Physiological and Biochemical Responses to Salt Stress in Cultivated Eggplant (Solanum melongena L.) and in S. insanum L., a Close Wild Relative

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Layout
2.2. Electrical Conductivity of the Substrate
2.3. Gaseous Exchange
2.4. Evaluation of Growth Parameters
2.5. Ion Quantification
2.6. Quantification of Photosynthetic Pigments
2.7. Quantification of Osmolytes
2.8. Measurement of Malondialdehyde (MDA) and Antioxidant Compounds
2.9. Antioxidant Enzyme Activities
2.10. Statistical Analysis
3. Results
3.1. Substrate Electrical Conductivity
3.2. Analysis of Morphological and Photosynthetic Parameters
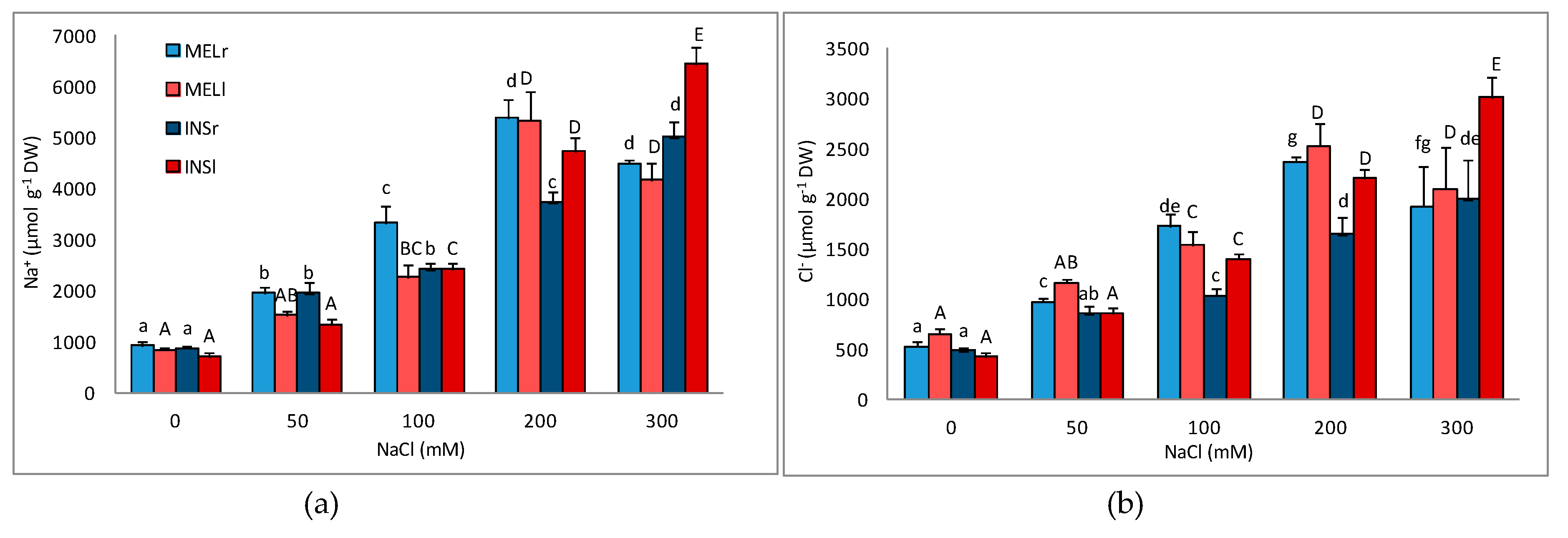
3.3. Ion Accumulation
3.4. Osmolytes, MDA, and Antioxidants
3.5. Principal Component Analysis
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wicke, B.; Smeets, E.; Dornburg, V.; Vashev, B.; Gaiser, T.; Turkenburg, W.; Faaij, A. The global technical and economic potential of bioenergy from salt-affected soils. Energy Environ. Sci. 2011, 8, 2669–2681. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.S.; Mohammed, M.; Daba, A.W.; Hailu, B.; Belay, G.; Tesfaye, A.; Ertebo, A-M. Improving agricultural productivity on salt-affected soils in Ethiopia: Farmers’ perceptions and proposals. Afr. J. Agric. Res. 2019, 14, 897–906. [Google Scholar] [CrossRef]
- Daliakopoulos, I.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.; Varouchakis, E.; Karatzas, G.; Ritsema, C. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology, 4th ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2003. [Google Scholar]
- Tang, X.; Mu, X.; Shao, H.; Wang, H.; Brestic, M. Global plant-responding mechanisms to salt stress: Physiological and molecular levels and implications in biotechnology. Crit. Rev. Biotechnol. 2015, 35, 425–437. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). FAOSTAT: Food and Agriculture Data; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; Available online: http://www.fao.org/faostat/en/ (accessed on 14 February 2020).
