits of the Ynbs Mutant in Barley

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Agronomic Trait Investigation and Data Analysis
2.3. Mapping the Gene Loci for Four Target Traits of F2 Plants
2.4. Mapping the Gene Loci for Four Target Traits of F2:5 Plant
2.5. Annotation and Screening of Candidate Genes
3. Results
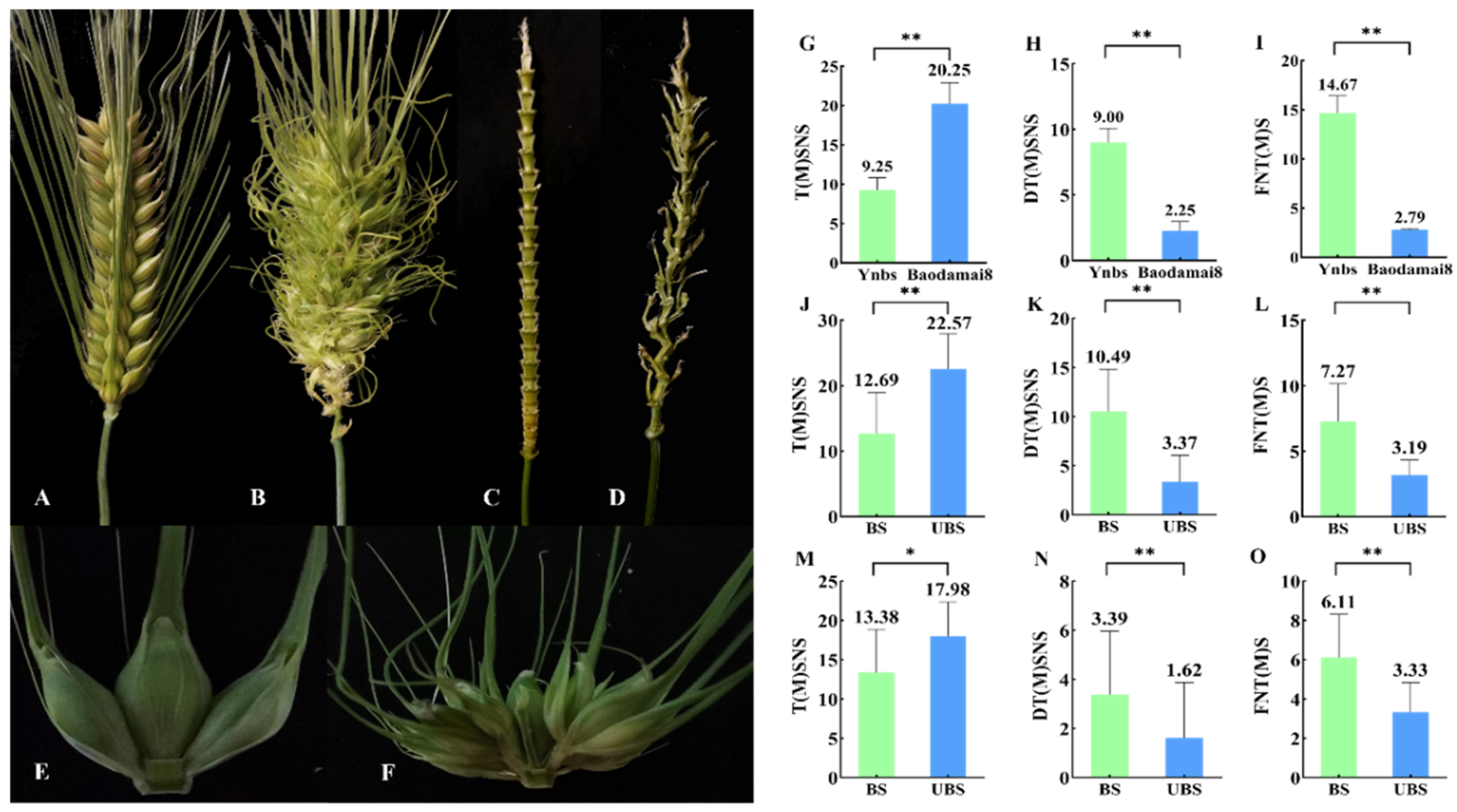
3.1. Differences in Target Trait Phenotypes between Different Materials
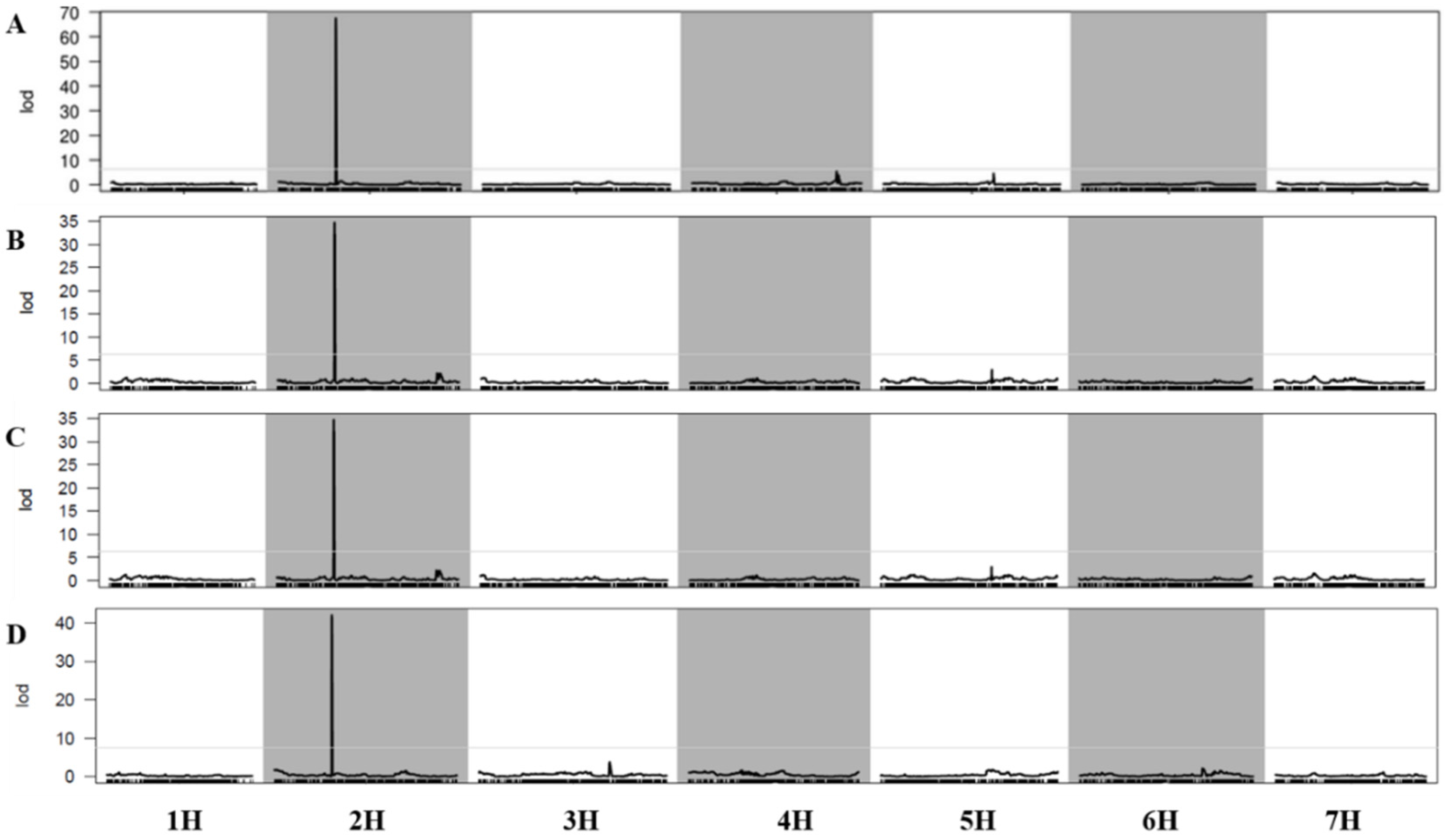
3.2. Mapping the Gene Loci for the Four Target Traits of the F2 Population
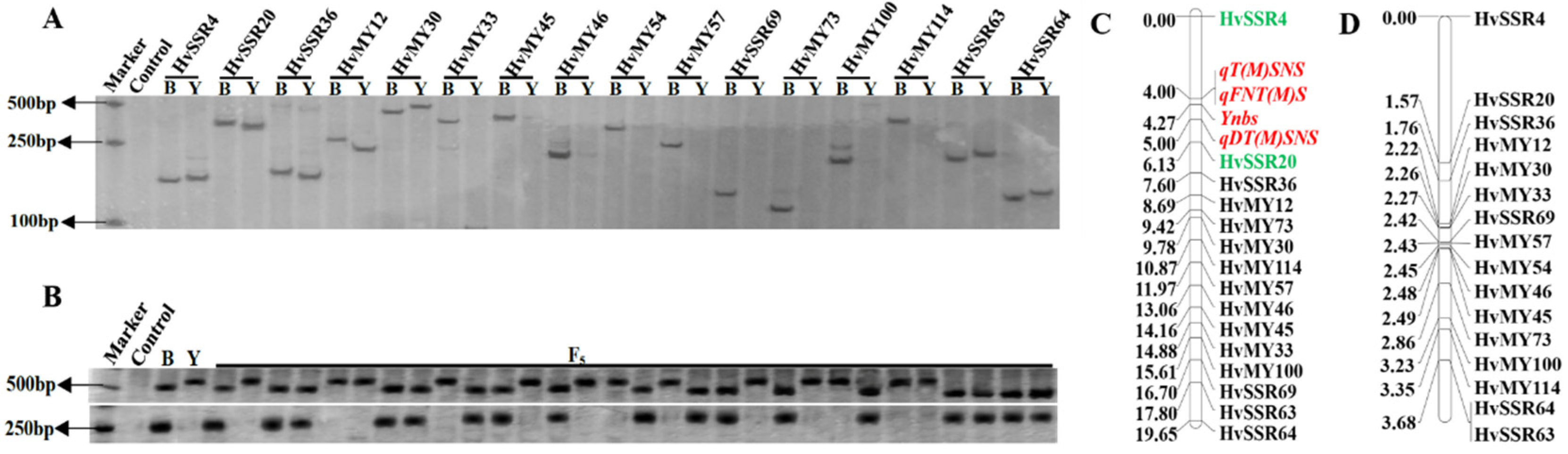
3.3. Mapping the Gene Loci for the Four Target Traits of the F2:5 Population
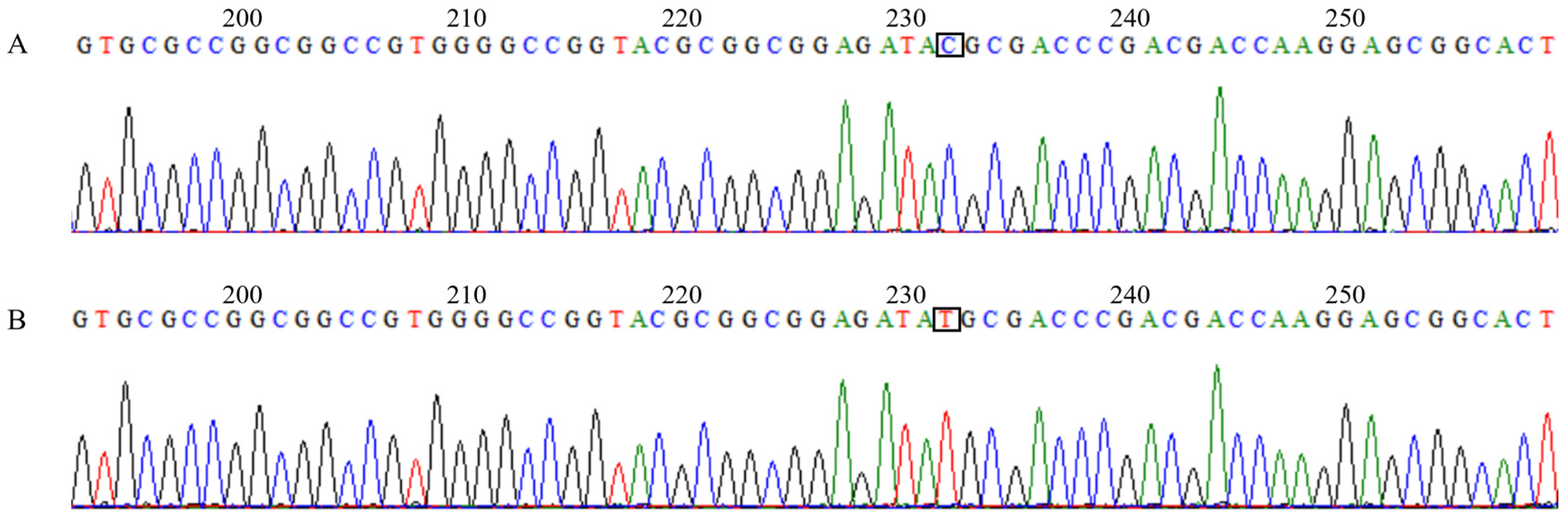
3.4. Screening of the Candidate Genes for the Four Target Traits
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harwood, W.A. An Introduction to Barley: The Crop and the Model. Methods Protoc. 2019, 1900, 1–5. [Google Scholar] [CrossRef]
- Pieper, R.; Tomé, F.; Pankin, A.; von Korff, M. FLOWERING LOCUS T4 delays flowering and decreases floret fertility in barley. J. Exp. Bot. 2021, 72, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Youssef, H.M.; Allam, M.; Boussora, F.; Himmelbach, A.; Milner, S.G.; Mascher, M.; Schnurbusch, T. Dissecting the Genetic Basis of Lateral and Central Spikelet Development and Grain Traits in Intermedium-Spike Barley (Hordeum vulgare Convar. Intermedium). Plants 2020, 9, 1655. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.; McDermott, H.I.; McAllister, T.; Cummins, M.; Silva, J.C.; Mollison, E.; Meikle, R.; Morris, J.; Hedley, P.E.; Waugh, R.; et al. APETALA2 control of barley internode elongation. Development 2019, 146, dev170373. [Google Scholar] [CrossRef]
