Impact of Colletotrichum acutatum Pathogen on Olive Phenylpropanoid Metabolism
by
 , , and
, , and
Irene Gouvinhas
1 ,
,
Paula Martins-Lopes
2,3,*,
Teresa Carvalho
4,
Ana Barros
1 and
and
Sónia Gomes
2,3,* 1
Centre for the Research and Technology of Agro-Environmental and Biological Sciences (CITAB), University of Trás-os-Montes and Alto Douro, 5000-901 Vila Real, Portugal
2
School of Life and Environment Sciences, University of Trás-os-Montes and Alto Douro, 5000-901 Vila Real, Portugal
3
BioISI–Biosystems & Integrative Sciences Institute, Faculty of Sciences, University of Lisboa, 1749-016 Lisboa, Portugal
4
National Institute for Agricultural and Veterinary Research, I. P. (INIAV), Estrada de Gil Vaz, 7351-901 Elvas, Portugal
*
Authors to whom correspondence should be addressed.
Agriculture 2019, 9(8), 173; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9080173
Submission received: 28 June 2019
/
Revised: 4 August 2019
/
Accepted: 5 August 2019
/
Published: 6 August 2019
(This article belongs to the Special Issue Plant-Microbe Interactions)
Abstract
:Olive anthracnose caused by the hemibiotrophic fungal pathogen Colletotrichum acutatum is a serious threat to the olive sector. Olive oil and fruit production is severely constrained by Colletotrichum spp. infection, being C. acutatum the most distributed pathogen in Portuguese olive orchards. To understand the impact of C. acutatum on phenylpropanoids biosynthesis, the enzyme activity, phenolic compounds, ortho-diphenols, and flavonoids content were determined and correlated with the expression of gene encoding key enzymes within phenylpropanoids metabolism in susceptible and tolerant olive fruits, during maturation and when infected with C. acutatum. Differences between cultivars was observed, the tolerant olive cv. Picual presented a higher basal value and a stable phenolic content throughout the infection process, supporting its high C. acutatum tolerance, whereas in the susceptible olive cv. Galega these secondary metabolites were significantly increased only after the elicitation with C. acutatum.
1. Introduction
Olea europaea L. is one of the most ancient fruit crops of the Mediterranean area. The olive sector is prevalently directed towards olive oil production. However, due to widespread, climatic changes and to the intensive production, several diseases have aroused compromising olive yield/quality. Among the several biotic constraints affecting olive production, the anthracnose disease caused by Colletotrichum spp. is one of the most damaging olive diseases [1,2,3]. Among the Colletotrichum spp., C. acutatum, a hemibiotrophic fungus, is a major production-limiting factor since it affects both the fruit yield and the olive oil quality (e.g., off₋flavor, reddish color, high acidity, and reduction in polyphenolic content) [2,4,5,6].
The disease incidence and severity has increased in the last years, being largely reliant on climatic conditions, cultivar susceptibility, and fungal isolates virulence [1,7]. During the last decades, this disease has spread from the initial affected olive growing region of Alentejo, where it mainly attacked the susceptible cv. Galega, to all the country and diverging in other cultivars [8,9,10,11]. The cv. Galega is a very important, economic and traditional Portuguese cultivar, giving rise to unique olive oils with differentiated flavors. Since cv. Galega is highly affected by C. acutatum it has been replaced by other olive cultivars with more tolerance to C. acutatum, such as foreign cvs. Picual and Arbequina, having direct implication on olive oil Portuguese denomination. Specific parameters led to the creation of a Protected Designation of Origin (PDO) of olive oil, being cultivar composition considered to be of major importance, influencing olive oil quality and sensory characteristics [12]. Portugal has six PDO denomination for olive oil and cv. Galega is present in five of the PDO regions.
Considering cultivars behavior to C. acutatum infection, several reports have indicated cvs. Picual, Cobrançosa, and recently Arbequina, as tolerant to C. acutatum [9,13,14]. Understanding the pathogen–cultivar interaction during the infection process is a major issue that needs to be achieved since it has a direct implication on yield and olive oil quality [6,11]. The cultivar–pathogen interaction and immunity are based on an array of defensive mechanisms such as: (1) physical barriers (e.g., cell wall, fruit cuticle, needles, and trichomes); (2) pathogenesis-related proteins (PR); and (3) secondary metabolites biosynthesis, induced during host–pathogen interaction [15,16,17,18]. The systemic induced defense responses are activated when the pathogen is in contact with the host, resulting in: (1) cell-wall thickening; (2) lignification; (3) papilla formation; and (4) secondary metabolites synthesis that derive from multiple branches of the phenylpropanoid pathway (e.g., flavonoids, phytoalexins, and other phenolic compounds) creating a hostile and toxic environment for pathogen growth [15,16,19,20,21,22,23,24,25,26]. Among the modular enzymes associated with plant secondary metabolites, phenylalanine ammonia-lyase (PAL) is one of the key enzymes in the plant defense response, mediating the oxidative burst that leads to cell death in the hypersensitive response [27,28]. PAL is a precursor of a large number of secondary metabolites (e.g., flavones, flavonols and isoflavonoid-phytoalexins, proanthocyanidins, among others) [29,30], which are accumulated in plants when submitted to stress conditions such as, when submitted to a pathogen attack as a defense mechanism [31,32,33,34,35]. Different reports on diverse fruit crops have referred significant changes in the global metabolite profiles of infected fruits [36]. During fruit development, the mesocarp accumulates a wide range of secondary metabolites, some of them having an important role in the plant’s defense mechanisms reducing the infection progression [21,32,34,37].
This work aims to understand the olive-C. acutatum interaction in the context of the phenylpropanoid pathway. In this sense, the phenolic composition, as well as the phenylalanine ammonia-lyase activity, were studied, both in cultivars susceptible and tolerant to C. acutatum infection, and during fruit maturation. Correspondingly, OePAL gene expression was accessed aiming to correlate its expression with phenolics biosynthesis (e.g., ortho-diphenols and flavonoids), which proved to be critical for the defense response. Our findings revealed that there is a close correlation between the genotype and the infection process and that the phenylpropanoid metabolism plays a key role in this pathosystem.
2. Materials and Methods
2.1. Plant Materials
Olive samples from three olive cultivars with different C. acutatum susceptibility (‘Galega Vulgar’—susceptible, ‘Cobrançosa’—moderately–tolerant; and ‘Picual’—tolerant), were harvested from a certified olive grove, at the National Institute for Agricultural and Veterinary Research (INIAV, Elvas, Portugal) (38°54′53.35″ N and 7°19′11.06″ O) (Table 1) during the crop seasons 2016 and 2017. For the maturation study, healthy olive fruits, without any kind of infection or physical damage, were collected from ten different trees of comparable age and vigour, evenly spaced between them, within the same growing area. More, all the olive trees of the same cultivar resulted from a clonal propagation, and were further checked using Single Sequence Repeats (SSR) markers to confirm their same genetic basis and cultivar denomination [9]. Olives were handpicked at three ripening stages (green, semi-ripe, and ripe). The harvesting dates and the ripening index (RI) are showed in Table 1. For the assessment of the RI, the harvested olives were evaluated according to the skin and pulp color, following the criteria previously described in the literature [38]. The RI values considered ranged from 0 (100% intense green skin) to 7 (100% purple flesh and black skin).
2.2. Preparation of C. acutatum Isolates, Plant Inoculation and Sampling
C. acutatum isolates were collected from diseased fruits (tolerant cv. Picual) as previously described by Gomes et al. [39]. All isolates were identified by the Plate Trapped Antigen-Enzyme Linked Immunosorbent Assay (PTA-ELISA) test using specific antibodies for C. acutatum, and monoconidial isolates were prepared to be used for the field inoculation experiments [8]. The isolates were cultured on potato dextrose agar (PDA) for 8 days at 22–24 °C under a 12-h photoperiod. Inoculum was prepared by flooding dishes with sterile distilled water, scraping the surface gently with a glass rod, and filtering the resulting suspension through sterile cheesecloth. A conidial suspension was determined by counting a solution of 2 × 106 conidia per mL and used to spray the olive trees. The same inoculum was used in the two consecutive years.
All the fruits inoculation experiments were conducted under field conditions and during two consecutive seasons (2016 and 2017), at a ripening index of 3. Two branches (approximately with 100 fruits) in each olive tree were inoculated with the conidial suspension to evaluate the disease symptoms. After inoculation, plants were enclosed in plastic bags, during 48 h, to create high humidity conditions in order to encourage spore germination and fungus development. All samples were collected at 0, 16, 48, and 144 h after infection (hai), frozen in liquid nitrogen and stored at –80 °C accordingly to the experimental procedure. Non-infected samples (control) w sprayed with sterile water and collected (Table 1). All analyses were performed in 12 olive trees/per cultivar.
