Husk Leaf Senescence Characteristics of Spring Maize (Zea mays L.) Cultivated in Two Row Directions and Three Plant Spacings in Northeast China


Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Area of Husk Leaves
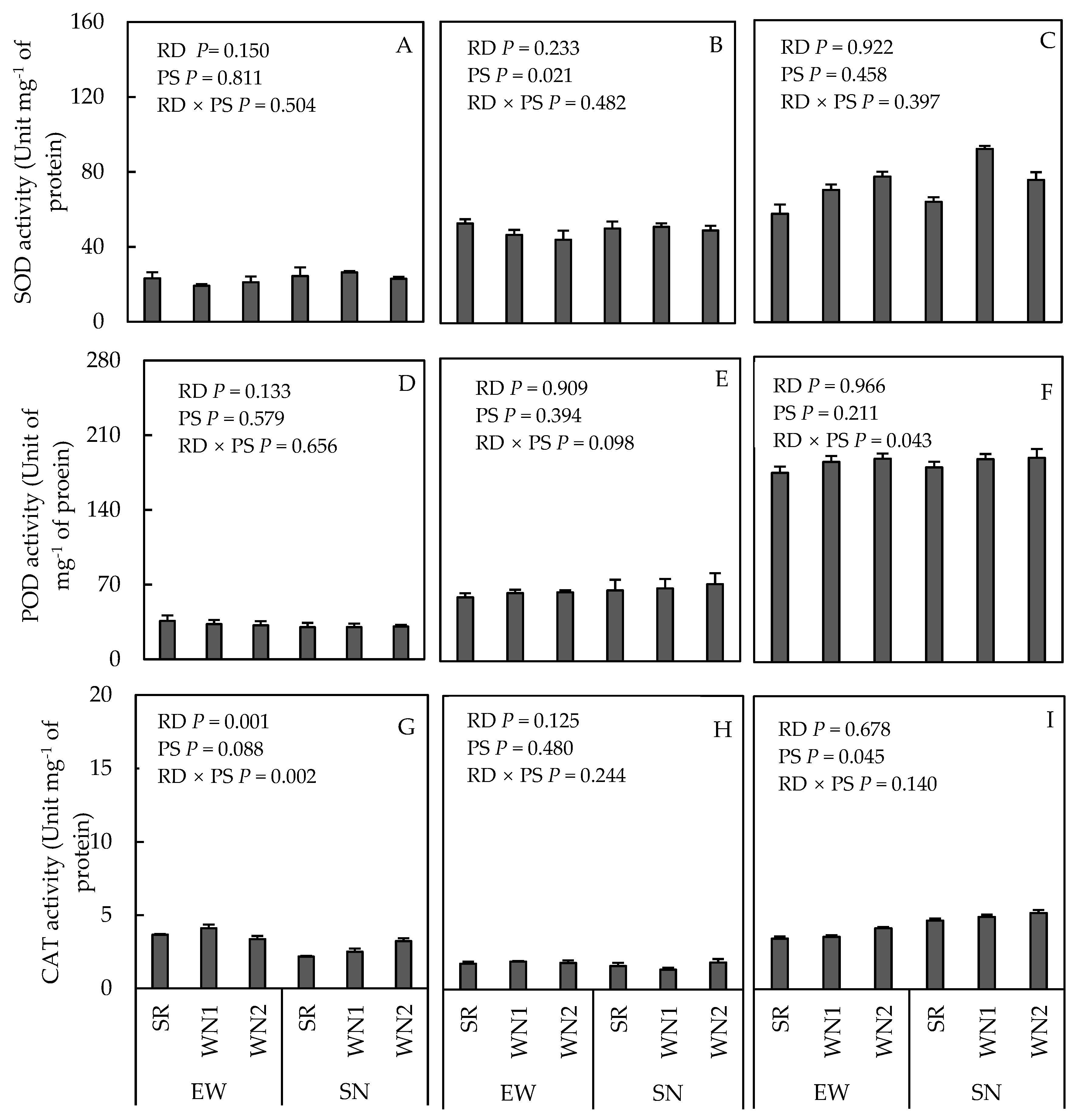
3.2. SOD, POD, and CAT Activities
3.3. ABA Concentration
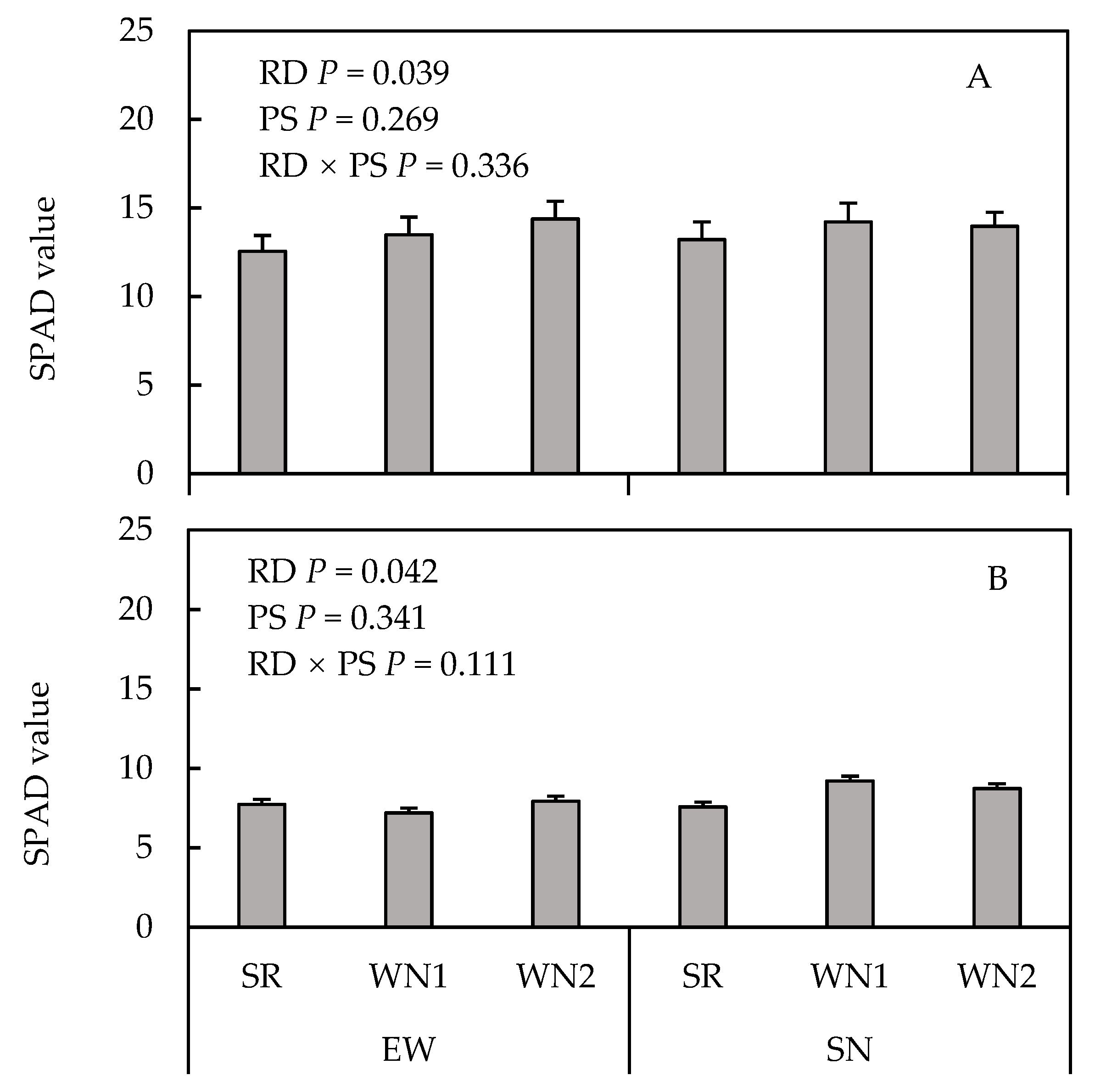
3.4. Chlorophyll Content of Husk Leaves
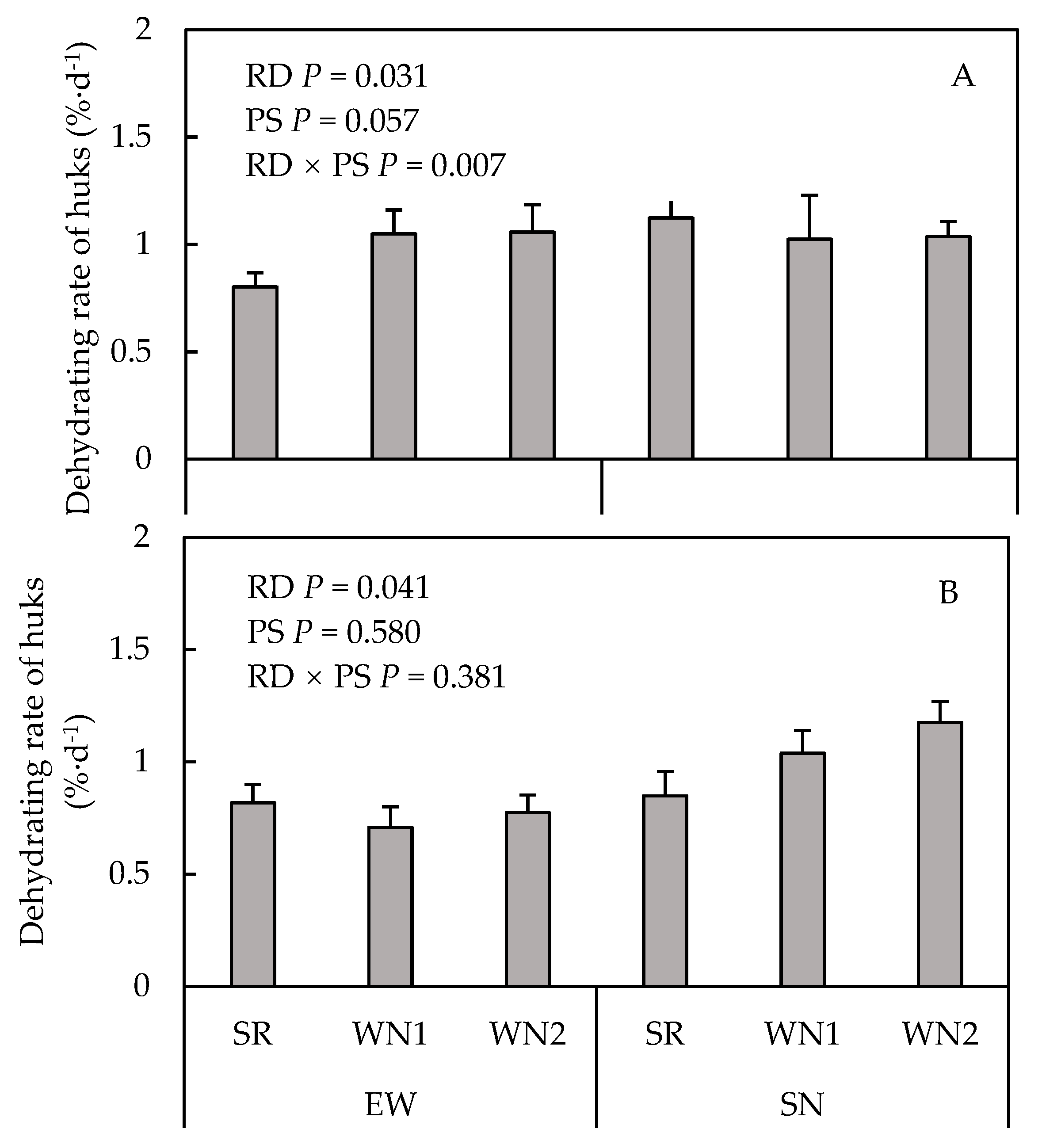
3.5. Dehydrating Rate of Husk Leaves
3.6. Correlation Analysis
3.7. Grain Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhou, Y.; Gu, W.R.; Lie, C.F.; Liu, X.S.; Wang, Y.B.; Li, J.; Li, C.F.; Wei, S. Relationships among grain filling, dehydration characteristics and hormones in spring maize with different maturities in Heilongjiang Province of China. Pak. J. Agric. Sci. 2019, 56, 531–540. [Google Scholar]
- Liu, S.Q.; Li, X.N.; Zhu, X.C.; Song, F.B. Tensile properties of seminal and nodal roots and their relationship with the root diameter and planting density of maize (Zea mays). Crop Pasture Sci. 2018, 69, 717–723. [Google Scholar] [CrossRef]
- Testa, G.; Reyneri, A.; Blandino, M. Maize grain yield enhancement through high plant density cultivation with different inter-row and intra-row spacings. Eur. J. Agron. 2016, 72, 28–37. [Google Scholar] [CrossRef]
- Li, J.; Xie, R.Z.; Wang, K.R.; Ming, B.; Guo, Y.Q.; Zhang, G.Q.; Li, S.K. Variation in maize dry matter, harvest index, and grain yield with plant density. Agron. J. 2015, 107, 829–834. [Google Scholar] [CrossRef]
- Borrás, L.; Maddonni, G.A.; Otegui, M.E. Leaf senescence in maize hybrids: Plant population, row spacing and kernel set effects. Field Crops Res. 2003, 82, 13–26. [Google Scholar] [CrossRef]
- Liu, T.N.; Wang, Z.L.; Cai, T. Canopy apparent photosynthetic characteristics and yield of two spike-type wheat cultivars in response to row spacing under high plant density. PLoS ONE 2016, 11, e0148582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, H.; Shi, D.F.; Shi, W.J.; Ban, X.B.; Chen, Y.C.; Ren, W.; Chen, F.J.; Mi, G.H. Nutrient accumulation and remobilization in relation to yield formation at high planting density in maize hybrids with different senescent characters. Arch. Agron. Soil Sci. 2020. [Google Scholar] [CrossRef]