- Zwirek, M.; Waugh, R.; McKim, S.M. Interaction between row-type genes in barley controls meristem determinacy and reveals novel routes to improved grain. N. Phytol. 2019, 221, 1950–1965. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sharma, R.; Tondelli, A.; Russell, J.; Comadran, J.; Schnaithmann, F.; Pillen, K.; Kilian, B.; Cattivelli, L.; Thomas, W.T.B.; et al. Genome-Wide Association Analysis of Grain Yield-Associated Traits in a Pan-European Barley Cultivar Collection. Plant Genome 2018, 11, 170073. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Draicchio, F.; Bull, H.; Herzig, P.; Maurer, A.; Pillen, K.; Thomas, W.T.B.; Flavell, A.J. Genome-wide association of yield traits in a nested association mapping population of barley reveals new gene diversity for future breeding. J. Exp. Bot. 2018, 69, 3811–3822. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Huang, X.Q.; Heinrichs, F.; Ganal, M.W.; Röder, M.S.; Gyenis, L.; Yun, S.; Smith, K.; Steffenson, B.; Bossolini, E.; et al. Analysis of QTLs for yield components, agronomic traits, and disease resistance in an advanced backcross population of spring barley. Genome 2006, 49, 454–466. [Google Scholar] [CrossRef]
- Ren, X.; Sun, D.; Sun, G.; Li, C.; Dong, W. Molecular detection of QTL for agronomic and quality traits in a doubled haploid barley population. Aust. J. Crop Sci. 2013, 7, 878–886. [Google Scholar] [CrossRef]
- Peighambari, S.A.; Samadi, B.Y.; Nabipour, A.; Charmet, G.; Sarrafi, A. QTL analysis for agronomic traits in a barley doubled haploids population grown in Iran. Plant Sci. 2005, 169, 1008–1013. [Google Scholar] [CrossRef]
- Wang, J.; Yang, J.; McNeil, D.L.; Zhou, M. Identification and molecular mapping of a dwarfing gene in barley (Hordeum vulgare L.) and its correlation with other agronomic traits. Euphytica 2010, 175, 331–342. [Google Scholar] [CrossRef]
- Hu, X.; Zuo, J.; Wang, J.; Liu, L.; Sun, G.; Li, C.; Ren, X.; Sun, D. Multi-locus genome-wide association studies for 14 main agronomic traits in barley. Front. Plant Sci. 2018, 9, 1683. [Google Scholar] [CrossRef]