- Ünlükara, A.; Kurunç, A.; Kesmez, G.D.; Yurtseven, E.; Suarez, D.L. Effects of salinity on eggplant (Solanum melongena L.) growth and evapotranspiration. Irrig. Drain. 2010, 59, 203–214. [Google Scholar] [CrossRef]
- Mennella, G.; Lo Scalzo, R.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Acciarri, N.; de Almeida, A.E.; Rotino, G.L. Chemical and bioactive quality traits during fruit ripening in eggplant (S. melongena L.) and allied species. J. Agric. Food Chem. 2012, 60, 11821–11831. [Google Scholar] [CrossRef]
- Plazas, M.; López Gresa, M.P.; Vilanova, S.; Torres, C.; Hurtado, M.; Gramazio, P.; Andújar, I.; Herráiz, F.J.; Bellés, J.M.; Prohens, J. Diversity and relationships in key traits for functional and apparent quality in a collection of eggplant: Fruit phenolics content, antioxidant activity, polyphenol oxidase activity, and browning. J. Agric. Food Chem. 2013, 61, 8871–8879. [Google Scholar] [CrossRef]
- Plazas, M.; Vilanova, S.; Gramazio, P.; Rodríguez-Burruezo, A.; Fita, A.; Herráiz, F.J.; Prohens, J. Interspecific hybridization between eggplant and wild relatives from different genepools. J. Am. Soc. Hort. Sci. 2016, 141, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Kouassi, B.; Prohens, J.; Gramazio, P.; Kouassi, A.B.; Vilanova, S.; Galán-Ávila, A.; Herráiz, F.J.; Kouassi, A.; Seguí-Simarro, J.M.; Plazas, M. Development of backcross generations and new interspecific hybrid combinations for introgression breeding in eggplant (Solanum melongena). Sci. Hortic. 2016, 213, 199–207. [Google Scholar] [CrossRef]
- Gramazio, P.; Prohens, J.; Plazas, M.; Mangino, G.; Herraiz, F.J.; Vilanova, S. Development and genetic characterization of advanced backcross materials and an introgression line population of Solanum incanum in a S. melongena background. Front. Plant Sci. 2017, 8, 1477. [Google Scholar] [CrossRef]
- García-Fortea, E.; Gramazio, P.; Vilanova, S.; Fita, A.; Mangino, G.; Villanueva, G.; Arrones, A.; Knapp, S.; Prohens, J.; Plazas, M. First successful backcrossing towards eggplant (Solanum melongena) of a New World species, the silverleaf nightshade (S. elaeagnifolium), and characterization of interspecific hybrids and backcrosses. Sci. Hortic. 2019, 246, 563–573. [Google Scholar] [CrossRef]
- Knapp, S.; Vorontsova, M.S. A revision of the “African non-spiny” clade of Solanum L. (Solanum sections Afrosolanum Bitter, Benderianum Bitter, Lemurisolanum Bitter, Lyciosolanum Bitter, Macronesiotes Bitter, and Quadrangulare Bitter: Solanaceae). PhytoKeys 2016, 66, 1–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranil, R.H.; Prohens, J.; Aubriot, X.; Niran, H.M.; Plazas, M.; Fonseka, R.M.; Vilanova, S.; Gramazio, P.; Knapp, S. Solanum insanum L. (subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant (S. melongena L.): A review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genet. Resour. Crop Evol. 2017, 64, 1707–1722. [Google Scholar] [CrossRef] [Green Version]
- Davidar, P.; Snow, A.A.; Rajkumar, M.; Pasquet, R.; Daunay, M.C.; Mutegi, E. The potential for crop to wild hybridization in eggplant (Solanum melongena; Solanaceae) in southern India. Am. J. Bot. 2015, 102, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinci, I.E.; Akinci, S.; Yilmaz, K.; Dikici, H. Response of eggplant varieties (Solanum melongena) to salinity in germination and seedling stages. N. Zeal. J. Crop Hort. Sci. 2004, 32, 193–200. [Google Scholar] [CrossRef]