- Avila-Ospina, L.; Moison, M.; Yoshimoto, K.; Masclaux-Daubresse, C. Autophagy, plant senescence, and nutrient recycling. J. Exp. Bot. 2014, 65, 3799–3811. [Google Scholar] [CrossRef] [Green Version]
- Diaz, C.; Lemaître, T.; Christ, A.; Azzopardi, M.; Kato, Y.; Sato, F.; Morot-Gaudry, J.-F.; Le Dily, F.; Masclaux-Daubresse, C. Nitrogen recycling and remobilization are differentially controlled by leaf senescence and development stage in Arabidopsis under low nitrogen nutrition. Plant Physiol. 2008, 147, 1437–1449. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowska, N.; Wilmowicz, E.; Marzec-Schmidt, K.; Ludwików, A.; Bagniewska-Zadworna, A. Abscisic acid and jasmonate metabolisms are jointly regulated during senescence in roots and leaves of Populus trichocarpa. Int. J. Mol. Sci. 2020, 21, 2042. [Google Scholar] [CrossRef] [Green Version]
- San-Oh, Y.; Sugiyama, T.; Yoshita, D.; Ookawa, T.; Hirasawa, T. The effect of planting pattern on the rate of photosynthesis and related processes during ripening in rice plants. Field Crop. Res. 2006, 96, 113–124. [Google Scholar] [CrossRef]
- Ren, B.Z.; Hu, J.; Zhang, J.W.; Dong, S.T.; Liu, P.; Zhao, B. Effect of urea mixed with nitrapyrin on leaf photosynthetic and senescence characteristics of summer maize (Zea mays. L.) waterlogged in the field. JIA 2020, 19, 1586–1595. [Google Scholar] [CrossRef]
- Zakari, S.A.; Asad, M.A.U.; Han, Z.Y.; Zhao, Q.; Cheng, F.M. Relationship of nitrogen deficiency-induced leaf senescence with ROS generation and ABA concentration in rice flag leaves. J. Plant Growth Regul. 2020. [Google Scholar] [CrossRef]
- Griffiths, C.A.; Gaff, D.F.; Neale, A.D. Drying without senescence in resurrection plants. Front. Plant Sci. 2014, 36, 36. [Google Scholar] [CrossRef] [Green Version]
- Balkcom, K.S.; Satterwhite, J.L.; Arriaga, F.J.; Price, A.J.; Van Santen, E. Conventional and glyphosate-resistant maize yields across plant densities in single- and twin-row configurations. Field Crop Res. 2011, 120, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Sato, H.; Sawada, O.; Sendo, S. Husk leaves contribution to dry matter and grain production as well as n distribution in flint corn (Zea mays L.) genotypes differing in husk leaf area. Soil Sci. Plant Nutr. 1995, 41, 587–596. [Google Scholar] [CrossRef]