2.3. Real-Time Polymerase Chain Reaction (PCR) Assay
2.3.1. Total RNA Isolation
Total RNA was isolated from frozen olive fruits with a Qiagen RNeasy Plant Mini kit (Qiagen, CA, USA) following the manufacturer’s instructions. After a column DNase treatment, the RNA quantity and quality were estimated by Nanodrop™ 1000 Spectrophotometer (Thermo Fisher Scientifc, Wilmington, DE, USA) measurements and by electrophoresis on a 0.8% agarose in 1× TAE (Tris-acetate-EDTA). The cDNA library preparation was synthesized from 5 µg of DNA-free RNA using a High-Capacity cDNA Reverse Transcription kit (Thermo Fisher Scientific, Porto, Portugal) according to the manufacturer’s instructions. The cDNA was adjusted to 100 ng µL−1 concentration and kept at −20 °C. The gene full-length, putatively encoding phenylalanine ammonia-lyase (PAL) was cloned from olive using degenerate primers. The olive PAL sequence was used to design specific olive primers to be used in the real-time PCR assay. The expression of the phenylpropanoid OePAL gene was performed in the three genotypes using C. acutatum non-infected (control) and infected samples (0, 16, 48, 144 hai).
2.3.2. Reverse Transcription−Quantitative Real-Time Polymerase Chain Reaction (qRT–pCR)
Reverse transcription−quantitative real-time PCR (qRT–pCR) was performed using the SYBR Green PCR Master Mix Kit (Thermo Fisher Scientific, Portugal) in an Applied Biosystems Real–Time PCR System (Thermo Fisher Scientific, Portugal). The cycle threshold (Ct) values were used for expression level analysis. Primer sequences of O. europaea actin (GenBank ID: AF545569), phenylalanine ammonia-lyase (GenBank ID: KJ511868), Glyceraldehyde 3-phosphate dehydrogenase (GenBank ID: EF506494) were designed based on the O. europaea nucleotide sequence for each deposited gene available at the public database (GenBank). All primers were designed ranging from 19 to 22 bp in length and were synthesized by STAB Vida, Portugal. Specific primers used to study the expression of O. europaea fruits under C. acutatum infection, as well as the accession numbers of the nucleotide sequences on which primer design was based, are listed in Table 2. All experiments were performed in triplicate (biological replicates and assays) and were analyzed using StepOne v2.2.2 Software (Life Technologies, Portugal), using both OeGAPDH and OeActin as internal controls. PCR amplifications were carried out in total volume of 20 µL, containing 10 µL of Power SYBR Green for Master Mix, 0.5 µM of each primer and 20 ng of cDNA. Amplification conditions for OePAL consisted on an initial denaturation of 10 min at 95 °C, followed by 40 cycles consisting of denaturing at 95 °C for 15 s, annealing at 59 °C for 1 min, and an extension at 72 °C for 45 s. After each run, a melt curve was generated to check for the existence of unspecific amplicons and primer–dimer generation. Non-template controls were included in each reaction. The real–time PCR data were analysed using StepOne v2.2.2 Software (Applied Biosystems, Portugal). Cycle threshold (Ct) values were obtained with the same software and the results were normalized to the OeActin and GAPDH expression levels and analyzed using the 2−ΔΔCT method [40].
2.4. Quantification of Phenylpropanoid Compounds
2.4.1. Extraction and L-Phenylalanine Ammonia-Lyase (PAL) Activity Assay
PAL enzyme activity was determined according Tovar et al. [35]. For every sample analyzed, 10 fresh fruits were first frozen at −80 °C and lyophilized. One gram of olive flesh powder was weighed into a beaker adding 25 mL of 0.05 M potassium phosphate buffer (pH 6.6). The mixture was homogenized and 0.2 g of Triton X–100 (Sigma-Aldrich, Steinheim, Germany) and 25 mg of insoluble polyvinylpyrrolidone (PVP, Sigma-Aldrich, Steinheim, Germany) were added. The suspension was maintained at 4 °C during extraction and centrifuged during 15 min at 10,000× g. For PAL activity quantification, 0.4 mL of olive flesh homogenate and 4.1 mL of sodium phosphate buffer were mixed in test tubes. The reaction was initiated when 1 mL of L-phenylalanine (10 mg/mL) was added to the solutions in the test tubes. The tubes were then heated for a period of 1 h at 37 °C. The reaction ended after adding 0.5 mL of 35% trifluoroacetic acid (w/w) (Sigma–Aldrich, Steinheim, Germany). Tubes were centrifuged during 5 min at 10,000× g. The PAL quantification was performed by measuring the absorbance at 290 nm according to McCallum et al. [41] using a spectrophotometer Genesys 2 PC (Spectronic Instruments, Leeds, UK). The enzymatic activity is expressed in μmol cinnamic acid liberated h−1 g−1 of dry weight. To confirm that the change observed in the absorbance was due to the formation of trans–cinnamic acid, the enzymatic reaction was followed over a 3 to 6 h period by Reversed-Phase High-Performance Liquid Chromatography coupled with Diode Array Detection (RP–HPLC–DAD), using the method described by Machado et al. [42]. A reaction without the substrate was used as a blank control. Triplicate assays were performed for each extract.
2.4.2. Phenolic Fraction Extraction
Olive samples (5 g) were macerated with 50 mL of a mixture of MeOH/H2O (50:50) and incubated at room temperature for 30 min. The supernatant was decanted, and the extractions were repeated three times. The liquid phase was filtered and subsequently centrifuged at 9000× g for 10 min. To remove the fat phase, the mixture was washed twice with hexane (50 mL) using a separating funnel and the organic phase was discarded. Three replicates per sample were performed [30,43].
2.4.3. Phenolic Composition
The content in total phenols, flavonoids, and ortho-diphenols was determined according to spectrophotometric methodologies previously reported by Gouvinhas et al. [43], with minor modifications. The total phenolic compounds content was determined using Folin–Ciocalteu (Panreac Química S.L.U., Barcelona, Spain), with 3,4,5-trihydroxybenzoic acid (gallic acid) extra pure (>99%) (Panreac Química S.L.U., Barcelona, Spain) as standard [44]. Briefly, 1 mL of diluted sample was mixed with 500 µL of Folin–Ciocalteu reagent, 2 mL of 7.5% sodium carbonate solution and 6.5 mL of MilliQ water (Millipore, USA). The mixture was homogenized and the absorbance of the standard solutions and samples was measured at 750 nm after 30 min reaction at 70 °C in relation to a gallic acid standard curve. All measurements were performed in triplicate. The results were expressed as milligrams of gallic acid per gram of dry weight (mg GA g−1 DW). The content of ortho-diphenols in olives was determined by adding 1 mL of Na2MoO4 (50 g L−1) to 4.0 mL of the samples appropriately diluted. Mixtures were vortexed and allowed to rest at room temperature, protected from light, for 15 min. The absorbance was recorded at 375 nm and quantified using gallic acid as standard. Results were expressed in mg GA g−1 DW. For the assessment of flavonoid content, 0.5 mL of the sample properly diluted were mixed with 150 μL of NaNO2 (50 g L−1). After 5 min precisely, 150 μL AlCl3 (100 g L−1) were added and the mixture was allowed to react for 6 min. Subsequently, 1.0 mL of NaOH (1 M) were added to the mixture. The absorbance was immediately recorded at 510 nm, and the flavonoid content quantified using catechin as standard. Results were expressed in mg of catechin per gram of dry weight (mg CAT g−1 DW).
2.5. Statistical Analysis
The results are presented as mean values ± standard deviation (n = 3). Statistical analysis was performed using one-way analysis of variance (ANOVA), and a multiple range test (Turkey’s test), with a significance level of p < 0.05, using IBM SPSS statistics 21.0 software (SPSS Inc., Chicago, IL, USA), in order to determine the significant differences of each assays among the different fruit stages (maturation/infection). Different letters are significantly different for each cultivar.