- Ranil, R.H.; Niran, H.M.L.; Plazas, M.; Fonseka, R.M.; Fonsekad, H.H.; Vilanova, S.; Andújar, I.; Gramazio, P.; Fita, A.; Prohens, J. Improving seed germination of the eggplant rootstock Solanum torvum by testing multiple factors using an orthogonal array design. Sci. Hortic. 2015, 193, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Weimberg, R. Solute adjustments in leaves of two species of wheat at two different stages of growth in response to salinity. Physiol. Plant. 1987, 70, 381–388. [Google Scholar] [CrossRef]
- Lichenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hodges, D.M.; Delong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.C.; Palazzodemello, J.C. Application and analysis of the Folin Ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Gil, R.; Bautista, I.; Boscaiu, M.; Lidón, A.; Wankhade, S.; Sánchez, H.; Llinares, J.; Vicente, O. Responses of five Mediterranean halophytes to seasonal changes in environmental conditions. AoB Plants 2014, 6, plu049. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Conell, J.P.; Mullet, J.E. Pea chloroplast glutathione reductase: Purification and characterization. Plant Physiol. 1986, 82, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovas. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Zayova, E.; Philipov, P.; Nedev, T.; Stoeva, D. Response of in vitro cultivated eggplant (Solanum melongena L.) to salt and drought stress. AgroLife Sci. J. 2017, 6, 276–282. [Google Scholar]
- Hannachi, S.; Van Labeke, M.C. Salt stress affects germination, seedling growth and physiological responses differentially in eggplant cultivars (Solanum melongena L.). Sci. Hortic. 2018, 228, 56–65. [Google Scholar] [CrossRef]
- Foolad, M.R. Recent advances in genetics of salt tolerance in tomato. Plant Cell Tissue Organ Cult. 2004, 76, 101–119. [Google Scholar] [CrossRef]
- Rotino, G.L.; Sala, T.; Toppino, L. Eggplant. In Alien Gene Transfer in Crop Plants, Volume 2: Achievements and Impacts; Pratap, A., Kumar, J., Eds.; Springer Science+Business Media: New York, NY, USA, 2014. [Google Scholar]
- Plazas, M.; Nguyen, T.; González-Orenga, S.; Fita, A.; Vicente, O.; Prohens, J.; Boscaiu, M. Comparative analysis of the responses to water stress in eggplant (Solanum melongena) cultivars. Plant Physiol. Biochem. 2019, 143, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, Z.; Ayyub, C.M.; Amjad, M.; Ahmad, R. Assessment of biochemical and ionic attributes against salt stress in eggplant (Solanum melongena L.) genotypes. J. Anim. Plant Sci. 2017, 27, 503–509. [Google Scholar]
- Hanachi, S.; Van Labeke, M.C.; Mehouachi, T. Application of chlorophyll fluorescence to screen eggplant (Solanum melongena L.) cultivars for salt tolerance. Photosynthetica 2014, 52, 57–62. [Google Scholar] [CrossRef]
- Intergovernmental panel on climate change (IPCC). Climate Change 2014: Impacts, Adaptation, and Vulnerability; 5th Assessment Report, WGII; Cambridge University Press: Cambridge, UK, 2015; Available online: http://www.ipcc.ch/report/ar5/wg2/ (accessed on 18 March 2020).