- Fujita, K.; El-Shemy, H.; Sakurail, N.; Sendo, S. Sugar metabolism in expanding husk leaf of flint corn (Zea mays L.) genotypes differing in husk leaf size. Plant Nutr. 2001, 92, 278–279. [Google Scholar]
- Kang, M.S.; Zuber, M.S. Combining ability for grain moisture, husk moisture, and maturity in maize with yellow and white endosperms. Crop Sci. 1989, 29, 689–692. [Google Scholar] [CrossRef]
- Li, J.; Xie, R.Z.; Wang, K.R.; Hou, P.; Ming, B.; Zhang, G.Q.; Liu, G.Z.; Wu, M.; Yang, Z.S.; Li, S.K. Response of canopy structure, light interception and grain yield to plant density in maize. J. Agric. Sci. 2018, 156, 1–10. [Google Scholar] [CrossRef]
- Luqman, L.; Hussain, Z.; Ilyas, M.; Khan, I.A.; Bakht, T. Influence of sowing orientation and intercropping of chilies on onion yield and its associated weeds in Peshawar, Pakistan. Pak. J. Bot. 2020, 52, 95–100. [Google Scholar] [CrossRef]
- Souza, C.R.D.; Mota, R.V.D.; Silva, C.P.C.; Raimundo, R.H.P.; Peregrino, I. Row orientation effects on Syrah grapevine performance during winter growing season. Rev. Ceres 2019, 66, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Piao, L.; Qi, H.; Li, C.F.; Zhao, M. Optimized tillage practices and row spacing to improve grain yield and matter transport efficiency in intensive spring maize. Field Crop Res. 2016, 198, 258–268. [Google Scholar] [CrossRef]
- Teasdale, J.R. Influence of narrow row/high population corn (Zea mays) on weed control and light transmittance. Weed Technol. 1995, 9, 113–118. [Google Scholar] [CrossRef]
- Robles, M.; Ciampitti, I.A.; Vyn, T.J. Responses of maize hybrids to twin-row spatial arrangement at multiple plant densities. Agron. J. 2012, 104, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.Q.; Wang, Y.; Song, F.B.; Qi, X.N.; Li, X.N.; Zhu, X.C. Responses of leaf architecture traits and yield in maize to different row orientation and planting patterns in northeast China. Rom. Agric. Res. 2017, 34, 1–12. [Google Scholar]