3. Results
3.1. OePAL Gene Transcript Levels during Fruit Development and C. acutatum Infection
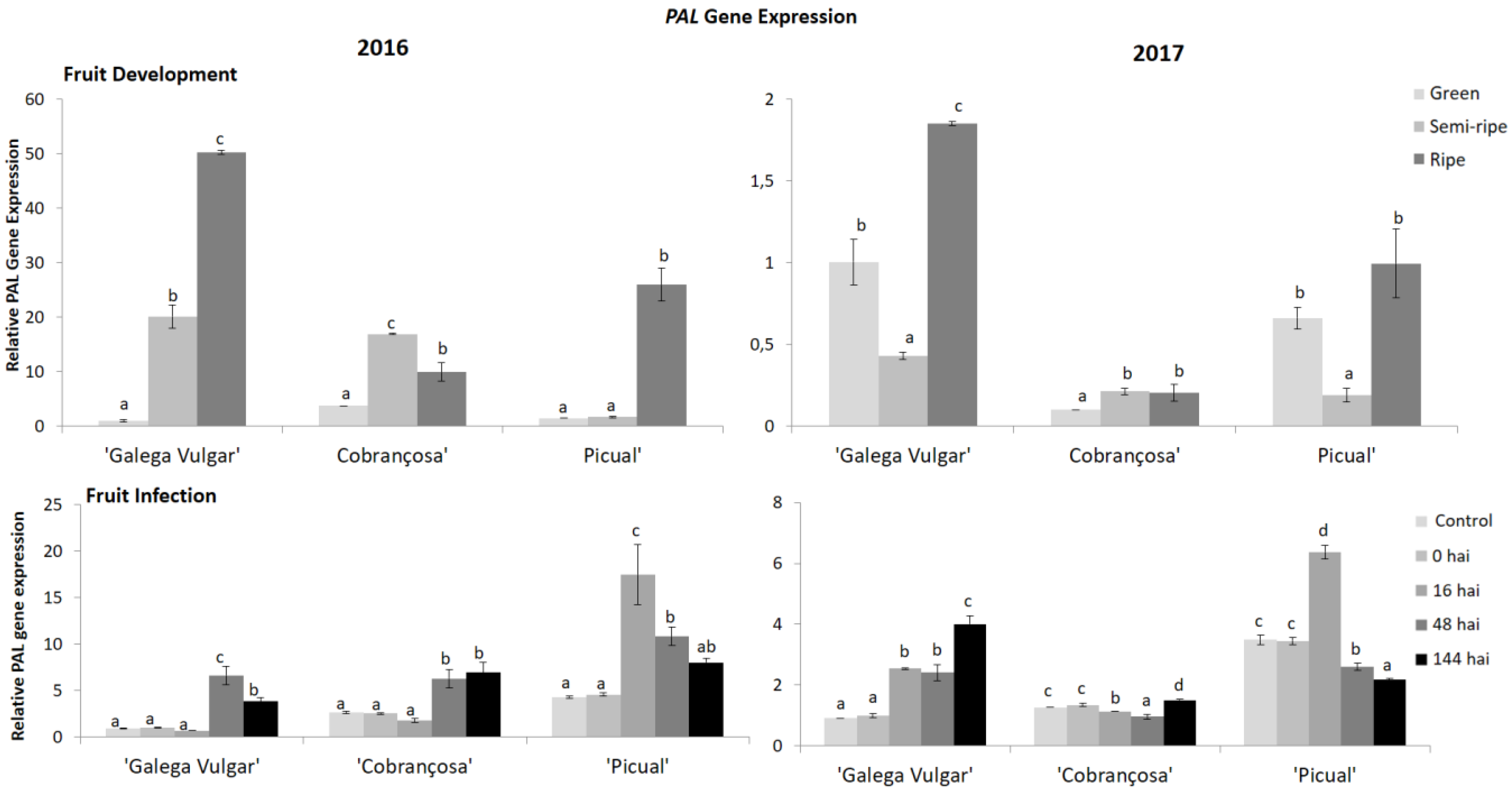
The influence of both cultivar (genotype) and fruit development on disease progression was achieved by phenylpropanoid metabolism changes, considering: (1) OePAL expression by real-time polymerase chain reaction (PCR) (Figure 1); (2) PAL enzyme activity (Figure 2), and (3) fruit phenolic composition (Figure 3, Figure 4 and Figure 5). The levels of OePAL gene transcripts observed in fruit ripening period were consistent with significant increase of expression from the green to the ripening stage (p < 0.05) (Figure 1) with some variations between these two phases. Indeed, in the first crop year (2016), all olive cvs. presented the lowest relative expression in the green stage, with a significant increase for the first two cultivars (Figure 1). While cv. Galega showed a significant increase of transcript accumulation until the last ripening stage, cv. Cobrançosa presented a decrease of PAL expression. Picual olive fruit samples didn’t show differences from the green to the semi-ripe stage, however it presented an important and significant increase in the last stage. Galega was the cv. that presented the highest transcript accumulation at ripe stage, followed by cvs. Picual and Cobrançosa. In the second-year crop (2017), while cv. Cobrançosa showed a slightly variation between all stages of fruit maturation, cvs. Galega and Picual presented the same behaviour, once from the green to the semi-ripe stage a significant decrease of PAL expression was observed, and then a significant increase from this one to the ripe stage was found in both samples. The susceptible cv. Galega showed the highest relative expression of PAL gene.
During C. acutatum infection, the PAL transcript abundance was higher in fruits belonging to the tolerant cv. Picual (Figure 1). The PAL relative expression was different among crop years, presenting an up-regulation at 48 hai in 2016, being anticipated to 16 hai in 2017. Comparing the different olive cvs. it is evident that cv. Picual consistently presented the highest PAL expression over the years. The Figure 1 also showed that Picual and Cobrançosa cvs. presented a higher basal expression level in comparison to cv. Galega. Nevertheless, the overall expression increase of cv. Galega was of 6-folds, whereas cv. Cobrançosa presented an increase of 3-fold in the field year of 2016. In the crop year 2017 cv. Galega and cv. Picual presented a similar behaviour in comparison to crop year 2016, whereas cv. Cobrançosa only had a slight change in the expression profile throughout the elicitation. Overall, as observed for the maturation study the OePAL expression was significantly lower in crop year 2017. Nevertheless, the PAL expression levels observed in 2017, after fungus elicitation, was higher than those observed for the maturation studies, indicating that this gene is up-regulated when exposed to this fungus.
3.2. PAL Enzyme Activity during Fruit Development and upon C. acutatum Infection
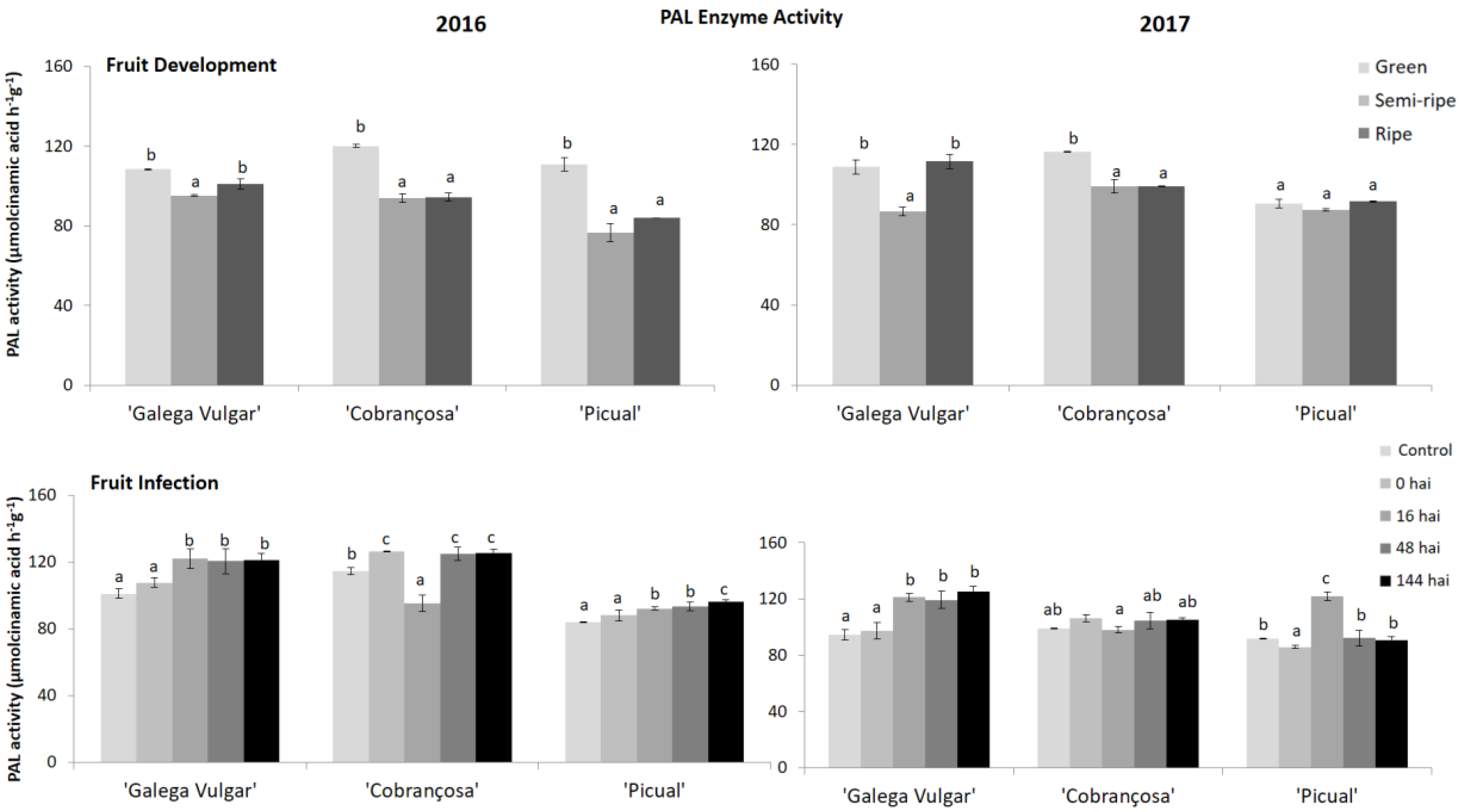
The enzyme activity in fruits from susceptible and tolerant cvs. was evaluated colorimetrically during fruit ripening, as well as in all time points of fruits C. acutatum infection (0, 16, 48, 144 hai) (Figure 2). Regarding the fruit ripening, the PAL enzyme activity was significantly higher in green fruits, in both crop years, except for Picual olive fruits that presented no significant differences between fruit ripeness. Concerning the other olive fruit samples, a significant decrease was observed in the enzyme activity, from the green to the semi-ripe fruit stage. However, no significant differences were found between the middle and the last stage of maturation, except in the crop year 2016–2017 for susceptible cv. Galega, since PAL activity reached approximately the same activity level as the one observed in the green stage (108.85 ± 3.51 and 111.66 ± 3.56 µmol cinnamic acid h−1 g−1, for green and ripe stage, respectively).