- Richards, L. Diagnosis and improvement of saline and alkali soils. Soil Sci. 1954, 78, 154. [Google Scholar] [CrossRef]
- Al Hassan, M.; López-Gresa, M.P.; Boscaiu, M.; Vicente, O. Stress tolerance mechanisms in Juncus: Responses to salinity and drought in three Juncus species adapted to different natural environments. Funct. Plant Biol. 2016, 43, 949–960. [Google Scholar] [CrossRef]
- González-Orenga, S.; Ferrer-Gallego, P.P.; Laguna, E.; López-Gresa, M.P.; Donat-Torres, M.P.; Verdeguer, M.; Vicente, O.; Boscaiu, M. Insights on salt tolerance of two endemic Limonium species from Spain. Metabolites 2019, 9, 294. [Google Scholar] [CrossRef] [Green Version]
- Al Hassan, M.; Morosan, M.; López-Gresa, M.D.P.; Prohens, J.; Vicente, O.; Boscaiu, M. Salinity-induced variation in biochemical markers provides insight into the mechanisms of salt tolerance in common (Phaseolus vulgaris) and runner (P. coccineus) beans. Int. J. Mol. Sci. 2016, 17, 1582. [Google Scholar] [CrossRef]
- Al Hassan, M.; Pacurar, A.; López-Gresa, M.P.; Donat-Torres, M.P.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Effects of salt stress on three ecologically distinct Plantago species. PLoS ONE 2016, 11, e0160236. [Google Scholar] [CrossRef]
- Jamil, M.; Rehman, S.U.; Lee, K.J.; Kim, J.M.; Kim, H.; Rha, E.S. Salinity reduced growth PS2 photochemistry and chlorophyll content in radish. Sci. Agric. 2007, 64, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.; Hernandez, J. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhu, Z.; Li, X.; Zha, D. Effects of cytokinin on photosynthetic gas exchange, chlorophyll fluorescence parameters and antioxidative system in seedlings of eggplant (Solanum melongena L.) under salinity stress. Acta Physiol. Plant. 2012, 34, 2105–2114. [Google Scholar] [CrossRef]
- Shaheen, S.; Naseer, S.; Ashraf, M.; Akram, N.A. Salt stress affects water relations, photosynthesis, and oxidative defense mechanisms in Solanum melongena L. J. Plant Interact. 2013, 8, 85–96. [Google Scholar] [CrossRef]
- Shahbaz, M.; Mushtaq, Z.; Andaz, F.; Masood, A. Does proline application ameliorate adverse effects of salt stress on growth, ions and photosynthetic ability of eggplant (Solanum melongena L.)? Sci. Hortic. 2013, 164, 507–511. [Google Scholar] [CrossRef]
- Volkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is not all about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, T.; Troke, P.F.; Yeo, A.R. The mechanism of salt tolerance in halophytes. Annu. Rev. Plant Physiol. 1977, 28, 89–121. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in non-halophytes. Annu. Rev. Plant Biol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Sarker, B.C.; Hara, M.; Uemura, M. Proline synthesis, physiological responses and biomass yield of eggplants during and after repetitive soil moisture stress. Sci. Hortic. 2005, 103, 387–402. [Google Scholar] [CrossRef]