- Tian, C.; Han, J.; Li, J.; Zhen, G.; Liu, Y.; Lu, Y.; Wang, Y.; Wang, Y. Effects of row direction and row spacing on maize leaf senescence. PLoS ONE 2019, 14, e0215330. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide dismutase. Annu. Rev. Biochem. 1975, 44, 147–159. [Google Scholar] [CrossRef]
- Kumar, K.B.; Khan, P.A. Peroxidase and polyphenol oxidase in excised ragi (Eleusine coracana cv PR 202) leaves during senescence [millets]. Indian J. Exp. Biol. 1982, 20, 412–416. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Lester, P., Ed.; Academic Press: San Diego, CA, USA, 1984; pp. 121–126. [Google Scholar]
- Asch, F. Determination of Abscisic Acid by Indirect Enzyme Linked Immuno Sorbent Assay (ELISA); Technical Report; The Royal Veterinary and Agricultural University, Laboratory for Agrohydrology and Bioclimatology, Department of Agricultural Sciences: Frederiksberg, Denmark, 2000. [Google Scholar]
- Bagniewska-Zadworna, A.; Stelmasik, A.; Minicka, J. From birth to death- Populus trichocarpa fibrous roots functional anatomy. Biol. Plant 2014, 58, 551–560. [Google Scholar] [CrossRef]
- Wojciechowska, N.; Marzec-Schmidt, K.; Kalemba, E.M.; Zarzyńska-Nowak, A.; Jagodziński, A.M.; Bagniewska-Zadworna, A. Autophagy counteracts instantaneous cell death during seasonal senescence of the fine roots and leaves in Populus trichocarpa. BMC Plant Biol. 2018, 18, 260. [Google Scholar] [CrossRef]
- Luoni, S.B.; Astigueta, F.H.; Nicosia, S.; Moschen, S.; Fernandez, P.; Heinz, R. Transcription factors associated with leaf senescence in crops. Plants 2019, 8, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Marquez, F.J.; Nishio, N.; Nagai, S. Promotive effects of 5-aminolevulinic acid on the growth and photosynthesis of Spirulina platensis. Ferm. Bioeng. 1995, 79, 453–457. [Google Scholar] [CrossRef]
- Moreira, A.; Moraes, L.A.C.; Schroth, G.; Mandarino, J.M.G. Effect of nitrogen, row spacing, and plant density on yield, yield components, and plant physiology in soybean-wheat intercropping. Agron. J. 2015, 107, 2162–2170. [Google Scholar] [CrossRef]