In susceptible and tolerant, subjected to pathogen infection, different enzyme activity responses were observed (Figure 2). Fruits from susceptible cv. Galega presented an increase in PAL enzyme activity until 16 hai (from 101.0 to 122.1 µmol cinnamic acid h−1 g−1 in the 2016 crop year, and from 111.7 to 121.1 µmol cinnamic acid h−1 g−1 in the 2017), followed by a constant enzyme activity during the fruit infection, presenting no significant activity (p ≥ 0.05) in the last hours of infection. In tolerant fruits (cv. Picual) the enzyme activity levels did not change significantly in the crop year 2016, whereas in the second year, 2017, the enzyme activity increased significantly at 16 hai (from 85.9 to 122.0 µmol cinnamic acid h−1 g−1) compared to the control. Fruits from cv. Cobrançosa displayed the same enzyme activity in both years. This cv. presented a significant decrease in enzyme activity (p < 0.05) at 16 hai compared to the control (from 126.2 to 95.1 µmol cinnamic acid h−1 g−1). Afterward, the enzyme activity increased continuously, reaching a similar level as that observed in the control samples (Figure 2). Overall, the tolerant cv. Picual presented the lowest enzyme activity values. Although the PAL activity is induced after C. acutatum elicitation, its activity is higher in susceptible genotypes compared to the moderate and tolerant ones.
3.3. Evaluation of Phenolics, Ortho-Diphenols, and Flavonoids during Olive Fruits Development and C. acutatum Infection
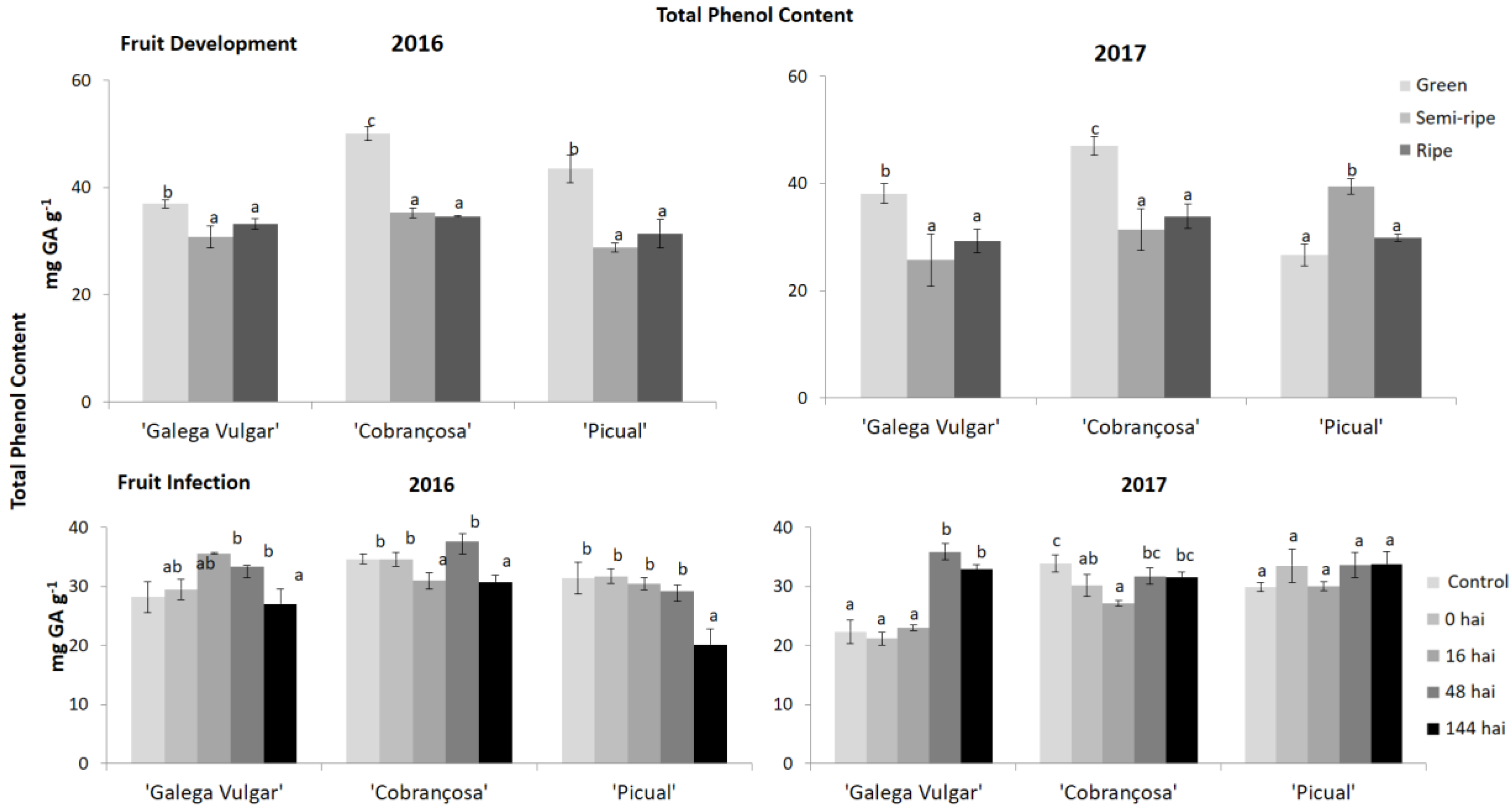
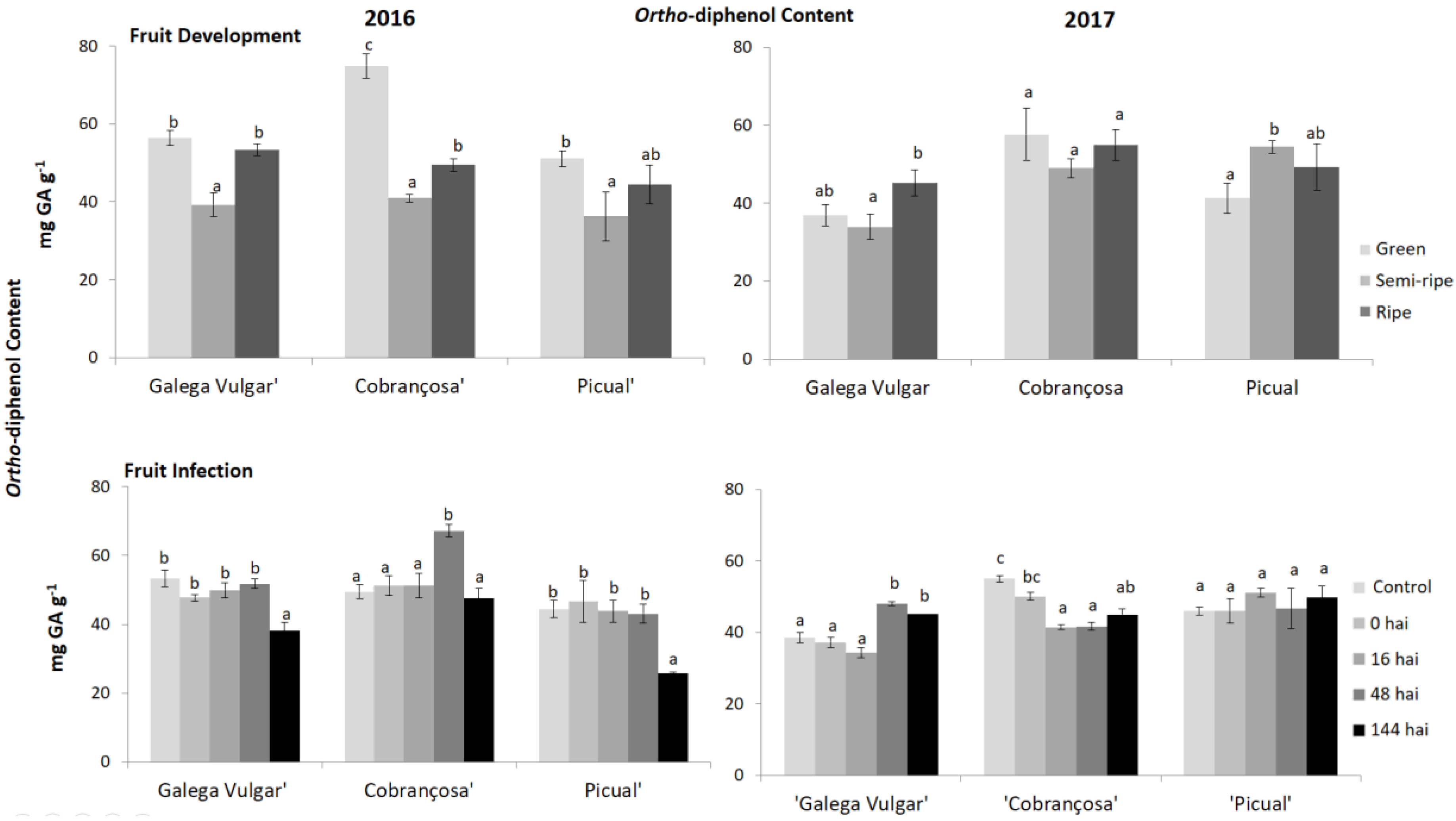
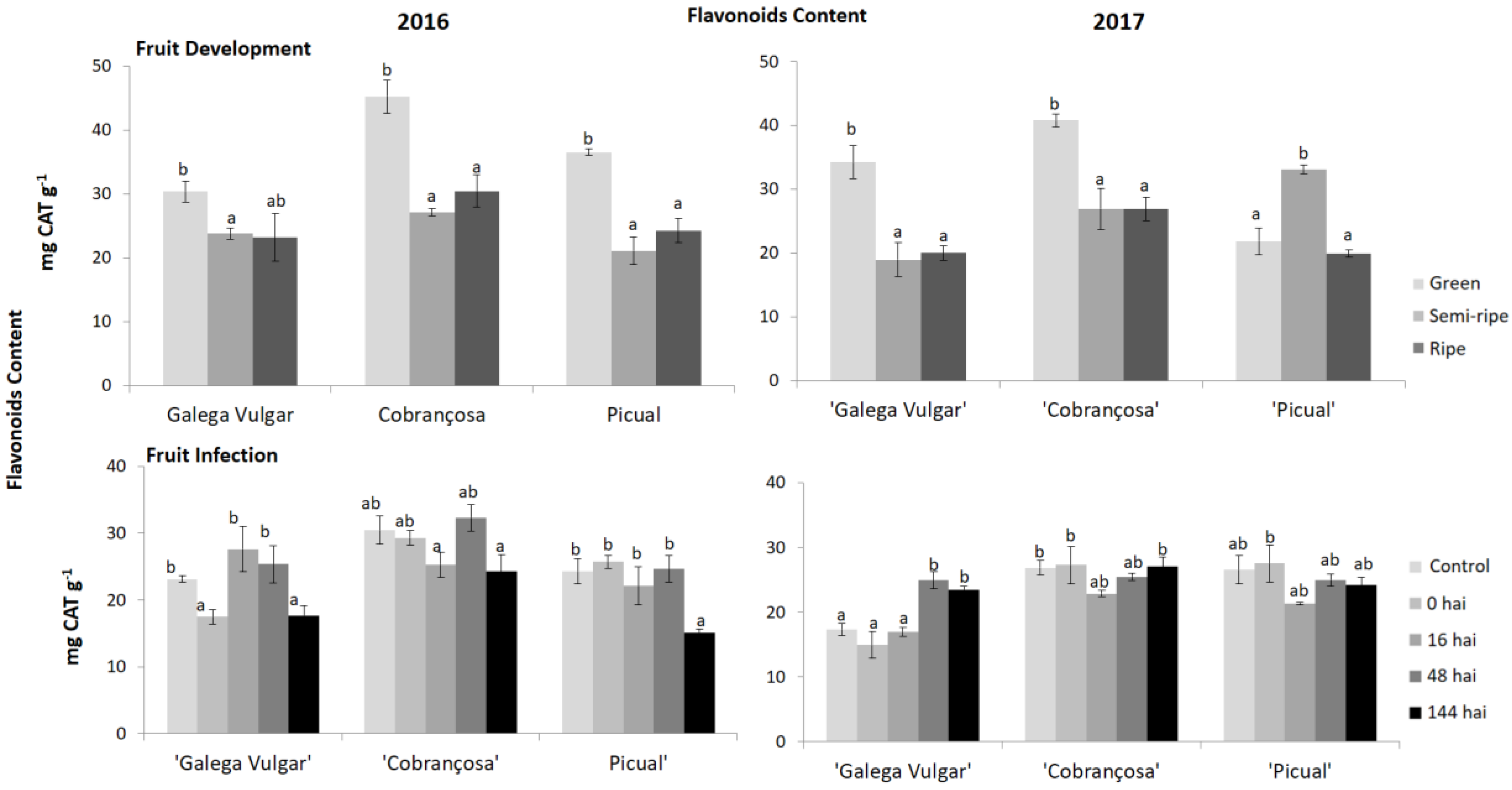
The phenolic composition of olive fruits from the three cultivars studied during the maturation process and fungus infection was evaluated (Figure 3). The behaviour of the olive samples during maturation is very similar between crop years, with a significant decrease of total phenols, ortho-diphenols (Figure 4), and flavonoids (Figure 5) in 2016 from green to semi-ripe stage, with a slight increase until the ripe stage, not reaching the values of the green phase. This was also observed in the second crop season (2017), except for cv. Picual that presented a significant increase of these parameters, from the green to the semi-ripe, followed by a significant decrease, reaching similar values to the other two olive cultivars.
Regarding the fruit infection by C. acutatum, the phenolic compounds in cv. Cobrançosa revealed, generally, a slight decrease at 16 hai compared to the control, followed by an increase until 48 hai (Figure 3). Tolerant cv. Picual did not present a significant (p ≥ 0.05) decrease in the pre-existing phenolics in the second crop year, however, in 2016, a significant decrease of total phenols, ortho-diphenols (Figure 4) and flavonoids (Figure 5) was observed at 144 hai.
The basal level of phenolic compounds was higher in moderate and tolerant olive fruits, suggesting a pre-existing antifungal barrier that acts as defense against pathogen proliferation in olive. Fruit samples, from susceptible cv. Galega, presented the lowest phenolics basal content (29.5 and 21.2 mg GA g−1, in 2016 and 2017, respectively) compared to the tolerant genotypes (31.74 and 33.50 mg GA g−1, in 2016 and 2017, respectively). Furthermore, the phenolic composition in susceptible fruits is higher at 16 and 48 hai.
4. Discussion
The secondary metabolites have an important role in the plant defense against fungal pathogen on different crops. Olives infected with C. acutatum are reported to have both substantial yield losses, as well as a considerable impact on fruit sensorial and chemical changes [11]. PAL is known to play an important role in the phenylpropanoid pathway being involved in the biosynthesis of the polyphenol and secondary metabolites. In this pathosystem the clear relationship between PAL and biosynthesis of secondary compounds, from phenylpropanoid metabolism, has not been previously reported. The high activity levels of PAL enzyme, observed in fruit samples at the early stage of maturation, were also reported by Morelló [45] in Arbequina, Farga and Morrut, and by Ortega-García [46] in Picual, Verdial and Frantoio, indicating an active participation of the phenylpropanoid pathway in the secondary metabolism of olive fruits, as well as an enlargement of all parts of the ovary and lignification of the endocarp; slightly correlated to the total phenol content in the olive pulp as described by Jimenez-Ruiz [14]. The decline of the PAL activity over the maturation agrees with the previous report by Tovar [35] that related the lower PAL activity with the decrease of the phenolic compounds. Morelló [45] also reported an important decrease of PAL activity at the beginning of the ripening process, when the olive fruit color turns from green to light green or with the appearance of red spots on the skin, giving rise to the beginning of black maturation. In cv. Galega which presented an increase on PAL activity in the last maturation stage, this raise can be explained by a higher accumulation of anthocyanins, and consequently the change of fruit color to purple/black, since this cv. reaches complete maturation previously to the other cvs under study, requiring a more active metabolism to achieve it in a short period. In strawberries, two peaks of PAL activity are also observed, the first one due to the synthesis of flavonoids and phenolic compounds, which occurs in the green stage, and the second peak in ripe fruits due to the anthocyanin’s accumulation [47,48].