- Gil, R.; Boscaiu, M.T.; Lull, C.; Bautista, I.; Lidón, A.; Vicente, O. Are soluble carbohydrates ecologically relevant for salt tolerance in halophytes? Funct. Plant Biol. 2013, 40, 805–818. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbr. | Treatment a | Species a | Interaction a | Residual | |
|---|---|---|---|---|---|
| Root length | RL | 11.42 * | 1.32 | 20.77 | 66.48 |
| Root fresh weight | RFW | 6.36 | 24.43 *** | 2.60 | 66.60 |
| Root water content | RWC | 55.17 *** | 14.41 *** | 5.12 | 25.29 |
| Stem elongation | SE | 68.59 *** | 2.26 | 6.44 * | 22.71 |
| Stem thickening | ST | 63.70 *** | 0.93 | 2.32 | 32.96 |
| Stem fresh weight | SFW | 56.05 *** | 0.26 | 3.30 | 40.54 |
| Stem water content | SWC | 5.04 | 4.83 | 4.66 | 85.47 |
| Increase in no. of leaves | Lno | 34.91 *** | 29.19 *** | 10.20 ** | 39.82 |
| Leaf area | LA | 34.69 *** | 29.19 *** | 10.20 ** | 25.69 |
| Leaf fresh weight | LFW | 43.15 *** | 10.54 ** | 11.11 * | 35.20 |
| Leaf water content | LWC | 7.45 | 7.10 * | 17.34 | 68.11 |
| Total fresh weight | TFW | 44.77 *** | 0.15 | 5.98 | 49.09 |
| Chlorophyll a | Chl a | 14.09 | 3.33 | 4.58 | 78.00 |
| Chlorophyll b | Chl b | 12.86 | 8.37 * | 9.39 | 69.43 |
| Carotenoids | Caro | 18.92 * | 13.35 *** | 4.19 | 63.40 |
| Photosynthestic rate | AN | 23.29 *** | 37.82 *** | 4.75 | 34.58 |
| Stomatal conductance | gs | 16.22 * | 29.92 *** | 3.63 | 50.23 |
| Int. CO2 concentration | Ci | 29.10 ** | 17.22 *** | 1.11 | 52.57 |
| Transpiration rate | E | 17.92 ** | 39.17 ** | 2.35 | 40.56 |
| Treatment (mM NaCl) | ||||||
|---|---|---|---|---|---|---|
| Trait | Taxa | 0 | 50 | 100 | 200 | 300 |
| RL | MEL | 26.2 ± 1.3 c | 26.7 ± 2.2 c | 24.6 ± 1.7 bc | 19.8 ± 1.7 ab | 17.8 ± 0.6 a |
| INS | 21.0 ± 3.5 A | 20.2 ± 0.5 A | 24.0 ± 1.1 A | 20.1 ± 2.5 A | 24.2 ± 2.9 A | |
| RFW | MEL | 9.0 ± 0.8 b | 9.8 ± 0.5 b | 9.1 ± 0.7 b | 9.1 ± 0.5 b | 6.9 ± 0.1 a |
| INS | 10.8 ± 2.1 A | 11.9 ± 0.8 A | 11.2 ± 0.3 A | 11.4 ± 0.9 A | 10.9 ± 0.8 A | |
| RWC | MEL | 71.2 ± 1.2 a | 76.8 ± 0.7 b | 78.8 ± 0.7 bc | 80.4 ± 0.4 c | 79.5 ± 0.5 bc |
| INS | 65.5 ± 3.6 A | 68.4 ± 1.2 AB | 76.0 ± 1.0 BC | 78.0 ± 0.5 C | 77.7 ± 0.6 C | |
| SE | MEL | 6.3 ± 1.9 c | 5.7 ± 1.1 bc | 5.8 ± 0.5 c | 3.7 ± 0.5 ab | 2.5 ± 0.2 a |
| INS | 8.3 ± 0.6 C | 6.7 ± 0.2 C | 4.9 ± 0.8 BC | 3.7 ± 0.2 AB | 3.3 ± 0.4 A | |
| ST | MEL | 3.5 ± 0.2 c | 2.9 ± 0.2 bc | 2.3 ± 0.1 bc | 1.6 ± 0.3 ab | 1.3 ± 0.3 a |
| INS | 3.2 ± 0.5 B | 2.2 ± 0.4 AB | 2.2 ± 0.3 AB | 1.8 ± 0.2 AB | 1.1 ± 0.1 A | |
| SFW | MEL | 4.4 ± 0.9 b | 4.7 ± 0.2 b | 3.9 ± 0.5 ab | 2.6 ± 0.2 ab | 1.9 ± 0.3 a |
