Plant Sex Prediction Using Genetic Markers in Cultivated Yams (Dioscorea rotundata Poir.) in Benin

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction, Amplification of the Genetic Markers and Sequencing
2.3. Data Analysis
3. Results
3.1. Floral Characteristics of Beninese Yam Cultivars
3.2. Use of the Marker sp16 to Discriminate Sex in Beninese Yam Cultivars
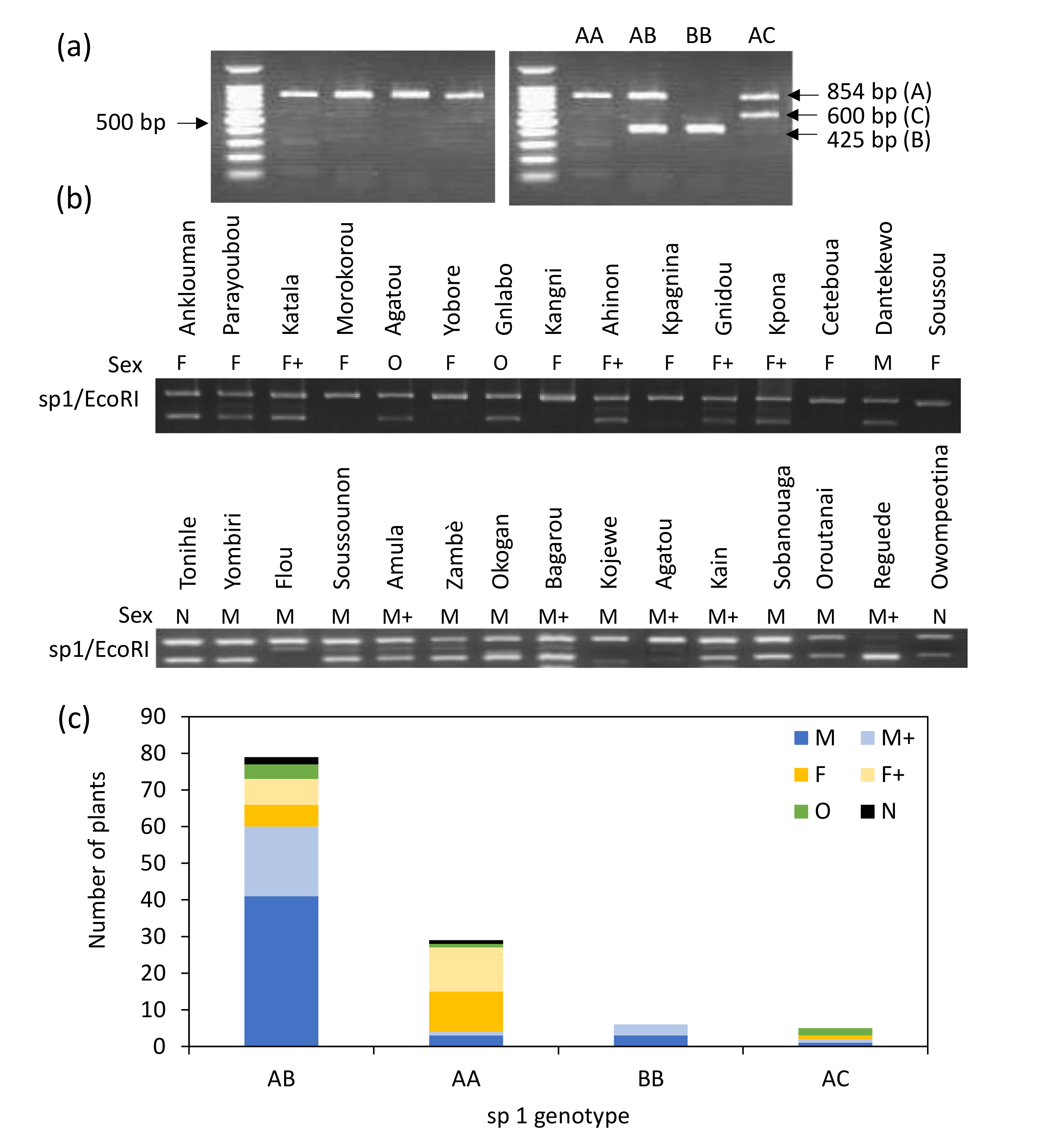
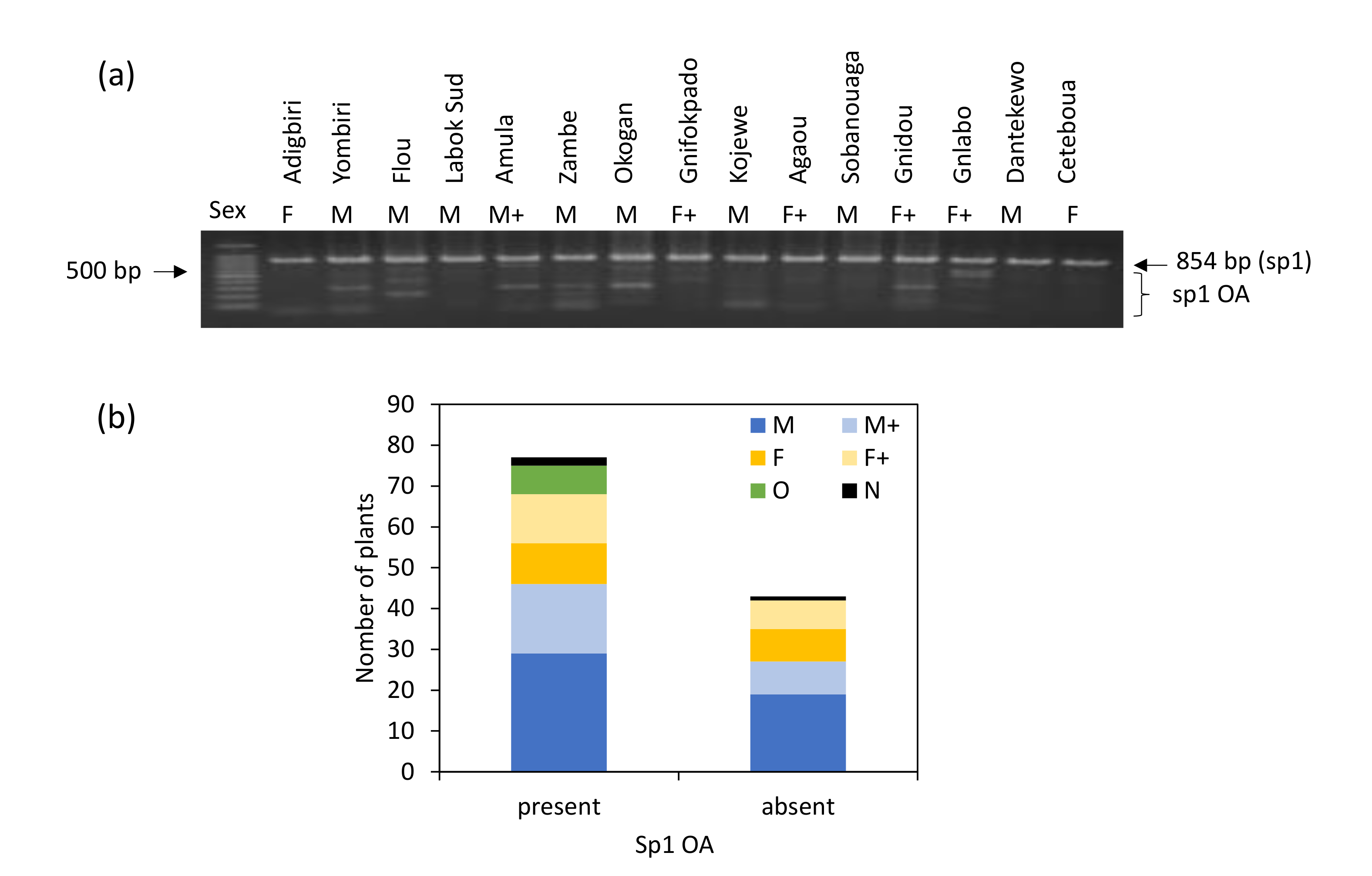
3.3. Use of the Marker sp1 to Discriminate Sex in Beninese Yam Cultivars
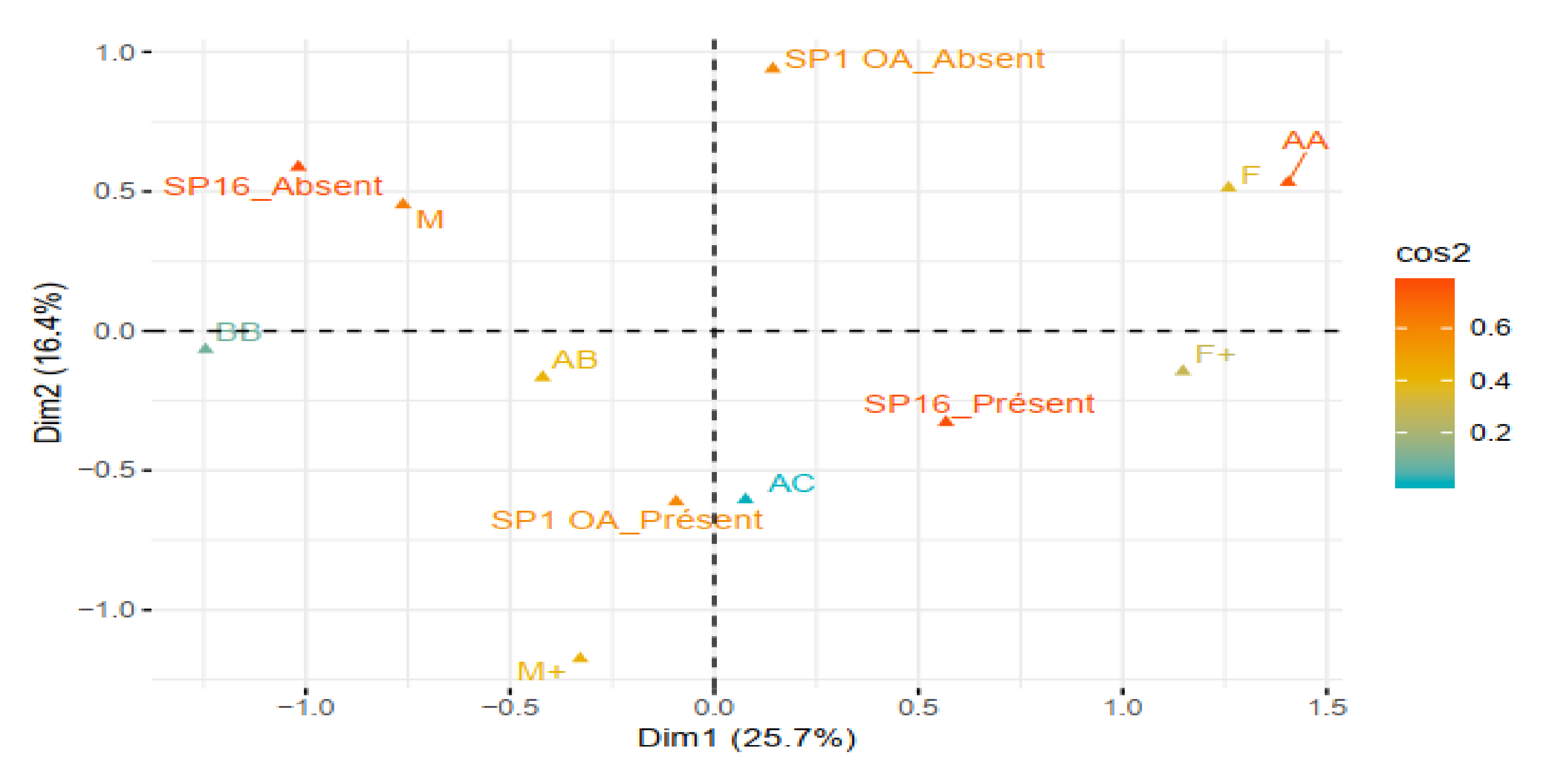
3.4. Relative Importance of sp16 and sp1 Markers in Sex Identification in Beninese Cultivars
3.5. Ploidy Level and Sex Expression
4. Discussion
4.1. Discriminating Power of the sp16 Marker and Link between the W Allele and Sex Determination
4.2. Homo/heterogametic System in D. rotundata