- Shamasbi, F.V.; Jamali, S.H.; Sadeghzadeh, B.; Mandoulakani, B.A. Genetic Mapping of Quantitative Trait Loci for Yield-Affecting Traits in a Barley Doubled Haploid Population Derived from Clipper × Sahara 3771. Front. Plant Sci. 2017, 8, 688. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, X.; Heinrichs, F.; Ganal, M.W.; Roder, M.S. Analysis of QTLs for yield, yield components, and malting quality in a BC3-DH population of spring barley. Theor. Appl. Genet. 2005, 110, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, G.; Ren, X.; Li, C.; Liu, L.; Wang, Q.; Du, B.; Sun, D. QTL underlying some agronomic traits in barley detected by SNP markers. BMC Genet. 2016, 17, 103. [Google Scholar] [CrossRef] [PubMed]
- Sameri, M.; Takeda, K.; Komatsuda, T. Quantitative Trait Loci Controlling Agronomic Traits in Recombinant Inbred Lines from a Cross of Oriental- and Occidental-type Barley Cultivars. Breed. Sci. 2006, 56, 243–252. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, G.; Ren, X.; Du, B.; Cheng, Y.; Wang, Y.; Li, C.; Sun, D. Dissecting the Genetic Basis of Grain Size and Weight in Barley (Hordeum vulgare L.) by QTL and Comparative Genetic Analyses. Front. Plant Sci. 2019, 10, 469. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, X.; Yue, W.; Zhao, C.; Yang, J.; Zhou, M. Identification of QTL for barley grain size. PeerJ 2021, 9, e11287. [Google Scholar] [CrossRef]
- Koppolu, R.; Schnurbusch, T. Developmental pathways for shaping spike inflorescence architecture in barley and wheat. J. Integr. Plant Biol. 2019, 61, 278–295. [Google Scholar] [CrossRef]
- Boussora, F.; Allam, M.; Guasmi, F.; Ferchichi, A.; Rutten, T.; Hansson, M.; Youssef, H.M.; Börner, A. Spike developmental stages and ABA role in spikelet primordia abortion contribute to the final yield in barley (Hordeum vulgare L.). Bot. Stud. 2019, 60, 13. [Google Scholar] [CrossRef]