| INS | 5.5 ± 1.0 C | 4.7 ± 0.2 BC | 3.5 ± 0.1 B | 2.5 ± 0.1 AB | 2.1 ± 0.1 A | |
| SWC | MEL | 68.4 ± 6.2 a | 78.1 ± 0.7 a | 76.8 ± 3.9 a | 73.6 ± 3.5 a | 75.1 ± 5.6 a |
| INS | 70.8 ± 2.3 A | 70.6 ± 1.1 A | 72.8 ± 1.7 A | 71.4 ± 0.7 A | 67.6 ± 1.4 A | |
| Lno | MEL | 2.4 ± 0.2 ab | 2.6 ± 0.2 ab | 3.0 ± 0.0 b | 1.8 ± 0.2 a | 1.8 ± 0.2 a |
| INS | 2.2 ± 0.4 C | 1.8 ± 0.4 BC | 1.4 ± 0.2 B | 0.8 ± 0.4 AB | −0.4 ± 0.2 A | |
| LA | MEL | 205.7 ± 12.0 c | 161.6 ± 8.4 b | 149.2 ± 4.1 ab | 132.0 ± 3.9 ab | 119.4 ± 8.6 a |
| INS | 143.4 ± 17.0 A | 148.6 ± 3.7 A | 139.8 ± 8.2 A | 136.6 ± 8.2 A | 105.5 ± 9.1 A | |
| LFW | MEL | 22.4 ± 1.9 d | 19.3 ± 1.0 cd | 16.6 ± 0.8 bc | 12.9 ± 1.1 b | 9.1 ± 0.7 a |
| INS | 13.8 ± 3.2 A | 14.8 ± 1.1 A | 14.4 ± 0.7 A | 12.0 ± 0.3 A | 9.6 ± 1.1 A | |
| LWC | MEL | 81.3 ± 2.3 a | 84.4 ± 4.8 a | 85.3 ± 1.7 a | 81.9 ± 2.7 a | 72.2 ± 2.8 a |
| INS | 75.5 ± 4.5 A | 78.3 ± 0.8 A | 79.5 ± 0.7 A | 80.8 ± 0.3 A | 82.2 ± 0.7 A | |
| TFW | MEL | 35.8 ± 2.8 d | 33.8 ± 1.5 cd | 29.7 ± 3.2 c | 24.5 ±0.8 b | 18.0 ± 1.3 a |
| INS | 30.1 ± 6.0 a | 31.5 ± 1.9 a | 29.1 ± 2.1 a | 25.9 ± 0.9 a | 22.7 ± 1.8 a | |
| Chl a | MEL | 9.4 ± 2.2 a | 8.4 ± 1.1 a | 11.0 ± 1.1 a | 5.7 ± 1.1 a | 8.1 ± 0.7 a |
| INS | 7.6 ± 1.9 A | 6.8 ± 1.9 A | 8.7 ± 1.1 A | 7.2 ± 0.5 A | 6.5 ± 0.5 A | |
| Chl b | MEL | 4.3 ± 1.0 a | 3.8 ± 0.6 a | 5.7 ± 1.4 a | 2.2 ± 0.3 a | 3.6 ± 0.4 a |
| INS | 3.3 ± 0.8 A | 2.5 ± 0.7 A | 2.9 ± 0.4 A | 2.7 ± 0.8 A | 3.1 ± 0.3 A | |
| Caro | MEL | 1.6 ± 0.4 a | 1.3 ± 0.5 a | 1.3 ± 0.4 a | 0.9 ± 0.4 a | 1.3 ± 0.3 a |
| INS | 2.1 ± 0.3 B | 1.9 ± 0.2 AB | 1.7 ± 0.2 AB | 1.4 ± 0.1 AB | 1.2 ± 0.1 A | |
| AN | MEL | 8.5 ± 1.1 b | 9.9 ± 1.3 b | 9.0 ± 0.8 b | 5.4 ± 0.9 ab | 3.4 ± 0.3 a |
| INS | 18.9 ± 3.3 B | 15.4 ± 1.5 AB | 18.2 ± 2.0 AB | 15.8 ± 1.9 AB | 7.7 ± 2.9 A | |
| gs | MEL | 0.11 ± 0.00 b | 0.11 ± 0.00 b | 0.09 ± 0.01 ab | 0.05 ± 0.00 ab | 0.03 ± 0.00 a |
| INS | 0.24 ± 0.10 A | 0.26 ± 0.10 A | 0.30 ± 0.15 A | 0.21 ± 0.10 A | 0.09 ± 0.03 A | |
| C | MEL | 234.6 ± 17.0 b | 230.0 ± 8.5 b | 213.8 ± 5.6 b | 195.0 ± 5.7 ab | 182.2 ± 9.7 a |
| INS | 274.6 ± 24.0 A | 248.2 ± 13.0 A | 243.2 ± 13.0 A | 222.0 ± 14.0 A | 219.2 ± 5.3 A | |
| E | MEL | 2.6 ± 0.5 b | 2.7 ± 0.4 b | 2.5 ± 0.1 b | 1.5 ± 0.2 ab | 0.9 ± 0.1a |
| INS | 4.7 ± 1.0 A | 5.3 ± 0.8 A | 5.8 ± 0.8 A | 4.8 ± 0.8 A | 2.6 ± 0.4 A | |
| Ion Contents and K+/Na+ Ratio | A a | B a | C a | AB a | AC a | BC a | ABC a | Residuals |
|---|---|---|---|---|---|---|---|---|
| Na+ | 84.70 *** | 0.24 | 0.04 | 2.00 ** | 1.16 * | 1.2 ** | 1.45 ** | 8.61 |
| Cl− | 79.30 *** | 1.85 *** | 1.81 *** | 2.52 ** | 1.29 * | 1.01 * | 2.13 * | 10.07 |
| K+ | 2.30 *** | 6.10 *** | 70.20 *** | 0.58 | 2.14 * | 0.61 | 3.39 ** | 14.69 |
| K+/Na+ | 71.27 *** | 0.04 | 15.54 *** | 0.25 | 10.42 *** | 0.01 | 0.16 | 2.26 |