Considering the gene expression pattern during fruit infection (Figure 1), our results were consistent with the microscopic data reported by Gomes et al. [9], where fruit mesocarp from susceptible cv. Galega present infection symptoms as early as 48 hai while tolerant cv. Picual presented infection symptoms, on fruit mesocarp, only after 72 hai. The fruit ripeness increases the fruit susceptibility to pathogen infection, with the unripened (or developing) fruit being tolerant to the C. acutatum infection [9,13]. According to the total phenolic measures (Figure 3), the cultivars respond to the infection by increasing the level of phenols at 16 hai (crop year 2016) and 48 hai (crop year 2017) directly associated with the elicitation increases of the PAL enzyme activity (Figure 2).
Recently, Gouvinhas et al. [49] measured the concentration of individual phenolic compounds in the olive cvs during C. acutatum infection and these individual compounds (e.g., hydroxytyrosol, hydroxybenzoic acid, verbascoside, luteolin, oleuropein and cinnamic acid) vary significantly among susceptible and tolerant cvs. Although total phenolics are in higher concentration in cv. Picual (27.77–33.80 mg GA g−1) in comparison to cv. Galega (21.19–24.67 mg GA g−1) the most remarkable difference is observed at the secoiridoid glucosides (oleuropein) level, where cv. Picual presents a higher concentration, supporting the idea that the resistance might also be related to the presence of certain types of phenolic compounds more than the amount of each of them [50,51,52]. One of the most important secoiridoids present in olive fruit and virgin olive oil is oleuropein, the composition of which varies considerably among cultivars, tissues, developmental stages and in response to different biotic stresses. It is also responsible for phytoalexins release. The cv. Galega was characterized by having low oleuropein concentration at 16 hai (4.8% relative abundance) in contrast to tolerant cv. Picual (14.5%), which ranged between 14.5% and 29.9% [53,54]. Curvers et al. [54] and Bassolino et al. [55] reported that plant cell wall has some composition modifications through the incorporation of phenolic compounds, which were considered an important defense mechanism against B. cinerea. In fact, these compounds play a crucial role in the plant response to environmental cues, being the most important defense compounds against biotic stress. Moreover, the observed differences in the individual phenolic compounds, among susceptible and tolerant cultivars, reflect the great variability in the modulation of their biosynthesis and accumulation [29]. Furthermore, the different fruit composition in ortho-diphenols (Figure 4) and flavonoids (Figure 5) observed during maturation in susceptible and tolerant genotypes could explain the difference of resistance among olive cultivars. The increasing levels of phenolics after an elicited activity of key enzymes, such as PAL, has been reported as a component of plant systemic immunity to overcome the infection process [56]. PAL activity can also act as a first line of defense against C. acutatum infection, since the phenolic compounds produced might inhibit the pathogens’ progression [15,26,57]. Thus, the phenolic composition of the olive fruits can be used as an indicator of the susceptibility/tolerance level of plants to diseases and pests attack [58]. Additionally, even though the physical barriers have not been considered in the present study, a previous work has demonstrated that the olive fruit cuticle thickness is higher in tolerant cv. Picual in comparison to susceptible fruits’ cuticles [16]. The response of olive fruits to C. acutatum is similar as reported in strawberry (Fragaria × ananassa Duchesne) [21]. Prusky et al. [59] also reported that the synthesized products of the phenylpropanoid pathway (i.e., phenolics and flavonoids) in avocado (Persea Americana Mill.) create a hostile and toxic environment for C. gloesporioides growth. A similar study in apple (Malus domestica Borkh.) infected by scab (Venturia inaequalis Cke.) revealed that the flavonoid biosynthesis limited the scab progression within the apple cells [60,61]. In line with these findings, Slatnar et al. [56] reported higher PAL enzyme activity, in scab spot tissue of infected apples. In onion (Allium cepa L.) the high levels of phenolic compounds have also been related to the tolerance to Colletotrichum circinans [62].