- Shang, Y.; Yuan, L.; Di, Z.; Jia, Y.; Zhang, Z.; Li, S.; Xing, L.; Qi, Z.; Wang, X.; Zhu, J.; et al. A CYC/TB1-type TCP transcription factor controls spikelet meristem identity in barley. J. Exp. Bot. 2020, 71, 7118–7131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wang, C.; Li, J.; Ma, J.; Guo, C. Identification and evaluation of Hordeum vulgare var.nudum resistance to barley yellow dwarf virus. J. Anhui Agric. Sci. 2019, 47, 152–153+158. [Google Scholar] [CrossRef]
- Shen, Z.; Li, J.; Chen, S.; Mo, F.; Li, Y.; Pu, X.; Wu, H.; Huang, W.; Luo, P. Spike characteristics and difference analysis on nine branched-spike mutants of naked kernel barley. J. Triticeae Crops 2017, 37, 66–72. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Chen, S.; Mo, F.; Li, Y.; Pu, X.; Wu, H.; Huang, W.; Luo, P. The identification of mutant traits of 9 naked barley lines with spike branch and the analysis of the association between these mutant traits. J. Yunnan Agric. Univ. (Nat. Sci.) 2017, 32, 387–394. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Chen, S.; He, J.; Zhu, S.; Mo, F.; Yu, Q.; Zhang, T.; Li, Y. Characteristics of branched-spike of hulless barley mutant-ynbs-1 and its genetic analysis. J. Triticeae Crops 2019, 39, 574–580. [Google Scholar] [CrossRef]
- Wang, W.; He, J.; Chen, S.; Peng, P.; Zhong, W.; Wang, X.; Zhang, T.; Li, Y. Construction of a high-density genetic map and fine mapping of a candidate gene locus for a novel branched-spike mutant in barley. PLoS ONE 2020, 15, e0227617. [Google Scholar] [CrossRef] [PubMed]
- Poursarebani, N.; Seidensticker, T.; Koppolu, R.; Trautewig, C.; Gawroński, P.; Bini, F.; Govind, G.; Rutten, T.; Sakuma, S.; Tagiri, A.; et al. The Genetic Basis of Composite Spike Form in Barley and ‘Miracle-Wheat’. Genetics 2015, 201, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Tanksley, S.D. Mapping polygenes. Annu. Rev. Genet. 1993, 27, 205–233. [Google Scholar] [CrossRef]
- Bai, X.; Huang, Y.; Mao, D.; Wen, M.; Zhang, L.; Xing, Y. Regulatory role of FZP in the determination of panicle branching and spikelet formation in rice. Sci. Rep. 2016, 6, srep19022. [Google Scholar] [CrossRef]