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coursey, D.G. The Civilizations of the Yam: Interrelationships of Man and Yams in Africa and the Indo-Pacific Region. Archaeol. Phys. Anthr. Ocean. 1972, 7, 215–233. [Google Scholar]
- Tostain, S.; Agbangla, C.; Scarcelli, N.; Mariac, C.; Daïnou, O.; Berthaud, J.; Pham, J.-L. Genetic diversity analysis of yam cultivars (Dioscorea rotundata Poir.) in Benin using simple sequence repeat (SSR) markers. Plant Genet. Resour. 2007, 5, 71–81. [Google Scholar] [CrossRef]
- Fakorede, J.; Faouziath, S.; Loko Yêyinou, L.E.; Dassou, G.A.T.; Tchekessi, C.; Ogundipe, O.; Bokossa, Y.I.; Dansi, A.A. Evaluation of proximate, sugars and mineral compositions of 48 yam (Dioscorea rotundata) cultivars used as parents in a breeding program in Republic of Benin. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 1663–1682. [Google Scholar] [CrossRef]
- Dansi, A.; Pillay, M.; Mignouna, H.D.; Daïnou, O.; Mondeil, F.; Moutaïrou, K. Ploidy level of the cultivated yams (Dioscorea cayenensis/D. rotundata complex) from Benin Republic as determined by chromosome counting and flow cytometry. Afr. Crop Sci. J. 2000, 8. [Google Scholar] [CrossRef]
- Yolou, M.; Zoundjihekpon, J.; Assaba, E.I.; Anizehou, I.; Akoegninnou, A. La floraison des ignames africaines cultivées (D. cayenensis—D. rotundata et D. dumetorum) dans les champs des producteurs du Centre-Bénin. J. Appl. Biosci. 2015, 91, 8480–8492. [Google Scholar] [CrossRef] [Green Version]
- Zoundjihekpon, J.; Essad, S.; Toure, B. Dénombrement chromosomique dans dix groupes variétaux du Complexe Dioscorea cayenensis-rotundata. Cytologia 1990, 55, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.K. Dioscorea rotundata. In Edible Medicinal and Non-Medicinal Plants: Volume 10, Modified Stems, Roots, Bulbs; Lim, T.K., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2016; pp. 291–300. ISBN 978-94-017-7276-1. [Google Scholar]
- Darkwa, K.; Olasanmi, B.; Asiedu, R.; Asfaw, A. Review of empirical and emerging breeding methods and tools for yam (Dioscorea spp.) improvement: Status and prospects. Plant Breed. 2020, 139, 474–497. [Google Scholar] [CrossRef] [Green Version]
- Girma, G.; Natsume, S.; Carluccio, A.V.; Takagi, H.; Matsumura, H.; Uemura, A.; Muranaka, S.; Takagi, H.; Stavolone, L.; Gedil, M.; et al. Identification of candidate flowering and sex genes in white Guinea yam (D. rotundata Poir.) by SuperSAGE transcriptome profiling. PLoS ONE 2019, 14, e0216912. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 15 April 2019).