In this study, fruits from tolerant cv. Picual presented endogenously the highest phenolic content; whereas the fruit samples from cv. Galega had an immediate response to the fungal attack, with an induced increase of phenolics. These responses of susceptible genotypes were expected, since phenolic compounds are known to actively participate in plant defense mechanisms, and their content in cell/tissues is significantly increased when submitted to different stress conditions. Furthermore, the low pre-existing phenolic content, observed in control fruit samples from susceptible cv. Galega, is also in accordance with the literature, since the tolerant cv. Picual, naturally overcame the development of the infectious processes due to its high amount of pre-existing phenolic compounds which act as an antifungal agent, inhibiting pathogen development and progression in comparison to cv. Galega. Del Rio et al. [63] reported the presence of some phenolic compounds (e.g., catechin) in olives that were linked to a positive reaction against Phytophtora spp. making the fruit an unfavorable host for further pathogen development.
When comparing the phenolic content (Figure 3) with the PAL enzyme activity (Figure 2) it is interesting to note that the last has increased significantly in the susceptible cultivar straight after elicitation. Our results suggest that the defense mechanism, in susceptible genotypes, involves the activation of PAL, a key enzyme of the phenylpropanoid pathway, from which phenolic metabolites are synthesized. However, it does not seem to be sufficient to block the fungus progression but it can limit the infection level in cultivars that present a higher level in their composition. Considering some of the primary stress response, it is generalized that the levels of PAL activity, considering gene expression and phenylpropanoids increase, is one of the firstly defense responses in plants [23]. In pepper (Capsicum annuum L.), CaPAL1 was proposed to function as a positive regulator of plant innate immunity [64], in rice (Oryza sativa L.) OsPAL4 was associated with improved broad-spectrum disease resistance [23], suggesting that defense response genes, including the PAL family, play a key role on host resistance.
5. Conclusions
In plant-pathogen studies, the combination of gene expression and metabolic data sets is currently being attempted to study metabolic pathways. Understanding the molecular mechanisms underlying olive anthracnose is critical for maintaining high olive oil production levels, and for developing strategies to control this disease. In this study, the infection process changed the secondary metabolism, and after an elicited increased activity of PAL enzyme and PAL gene expression, which is involved in olive anthracnose defense response. Furthermore, the tolerant cv. Picual presented the highest PAL gene expression at 16 hai, which could be linked to a genetic resistance to C. acutatum. Moreover, the constitutive phenolic content of fruits and whether their biosynthesis is activated or inhibited in olive-C. acutatum pathosystem, are crucial to explain the changes in gene expression in the early stages of pathogen colonization and infection process.
Author Contributions
Conceptualization, I.G., P.M.-L., A.B. and S.G.; Funding acquisition, P.M.-L. and A.B.; Investigation, I.G., Teresa Carvalho, A.B., P.M.-L. and S.G.; Methodology, I.G., T.C., A.B. and S.G.; Supervision, P.M.-L.; Writing—original draft, I.G. and S.G.; Writing—review and editing, I.G., P.M.-L., A.B. and S.G.
Funding
This research was funded by SFRH/BD/78013/2011 to Irene Gouvinhas and Postdoctoral grant SFRH/BPD/70378/2010 to Sónia Gomes from the Fundação para a Ciência e Tecnologia (POPH-QREN). This research was supported by the Project INNOFOOD–INNOvation in the FOOD sector through the valorization of food and agro-food by-products–NORTE-07-0124-FEDER-0000029, financed by the North Portugal Regional Operational Programme (ON.2–O Novo Norte) under the National Strategic Reference Framework (QREN), through FEDER, as well as by PIDDAC through FCT/MEC, and by the projects UID/MULTI/04046/2013, UID/AGR/04033/2019 and POCI-01-0145-FEDER-006958.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moral, J.; Trapero, A. Assessing the Susceptibility of Olive Cultivars to Anthracnose Caused by Colletotrichum acutatum. Plant Dis. 2009, 93, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Talhinhas, P.; Sreenivasaprasad, S.; Neves-Martins, J.; Oliveira, H. Molecular and Phenotypic Analyses Reveal Association of Diverse Colletotrichum acutatum Groups and a Low Level of C. gloeosporioides with Olive Anthracnose. Appl. Environ. Microbiol. 2005, 71, 2987–2998. [Google Scholar] [CrossRef] [PubMed]
- Trapero, A.; Blanco, M.A. Enfermedades. In El Cultivo de Olivo; Barranco, D., Fernández-Escobar, R., Rallo, L., Eds.; Mundi-Prensa: Madrid, Spain, 2008; pp. 557–614. ISBN 9788484767145. [Google Scholar]
- Cacciola, S.O.; Faedda, R.; Sinatra, F.; Agosteo, G.E.; Schena, L.; Frisullo, S.; Magnano di San Lio, G. Olive Anthracnose. J. Plant Pathol. 2012, 94, 29–44. [Google Scholar]
- Leyva-Pérez, M.D.L.O.; Jiménez-Ruiz, J.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Barroso, J.B.; Luque, F.; Mercado-Blanco, J. Tolerance of olive (Olea europaea) cv Frantoio to Verticillium dahliae relies on both basal and pathogen-induced differential transcriptomic responses. New Phytol. 2018, 217, 671–686. [Google Scholar] [CrossRef] [PubMed]
- Moral, J.; Alsalimiya, M.; Roca, L.F.; Díez, C.M.; León, L.; de la Rosa, R.; Barranco, D.; Rallo, L.; Trapero, A. Relative Susceptibility of New Olive Cultivars to Spilocaea oleagina, Colletotrichum acutatum, and Pseudocercospora cladosporioides. Plant Dis. 2015, 99, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.T.; Piteira, M.C.C.; Clara, M.I.E. Identificação de Colletotrichum acutatum em Olea europaea afectada pela doença da gafa. In Proceedings of the III Simpósio Nacional de Olivicultura-Revista de Ciências Agrárias, Castelo Branco, Portugal, 29–31 October 2013. [Google Scholar]
- Gomes, S.; Prieto, P.; Martins-Lopes, P.; Carvalho, T.; Martin, A.; Guedes-Pinto, H. Development of Colletotrichum acutatum on Tolerant and Susceptible Olea europaea L. cultivars: A Microscopic Analysis. Mycopathologia 2009, 168, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Moral, J.; Bouhmidi, K.; Trapero, A. Influence of Fruit Maturity, Cultivar Susceptibility, and Inoculation Method on Infection of Olive Fruit by Colletotrichum acutatum. Plant Dis. 2008, 92, 1421–1426. [Google Scholar] [CrossRef]
- Talhinhas, P.; Loureiro, A.; Oliveira, H. Olive anthracnose: A yield-and oil quality-degrading disease caused by several species of Colletotrichum that differ in virulence, host preference and geographical distribution. Mol. Plant Pathol. 2018, 19, 1797–1807. [Google Scholar] [CrossRef]
- Amaral, J.S.; Mafra, I.; Oliveira, M.B.P.P. Characterization of Three Portuguese Varietal Olive Oils Based on Fatty Acids, Triacylglycerols, Phytosterols and Vitamin E Profiles. In Olives and Olive Oil in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2010; pp. 581–589. [Google Scholar]
- Moral, J.; Xaviér, C.J.; Viruega, J.R.; Roca, L.F.; Caballero, J.; Trapero, A. Variability in Susceptibility to Anthracnose in the World Collection of Olive Cultivars of Cordoba (Spain). Front. Plant Sci. 2017, 8, 1892. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, J.; Leyva-Pérez, M.D.L.O.; Schilirò, E.; Barroso, J.B.; Bombarely, A.; Mueller, L.; Mercado-Blanco, J.; Luque, F. Transcriptomic Analysis of L. Roots during the Early Infection Process. Plant Genome 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Bhadauria, V.; Banniza, S.; Wang, L.-X.; Wei, Y.-D.; Peng, Y.-L. Proteomic studies of phytopathogenic fungi, oomycetes and their interactions with hosts. Eur. J. Plant Pathol. 2010, 126, 81–95. [Google Scholar] [CrossRef]
- Gomes, S.; Bacelar, E.; Martins-Lopes, P.; Carvalho, T.; Guedes-Pinto, H. Infection Process of Olive Fruits by Colletotrichum acutatum and the Protective Role of the Cuticle and Epidermis. J. Agric. Sci. 2012, 4, 101. [Google Scholar] [CrossRef]