- Ren, D.; Hu, J.; Xu, Q.; Cui, Y.; Zhang, Y.; Zhou, T.; Rao, Y.; Xue, D.; Zeng, D.; Zhang, G.; et al. FZP determines grain size and sterile lemma fate in rice. J. Exp. Bot. 2018, 69, 4853–4866. [Google Scholar] [CrossRef]
- Houston, K.; McKim, S.M.; Comadran, J.; Bonar, N.; Druka, I.; Uzrek, N.; Cirillo, E.; Guzy-Wrobelska, J.; Collins, N.C.; Halpin, C.; et al. Variation in the interaction between alleles of HvAPETALA2 and microRNA172 determines the density of grains on the barley inflorescence. Proc. Natl. Acad. Sci. USA 2013, 110, 16675–16680. [Google Scholar] [CrossRef]
- Zhong, J.; van Esse, G.W.; Bi, X.; Lan, T.; Walla, A.; Sang, Q.; Franzen, R.; von Korff, M. INTERMEDIUM-M encodes an HvAP2L-H5 ortholog and is required for inflorescence indeterminacy and spikelet determinacy in barley. Proc. Natl. Acad. Sci. USA 2021, 118, e2011779118. [Google Scholar] [CrossRef]
- Shang, Y.; Zhu, J.; Hua, W.; Wang, J.; Jia, Q.; Yang, J. Characterization and mapping of a Prbs gene controlling spike development in Hordeum vulgare L. Genes Genom. 2014, 36, 275–282. [Google Scholar] [CrossRef]
- Pérez, L.; Pinazo, A.; Pons, R.; Infante, M. Gemini surfactants from natural amino acids. Adv. Colloid Interface Sci. 2014, 205, 134–155. [Google Scholar] [CrossRef]
- Feng, K.; Hou, X.; Xing, G.; Liu, J.; Duan, A.; Xu, Z.; Li, M.; Zhuang, J.; Xiong, A. Advances in AP2/ERF super-family transcription factors in plant. Crit. Rev. Biotechnol. 2020, 40, 750–776. [Google Scholar] [CrossRef]
- McSteen, P.; Laudencia-Chingcuanco, D.; Colasanti, J. A floret by any other name: Control of meristem identity in maize. Trends Plant Sci. 2000, 5, 61–66. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Annotation |
|---|---|
| HORVU.MOREX.r 3.2HG0114010 | DNA-binding pseudobarrel domain superfamily |
| HORVU.MOREX.r 3.2HG0114020 | SRC2/BAP, C2 domain |