- Loko, Y.L.; Dansi, A.; Agre, A.P.; Akpa, N.; Dossou-Aminon, I.; Assogba, P.; Dansi, M.; Akpagana, K.; Sanni, A. Perceptions paysannes et impacts des changements climatiques sur la production et la diversité variétale de l’igname dans la zone aride du nord-ouest du Bénin. Int. J. Biol. Chem. Sci. 2013, 7, 672–695. [Google Scholar] [CrossRef] [Green Version]
- Degras, L. L’ Igname, plante à tubercule tropicale. In Techniques Agricoles et Productions Tropicales; G.P. Maisonneuve & Larose: Paris, France, 1986; ISBN 978-2-7068-0915-6. [Google Scholar]
- Bousalem, M.; Dallot, S.; Guyader, S. The use of phylogenetic data to develop molecular tools for the detection and genotyping of Yam mosaic virus. Potential application in molecular epidemiology. J. Virol. Methods 2000, 90, 25–36. [Google Scholar] [CrossRef]
- Egesi, C.N.; Onyeka, T.J.; Asiedu, R. Environmental stability of resistance to anthracnose and virus dyases of water yam (Dioscorea alata). Afr. J. Agric. Res. 2009, 4, 113–118. [Google Scholar]
- Amusa, N.A.; Adigbite, A.A.; Muhammed, S.; Baiyewu, R.A. Yam diseases and its management in Nigeria. Afr. J. Biotechnol. 2003, 2, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Ogaraku, A.O.; Usman, H.O. Storage Rot of Some Yams (Dioscorea spp) in Keffi and Environs, Nasarawa State, Nigeria. Patnsuk J. 2008, 4, 22–27. [Google Scholar]
- Asala, S.; Alegbejo, M.D.; Kashina, B.; Banwo, O.O.; Asiedu, R.; Kumar, P.L. Distribution and incidence of viruses infecting yam (Dioscorea spp.) in Nigeria. Glob. J. Biotechnol. Biosci. 2012, 1, 163–167. [Google Scholar]
- Ayisah, K.D.; Gumedzoe, Y.M.D. Genetic diversity among yam mosaic virus (YMV) isolates infecting yam of the complex Dioscorea cayenensis rotundata in Togo. Int. J. Biol. Chem. Sci. 2012, 6, 1090–1101. [Google Scholar] [CrossRef]
- Thouvenel, J.C.; Dumont, R. Perte de rendement de l’igname infectée par le virus de la mosaique en Côte d’Ivoire. L’Agron. Trop. 1975, 45, 125–129. [Google Scholar]
- Yolou, M.; Anizehou, I.; Dossou-Yovo, R.; Akoegninnou, A.; Zongo, J.D.; Zoundjihekpon, J. Etat des lieux de la reproduction sexuée des ignames africaines Dioscorea cayenensis–Dioscorea rotundata cultivées au Bénin. Int. J. Biol. Chem. Sci. 2015, 9, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Assaba, E.; Yolou, M.; Totin, V.; Quenum, H.; Zoundjihekpon, J. Caractéristiques des graines d’ignames cultivées du complexe d’espèces Dioscorea cayenensis-Dioscorea rotundata et évolution du taux de germination des graines. J. Afr. Sci. 2018, 14, 198–208. [Google Scholar]
- Tamiru, M.; Natsume, S.; Takagi, H.; White, B.; Yaegashi, H.; Shimizu, M.; Yoshida, K.; Uemura, A.; Oikawa, K.; Abe, A.; et al. Genome sequencing of the staple food crop white Guinea yam enables the development of a molecular marker for sex determination. BMC Biol. 2017, 15, 86. [Google Scholar] [CrossRef] [Green Version]
- Girma, G.; Hyma, K.E.; Asiedu, R.; Mitchell, S.E.; Gedil, M.; Spillane, C. Next-generation sequencing based genotyping, cytometry and phenotyping for understanding diversity and evolution of guinea yams. Theor. Appl. Genet. 2014, 127, 1783–1794. [Google Scholar] [CrossRef]