- Patil, B.; Kumar, V.; Merwade, M.N. Effect of inter row spacing and fertilizer levels on crop growth, seed yield and seed quality of perennial fodder sorghum cv. CoFS-29. Range Manag. Agrofor. 2018, 39, 59–64. [Google Scholar]
- Arafat, A.H.A.L.; Mostafa, M.G.; Rahman, M.M.; Abdel-Farid, A.B.; Tran, L.S.P. Extracts from yeast and carrot roots enhance maize performance under seawater-induced salt stress by altering physio-biochemical characteristics of stressed plants. J. Plant Growth Regul. 2019, 38, 966–979. [Google Scholar]
- Li, X.; Li, J.H.; Wang, W.; Chen, N.Z.; Ma, T.S.; Xi, Y.N.; Zhang, X.L.; Lin, H.F.; Bai, Y.; Huang, S.J.; et al. ARP2/3 complex-mediated actin dynamics is required for hydrogen peroxide-induced stomatal closure in Arabidopsis. Plant Cell Environ. 2014, 37, 1548–1560. [Google Scholar] [CrossRef]
- Shimizu, M.; Meguro, A.; Hasegawa, S.; Nishimura, T.; Kunoh, H. Disease resistance induced by nonantagonistic endophytic Streptomyces, spp. on tissue-cultured seedlings of rhododendron. J. Gen. Plant Pathol. 2006, 6, 351–354. [Google Scholar] [CrossRef]
- White, A.J.; Critchley, C. Rapid light curves: A new fluorescence method to assess the state of the photosynthetic apparatus. Photosynth Res. 1999, 1, 63–72. [Google Scholar] [CrossRef]
- Zhan, X.M.; Han, X.R.; Yang, J.F.; Wang, S.; Gao, M.; Zhao, L.Y. Effects of different fertilization treatments on leaf protective enzyme activity and membrane lipid peroxidation in late growth stage of maize. Maize Sci. 2007, 1, 123–127. [Google Scholar]
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Responses of carbon metabolism and antioxidant system of summer maize to waterlogging at different stages. J. Agron. Crop Sci. 2018, 204, 505–514. [Google Scholar] [CrossRef]
- Plett, S. Corn kernel breakage as a function of grain moisture at harvest in a prairie environment. Can. J. Plant Sci. 1994, 74, 543–544. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.X.; He, J.Y. Response characteristics between drip irrigation cotton chlorophyll and ABA under the different conditions of nitrogen fertilizers. Xinjiang Agric. Sci. 2015, 52, 995–1001. [Google Scholar]