| Trait | Treatment a | Species a | Interaction a | Residual |
|---|---|---|---|---|
| Pro | 63.60 *** | 18.20 *** | 14.06 *** | 4.29 |
| TSS | 5.31 | 22.87 *** | 18.76 * | 53.05 |
| MDA | 25.29 *** | 29.23 *** | 6.48 | 38.98 |
| TPC | 8.19 * | 28.90 *** | 15.32 * | 47.57 |
| TF | 27.01 ** | 1.83 | 4.30 | 66.85 |
| SOD | 13.76 * | 21.11 *** | 16.92 * | 48.07 |
| CAT | 2.82 | 13.21 * | 10.26 | 73.24 |
| GR | 16.50 | 1.59 | 11.30 | 70.60 |
| Treatment (mM NaCl) | ||||||
|---|---|---|---|---|---|---|
| Trait | Taxa | 0 | 50 | 100 | 200 | 300 |
| MDA | MEL | 145.7 ± 12.2 b | 134.4 ± 9.1 ab | 114.7 ± 4.1 ab | 107.3 ± 7.5 a | 107.9 ± 7.8 a |
| INS | 207.1 ±2 4.6 B | 143.9 ± 11.7 A | 145.3 ± 7.8 A | 145.2 ± 4.2 A | 162.1 ± 8.1 AB | |
| TPC | MEL | 12.3 ± 0.7 a | 11.1 ± 2.1 a | 6.3 ± 0.6 a | 5.7 ± 0.4 a | 7.8 ± 1.3 a |
| INS | 15.5 ± 0.8 B | 10.7 ± 1.8 AB | 7.3 ± 0.5 A | 6.3 ± 0.4 A | 10.6 ± 0.4 AB | |
| TF | MEL | 9.5 ± 2.3 a | 7.8 ± 3.4 a | 9.1 ± 2.9 a | 5.0 ± 0.7 a | 6.0 ± 0.6 a |
| INS | 15.5 ± 1.8 B | 10.5 ± 1.9 AB | 6.5 ± 0.9 A | 5.5 ± 0.6 A | 8.7 ± 0.7 A | |
| SOD | MEL | 377.5 ± 46.9 a | 272.4 ± 24.2 a | 180.5 ± 30.8 a | 315.0 ± 20.3 a | 412.8 ± 75.0 a |
| INS | 1181.0 ± 263.0 A | 464.9 ± 134.0 A | 915.0 ± 221.0 A | 586.3 ± 163.0 A | 315.6 ± 71.0 A | |
| CAT | MEL | 280.2 ± 92.8 a | 453.4 ± 84.5 a | 304.3 ± 69.7 a | 514.1 ± 96.2 a | 413.2 ± 131.0 a |
| INS | 1135.0 ± 416.0 A | 523.9 ± 80.4 A | 7214.0 ± 140.0 A | 560.8 ± 85.4 A | 692.1 ± 147 A | |
| GR | MEL | 2419.0 ± 454.0 a | 1937.0 ± 384.0 a | 1468.0 ± 433.0 a | 1426.0 ± 268.0 a | 1484.0 ± 263.0 a |
| INS | 2881 ± 684 A | 1523 ± 197 A | 3571 ± 764 A | 1390 ± 144 A | 1114 ± 77 A | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brenes, M.; Solana, A.; Boscaiu, M.; Fita, A.; Vicente, O.; Calatayud, Á.; Prohens, J.; Plazas, M. Physiological and Biochemical Responses to Salt Stress in Cultivated Eggplant (Solanum melongena L.) and in S. insanum L., a Close Wild Relative. Agronomy 2020, 10, 651. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050651
Brenes M, Solana A, Boscaiu M, Fita A, Vicente O, Calatayud Á, Prohens J, Plazas M. Physiological and Biochemical Responses to Salt Stress in Cultivated Eggplant (Solanum melongena L.) and in S. insanum L., a Close Wild Relative. Agronomy. 2020; 10(5):651. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050651
Chicago/Turabian StyleBrenes, Marco, Andrea Solana, Monica Boscaiu, Ana Fita, Oscar Vicente, Ángeles Calatayud, Jaime Prohens, and Mariola Plazas. 2020. "Physiological and Biochemical Responses to Salt Stress in Cultivated Eggplant (Solanum melongena L.) and in S. insanum L., a Close Wild Relative" Agronomy 10, no. 5: 651. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050651