- Mithfer, A.; Boland, W.; Maffei, M.E. Chemical Ecology of Plant–Insect Interactions. In Molecular Aspects of Plant Disease Resistance; Wiley-Blackwell: Oxford, UK, 2008; pp. 261–291. [Google Scholar]
- Truman, W.; Zabala, M.T.; Grant, M. Type III effectors orchestrate a complex interplay between transcriptional networks to modify basal defence responses during pathogenesis and resistance. Plant J. 2006, 46, 14–33. [Google Scholar] [CrossRef] [PubMed]
- Anand, T.; Bhaskaran, R.; Raguchander, T.; Samiyappan, R.; Prakasam, V.; Gopalakrishnan, C. Defence responses of chilli fruits to Colletotrichum capsici and Alternaria alternata. Biol. Plant. 2009, 53, 553–559. [Google Scholar] [CrossRef]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Guidarelli, M.; Carbone, F.; Mourgues, F.; Perrotta, G.; Rosati, C.; Bertolini, P.; Baraldi, E. Colletotrichum acutatum interactions with unripe and ripe strawberry fruits and differential responses at histological and transcriptional levels. Plant Pathol. 2011, 60, 685–697. [Google Scholar] [CrossRef]
- Montesano, M.; Brader, G.; Palva, E.T. Pathogen derived elicitors: Searching for receptors in plants. Mol. Plant Pathol. 2003, 4, 73–79. [Google Scholar] [CrossRef]
- Salazar, S.M.; Castagnaro, A.P.; Arias, M.E.; Chalfoun, N.; Tonello, U.; Díaz Ricci, J.C. Induction of a defense response in strawberry mediated by an avirulent strain of Colletotrichum. Eur. J. Plant Pathol. 2007, 117, 109–122. [Google Scholar] [CrossRef]
- Shan, X.C.; Goodwin, P.H. Reorganization of Filamentous Actin in Nicotiana benthamiana Leaf Epidermal Cells Inoculated with Colletotrichum destructivum and Colletotrichum graminicola. Int. J. Plant Sci. 2005, 166, 31–39. [Google Scholar] [CrossRef]
- Tonnessen, B.W.; Manosalva, P.; Lang, J.M.; Baraoidan, M.; Bordeos, A.; Mauleon, R.; Oard, J.; Hulbert, S.; Leung, H.; Leach, J.E. Rice phenylalanine ammonia-lyase gene OsPAL4 is associated with broad spectrum disease resistance. Plant Mol. Biol. 2015, 87, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Pattern-recognition receptors in plant innate immunity. Curr. Opin. Immunol. 2008, 20, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.J. Defense-Related Proteins in Higher Plants. Annu. Rev. Biochem. 1990, 59, 873–907. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Chacón, A.L.; Camperos-Reyes, J.E.; Ávila Diazgranados, R.A.; Romero, H.M. Biochemical and physiological responses of oil palm to bud rot caused by Phytophthora palmivora. Plant Physiol. Biochem. 2013, 70, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Mariotti, R.; Panara, F.; Caporali, S.; Urbani, S.; Veneziani, G.; Esposto, S.; Taticchi, A.; Rosati, A.; Rao, R.; et al. Olive phenolic compounds: Metabolic and transcriptional profiling during fruit development. BMC Plant Biol. 2012, 12, 162. [Google Scholar] [CrossRef] [PubMed]
- Gouvinhas, I.; Machado, N.; Sobreira, C.; Domínguez-Perles, R.; Gomes, S.; Rosa, E.; Barros, A. Critical Review on the Significance of Olive Phytochemicals in Plant Physiology and Human Health. Molecules 2017, 22, 1986. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Naqvi, S.H.; Dahot, M.U.; Rafiq, M.; Khan, M.Y.; Ibrahim, I.; Lashari, K.H.; Ali, A.; Korai, A.L. Anti-microbial efficacy and biochemical analysis from different parts of Acacia nilotica L. and Ricinus communis L. extracts. J. Med. Plants Res. 2011, 5, 6299–6308. [Google Scholar] [CrossRef]
- Reimers, P.J.; Leach, J.E. Race-specific resistance to Xanthomonas oryzae pv. oryzae conferred by bacterial blight resistance gene Xa-10 in rice (Oryza sativa) involves accumulation of a lignin-like substance in host tissues. Physiol. Mol. Plant Pathol. 1991, 38, 39–55. [Google Scholar] [CrossRef]
- Schovánková, J.; Opatová, H. Changes in phenols composition and activity of phenylalanine-ammonia lyase in apples after fungal infections. Hortic. Sci. 2011, 38, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tovar, M.J.; Romero, M.P.; Girona, J.; Motilva, M.J. L-Phenylalanine ammonia-lyase activity and concentration of phenolics in developing olive (Olea europaea L. cv Arbequina) fruit grown under different irrigation regimes. J. Sci. Food Agric. 2002, 82, 892–898. [Google Scholar] [CrossRef]
- Agudelo-Romero, P.; Erban, A.; Rego, C.; Carbonell-Bejerano, P.; Nascimento, T.; Sousa, L.; Martínez-Zapater, J.M.; Kopka, J.; Fortes, A.M. Transcriptome and metabolome reprogramming in Vitis vinifera cv. Trincadeira berries upon infection with Botrytis cinerea. J. Exp. Bot. 2015, 66, 1769–1785. [Google Scholar] [CrossRef] [PubMed]
- Alkan, N.; Fortes, A.M. Insights into molecular and metabolic events associated with fruit response to post-harvest fungal pathogens. Front. Plant Sci. 2015, 6, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermoso Fernández, M.; Gonzélez Delgado, J.; Uceda Ojeda, M.; García-Ortiz Rodríguez, Á.; Morales Bernardino, J.; Frías Ruiz, L.; Fernández García, Á. Elaboración de Aceite de Oliva de Calidad. Obtención por el Sistema de Dos Fases, 3rd ed.; Junta de Andalucía–Consejería de Agricultura y Pesca, Ed.; VICECONCEJERÍA: Sevilla, Spain, 1991; ISBN 84-89802-37-8. [Google Scholar]
- Gomes, S.; Martins-Lopes, P.; Bacelar, E.; Guedes-Pinto, H. An integrated functional genomic approach in the Olea europaea L.–Colletotrichum acutatum pathosystem. J. Biotechnol. 2010, 150, 503. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- McCallum, J.A.; Walker, J.R.L. Phenolic biosynthesis during grain development in wheat: Changes in phenylalanine Ammonia-lyase activity and soluble phenolic content. J. Cereal Sci. 1990, 11, 35–49. [Google Scholar] [CrossRef]
- Machado, M.; Felizardo, C.; Fernandes-Silva, A.A.; Nunes, F.M.; Barros, A. Polyphenolic compounds, antioxidant activity and l-phenylalanine ammonia-lyase activity during ripening of olive cv. “Cobrançosa” under different irrigation regimes. Food Res. Int. 2013, 51, 412–421. [Google Scholar] [CrossRef]
- Gouvinhas, I.; Domínguez-Perles, R.; Gironés-Vilaplana, A.; Carvalho, T.; Machado, N.; Barros, A. Kinetics of the Polyphenolic Content and Radical Scavenging Capacity in Olives through On-Tree Ripening. J. Chem. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Barros, A.I.; Freire, I.; Gonçalves, B.; Bacelar, E.; Gomes, S.; Lopes, J.; Guedes-Pinto, H.; Martins-Lopes, P. Evaluation of chemical and phenotypic changes in Blanqueta, Cobrançosa, and Galega during olive fruits ripening. CyTA–J. Food 2013, 11, 136–141. [Google Scholar] [CrossRef]
- Morelló, J.-R.; Romero, M.-P.; Ramo, T.; Motilva, M.-J. Evaluation of l-phenylalanine ammonia-lyase activity and phenolic profile in olive drupe (Olea europaea L.) from fruit setting period to harvesting time. Plant Sci. 2005, 168, 65–72. [Google Scholar] [CrossRef]
- Ortega-García, F.; Peragón, J. Phenylalanine Ammonia-Lyase, Polyphenol Oxidase, and Phenol Concentration in Fruits of Olea europaea L. cv. Picual, Verdial, Arbequina, and Frantoio during Ripening. J. Agric. Food Chem. 2009, 57, 10331–10340. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; Preuss, A.; DE Vos, R.C.H.; D’amico, E.; Perrotta, G.; Bovy, A.G.; Martens, S.; Rosati, C. Developmental, genetic and environmental factors affect the expression of flavonoid genes, enzymes and metabolites in strawberry fruits. Plant. Cell Environ. 2009, 32, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.W.; Breen, P.J. Activity of Phenylalanine Ammonia-Lyase (PAL) and Concentrations of Anthocyanins and Phenolics in Developing Strawberry Fruit. J. Am. Soc. Hortic. Sci. 1991, 116, 865–869. [Google Scholar] [CrossRef]
- Gouvinhas, I.; Machado, N.; Gironés-Vilaplana, A.; Gomes, S.; Carvalho, T.; Domínguez-Perles, R.; Barros, A.I.R.N.A. Sorting out the value of spectroscopic tools to assess the Colletotrichum acutatum impact in olive cultivars with different susceptibilities. J. Chemom. 2016, 30, 548–558. [Google Scholar] [CrossRef]
- Goetz, G.; Fkyerat, A.; Métais, N.; Kunz, M.; Tabacchi, R.; Pezet, R.; Pont, V. Resistance factors to grey mould in grape berries: Identification of some phenolics inhibitors of Botrytis cinerea stilbene oxidase. Phytochemistry 1999, 52, 759–767. [Google Scholar] [CrossRef]