| HORVU.MOREX.r 3.2HG0114030 | Aspartic peptidase domain superfamily |
| HORVU.MOREX.r 3.2HG0114060 | SRC2/BAP, C2 domain |
| HORVU.MOREX.r 3.2HG0114070 | Aspartic peptidase domain superfamily |
| HORVU.MOREX.r3.2HG0114100 | Aspartic peptidase domain superfamily |
| HORVU.MOREX.r3.2HG0114120 | C2 domain superfamily |
| HORVU.MOREX.r3.2HG0114130 | Sodium/calcium exchanger membrane region |
| HORVU.MOREX.r3.2HG0114150 | Homeobox-like domain superfamily; Myb family transcription factor HRS1-like |
| HORVU.MOREX.r3.2HG0114200 | Tetratricopeptide-like helical domain superfamily |
| HORVU.MOREX.r3.2HG0114210 | Zinc finger, CW-type |
| HORVU.MOREX.r3.2HG0114240 | Potassium transporter |
| HORVU.MOREX.r3.2HG0114260 | AP2/ERF domain; DNA-binding domain superfamily |
| HORVU.MOREX.r3.2HG0114270 | SRC2/BAP, C2 domain |
| HORVU.MOREX.r3.2HG0114280 | GTP-binding protein OBG, C-terminal domain superfamily |
| HORVU.MOREX.r3.2HG0114290 | Xylose isomerase |
| HORVU.MOREX.r3.2HG0114320 | DNA polymerase |
| HORVU.MOREX.r3.2HG0114330 | Ankyrin repeat-containing domain superfamily |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; He, J.; Wang, M.; Wang, X.; Chen, S. Gene Locus Mapping and Candidate Gene Screening for Branched Spike and Its Associated Traits of the Ynbs Mutant in Barley. Agriculture 2023, 13, 1934. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13101934
Zhou H, He J, Wang M, Wang X, Chen S. Gene Locus Mapping and Candidate Gene Screening for Branched Spike and Its Associated Traits of the Ynbs Mutant in Barley. Agriculture. 2023; 13(10):1934. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13101934
Chicago/Turabian StyleZhou, Hongbin, Junyu He, Mengyue Wang, Xintian Wang, and Shengwei Chen. 2023. "Gene Locus Mapping and Candidate Gene Screening for Branched Spike and Its Associated Traits of the Ynbs Mutant in Barley" Agriculture 13, no. 10: 1934. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture13101934