- Wan, Z.H.; Ren, B.Z.; Zhao, B.; Liu, P.; Zhang, J.W. Grain filling, dehydration characteristics and changes of endogenous hormones of summer maize hybrids differing in maturities. Acta Agron. Sin. 2019, 45, 1446–1453. [Google Scholar]
- Cavalieri, A.J.; Smith, O.S. Grain filling and field drying of a set of maize hybrid released from 1930 to 1982. Crop Sci. 1985, 25, 856–860. [Google Scholar] [CrossRef]
- Wall, G.W.; Kanemasu, E.T. Carbon dioxide exchange rates in wheat canopies. Part I. Influence of canopy geometry on trends in leaf area index, light interception and instantaneous exchange rates. Agric. For. Meteorol. 1990, 49, 81–102. [Google Scholar] [CrossRef]
- Widdicombe, W.D.; Thelen, K.D. Row width and plant density effects on corn grain production in the Northern corn belt. Agron. J. 2002, 94, 1020–1023. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Row Direction | Plant Spacing |
|---|---|
| SR (single rows spaced at 65 cm) | |
| EW (east to west) | WN1 (40 cm twin rows spaced at 90 cm between the paired rows of narrow–wide rows) |
| SN (south to north) | WN2 (40 cm twin rows spaced at 160 cm between the paired rows of narrow–wide rows |
| Physiological Index | Significance Level at P ≤ 0.05 |
|---|---|
| SOD activities | * |
| POD activities | * |
| CAT activities | * |
| ABA concentration | * |
| SPAD value | * |
| Dehydrating rate | * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Gu, Y.; Wang, X.; Li, X.; Wang, Y. Husk Leaf Senescence Characteristics of Spring Maize (Zea mays L.) Cultivated in Two Row Directions and Three Plant Spacings in Northeast China. Agronomy 2020, 10, 1216. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081216
Liu S, Gu Y, Wang X, Li X, Wang Y. Husk Leaf Senescence Characteristics of Spring Maize (Zea mays L.) Cultivated in Two Row Directions and Three Plant Spacings in Northeast China. Agronomy. 2020; 10(8):1216. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081216
Chicago/Turabian StyleLiu, Shengqun, Yan Gu, Xiaobo Wang, Xiangnan Li, and Yang Wang. 2020. "Husk Leaf Senescence Characteristics of Spring Maize (Zea mays L.) Cultivated in Two Row Directions and Three Plant Spacings in Northeast China" Agronomy 10, no. 8: 1216. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081216