- Ockels, F.S.; Eyles, A.; McPherson, B.A.; Wood, D.L.; Bonello, P. Phenolic Chemistry of Coast Live Oak Response to Phytophthora ramorum Infection. J. Chem. Ecol. 2007, 33, 1721–1732. [Google Scholar] [CrossRef] [PubMed]
- Wallis, C.; Eyles, A.; Chorbadjian, R.; McSpadden Gardener, B.; Hansen, R.; Cipollini, D.; Herms, D.A.; Bonello, P. Systemic induction of phloem secondary metabolism and its relationship to resistance to a canker pathogen in Austrian pine. New Phytol. 2008, 177, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Gouvinhas, I.; de Almeida, J.M.M.M.; Carvalho, T.; Machado, N.; Barros, A.I.R.N.A. Discrimination and characterisation of extra virgin olive oils from three cultivars in different maturation stages using Fourier transform infrared spectroscopy in tandem with chemometrics. Food Chem. 2015, 174, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Curvers, K.; Seifi, H.; Mouille, G.; de Rycke, R.; Asselbergh, B.; Van Hecke, A.; Vanderschaeghe, D.; Höfte, H.; Callewaert, N.; Van Breusegem, F.; et al. Abscisic Acid Deficiency Causes Changes in Cuticle Permeability and Pectin Composition That Influence Tomato Resistance to Botrytis cinerea. Plant Physiol. 2010, 154, 847–860. [Google Scholar] [CrossRef]
- Bassolino, L.; Zhang, Y.; Schoonbeek, H.; Kiferle, C.; Perata, P.; Martin, C. Accumulation of anthocyanins in tomato skin extends shelf life. New Phytol. 2013, 200, 650–655. [Google Scholar] [CrossRef] [Green Version]
- Slatnar, A.; Mikulic Petkovsek, M.; Halbwirth, H.; Stampar, F.; Stich, K.; Veberic, R. Enzyme activity of the phenylpropanoid pathway as a response to apple scab infection. Ann. Appl. Biol. 2010, 156, 449–456. [Google Scholar] [CrossRef]
- Schwalb, P.; Feucht, W. Changes in the concentration of phenolic substances in the bark during the annual development of the cherry tree (Prunus avium L.). Adv. Hortic. Sci. 1999, 13, 71–75. [Google Scholar]
- Usenik, V.; Mikuli-Petkovšek, M.; Solar, A.; Štampar, F. Flavonols of leaves in relation to apple scab resistance. J. Plant Dis. Prot. 2004, 111, 137–144. [Google Scholar] [CrossRef]
- Prusky, D.; Kobiler, I.; Ardi, R.; Beno-Moalem, D.; Yakoby, N.; Keen, N.T. Resistance Mechanisms of Subtropical Fruits to Colletotrichum gloeosporioides. In Colletotrichum: Host Specificity, Pathology, and Host-Pathogen Interaction; Prusky, D., Freeman, S., Dickman, M., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2000; pp. 232–244. ISBN 978-0-89054-258-3. [Google Scholar]
- Mayr, U.; Michalek, S.; Treutter, D.; Feucht, W. Phenolic Compounds of Apple and their Relationship to Scab Resistance. J. Phytopathol. 1997, 145, 69–75. [Google Scholar] [CrossRef]
- Michalek, S.; Mayr, U.; Treutter, D.; Lux-Endrich, A.; Gutmann, M.; Feucht, W.; Geibel, M. Role of Flavan-3-ols in resistance of apple trees to Venturia inaequalis. Acta Hortic. 1998, 484, 535–540. [Google Scholar] [CrossRef]
- Walker, J.C.; Stahmann, M.A. Chemical Nature of Disease Resistance in Plants. Annu. Rev. Plant Physiol. 1955, 6, 351–366. [Google Scholar] [CrossRef]
- Del Río, J. Enhancement of phenolic compounds in olive plants (Olea europaea L.) and their influence on resistance against Phytophthora sp. Food Chem. 2003, 83, 75–78. [Google Scholar] [CrossRef]
- Kim, D.S.; Hwang, B.K. An important role of the pepper phenylalanine ammonia-lyase gene (PAL1) in salicylic acid-dependent signalling of the defence response to microbial pathogens. J. Exp. Bot. 2014, 65, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Relative gene expression of phenylalanine ammonia-lyase (PAL) of phenylpropanoid pathway by quantitative polymerase chain reaction (qPCR) in developing (green, semi-ripe, and ripe stage) and infected fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual cultivars during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.
Figure 1.
Relative gene expression of phenylalanine ammonia-lyase (PAL) of phenylpropanoid pathway by quantitative polymerase chain reaction (qPCR) in developing (green, semi-ripe, and ripe stage) and infected fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual cultivars during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.

Figure 2.
Activity of PAL enzyme in developing (green, semi-ripe, and ripe stage) and infected fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual cultivars during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.
Figure 2.
Activity of PAL enzyme in developing (green, semi-ripe, and ripe stage) and infected fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual cultivars during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.

Figure 3.
Total phenolic composition in developing (green, semi-ripe, and ripe stage) and infected olive fruit samples (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega Vulgar; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.
Figure 3.
Total phenolic composition in developing (green, semi-ripe, and ripe stage) and infected olive fruit samples (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega Vulgar; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.

Figure 4.
Ortho-diphenols content in developing (green, semi-ripe, and ripe stage) and infected olive fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.
Figure 4.
Ortho-diphenols content in developing (green, semi-ripe, and ripe stage) and infected olive fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.

Figure 5.
Flavonoids content in developing (green, semi-ripe, and ripe stage) and infected olive fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.
Figure 5.
Flavonoids content in developing (green, semi-ripe, and ripe stage) and infected olive fruits (non-infected fruit (control), 0, 16, 48, and 144 h after C. acutatum inoculation (hai)), from susceptible cv. Galega; moderate-tolerant cv. Cobrançosa, and tolerant cv. Picual during two crop years (2016 and 2017). Bars indicate standard error of three biological replicates at each sampling time point within cultivars and data were subjected to analysis of variance (ANOVA) and multiple range test (Tukey’s test) with a significance of p < 0.05.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Schedule for olive sampling.
| Cultivar | Ripening Index | Hours after Infection in Ripe Fruits (hai) | Susceptibility to Colletotrichum acutatum |
|---|---|---|---|
| Galega Vulgar | Green (RI = 1.30) Semi-ripe (RI = 2.74) Ripe (RI = 4.87) | Control (non-infected fruits) | Susceptible |
| 0 | |||
| 16 | |||
| 48 | |||
| 144 | |||
| Cobrançosa | Green (RI = 1.30) Semi-ripe (RI = 2.96) Ripe (RI = 4.67) | Control (non-infected fruits) | Moderate-Tolerant |
| 0 | |||
| 16 | |||
| 48 | |||
| 144 | |||
| Picual | Green (RI = 0.8) Semi-ripe (RI = 2.43) Ripe (RI = 4.57) | Control (non-infected fruits) | Tolerant |
| 0 | |||
| 16 | |||
| 48 | |||
| 144 | |||
| RI = Ripening Index | |||
Table 2.
Primers used for quantitative real-time polymerase chain reaction (PCR) assays.
| Gene | Accession Number | Primer | Sequence (5′–3′) | Amplicon Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|---|
| ACT | AF545569 | Sense | AGCTTGCTTATGTTGCTCTC | 169 | 59 |
| Antisense | GATTCCATTCCAATCAAAGA | ||||
| GAPDH | EF506494 | Sense | TTCTGCAACGATAGACTCCT | 224 | 59 |
| Antisense | AAGCTTCGTAAACTTTGCAC | ||||
| PAL | KJ511868 | Sense | CATTGAAAGGTAGCCATCTA | 101 | 59 |
| Antisense | CTAGCAAATTGGAAGAGGTT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gouvinhas, I.; Martins-Lopes, P.; Carvalho, T.; Barros, A.; Gomes, S. Impact of Colletotrichum acutatum Pathogen on Olive Phenylpropanoid Metabolism. Agriculture 2019, 9, 173. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9080173
AMA Style
Gouvinhas I, Martins-Lopes P, Carvalho T, Barros A, Gomes S. Impact of Colletotrichum acutatum Pathogen on Olive Phenylpropanoid Metabolism. Agriculture. 2019; 9(8):173. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9080173
Chicago/Turabian StyleGouvinhas, Irene, Paula Martins-Lopes, Teresa Carvalho, Ana Barros, and Sónia Gomes. 2019. "Impact of Colletotrichum acutatum Pathogen on Olive Phenylpropanoid Metabolism" Agriculture 9, no. 8: 173. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture9080173
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.