- Daïnou, O.; Agbangla, C.; Berthaud, J.; Tostain, S. Le nombre chromosomique de base des espèces de Dioscorea constituant la section Enantiophyllum pourrait être égal a X = 20. Quelques preuves. Ann. Sci. Agron. 2002, 3. [Google Scholar] [CrossRef]
- Scarcelli, N.; Daïnou, O.; Agbangla, C.; Tostain, S.; Pham, J.-L. Segregation patterns of isozyme loci and microsatellite markers show the diploidy of African yam Dioscorea rotundata (2n= 40). Theor. Appl. Genet. 2005, 111, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Arnau, G.; Nemorin, A.; Maledon, E.; Abraham, K. Revision of ploidy status of Dioscorea alata L. (Dioscoreaceae) by cytogenetic and microsatellite segregation analysis. Theor. Appl. Genet. 2009, 118, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Cormier, F.; Lawac, F.; Maledon, E.; Gravillon, M.-C.; Nudol, E.; Mournet, P.; Vignes, H.; Chaïr, H.; Arnau, G. A reference high-density genetic map of greater yam (Dioscorea alata L.). Theor. Appl. Genet. 2019, 132, 1733–1744. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.W. Sex Ratio and Sex Determination in Dioscorea. J. Hered. 1966, 57, 95–99. [Google Scholar] [CrossRef]
- Dansi, A.; Mignouna, H.D.; Zoundjihékpon, J.; Sangare, A.; Asiedu, R.; Quin, F.M. Morphological diversity, cultivar groups and possible descent in the cultivated yams (Dioscorea cayenensis/D. rotundata) complex in Benin Republic. Genet. Resour. Crop Evol. 1999, 46, 371–388. [Google Scholar] [CrossRef]
- Yolou, M. Diversité génétique des ignames africaines cultivées (Complexe Dioscorea cayenensis D. rotundata et D. dumetorum) et perception des paysans du Centre-Bénin des questions des droits de propriété intellectuelle. Ph.D. Thesis, Abomey-Calavi University, Cotonou, Benin, 2016; p. 155. [Google Scholar]
- Fulton, T.M.; Chunwongse, J.; Tanksley, S.D. Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Report. 1995, 13, 207–209. [Google Scholar] [CrossRef]
- RDevelopment; C.TEAM 2009. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: http://www.R-project.org (accessed on 30 June 2019).
- Lebot, V.; Abraham, K.; Kaoh, J.; Rogers, C.; Molisalé, T. Development of anthracnose resistant hybrids of the Greater Yam (Dioscorea alata L.) and interspecific hybrids with D. nummularia Lam. Genet. Resour. Crop Evol. 2019, 66, 871–883. [Google Scholar] [CrossRef]
- Hamadina, E.I.; Craufurd, P.Q.; Asiedu, R. Flowering intensity in white yam (Dioscorea rotundata). J. Agric. Sci. 2009, 147, 469–477. [Google Scholar] [CrossRef]
- Nwankwo, I.I.M.; Bassey, E.E. A study of variability and heritability for yield and yield related traits in guinea white yam (Dioscorea rotundata Poir.) genotypes in Umudike, Southeastern Nigeria. Wudpecker J. Agric. Res. 2013, 2, 122–127. [Google Scholar]
- Alam, S.; Shylla, E.; Bora, P.; Saud, B.K. Genetic Variation in Different Cultivars of Greater Yam (Dioscorea alata). J. Root Crops 2014, 40, 1–5. [Google Scholar]
- Terauchi, R.; Kahl, G. Mapping of the Dioscorea tokoro genome: AFLP markers linked to sex. Genome 1999, 42, 752–762. [Google Scholar] [CrossRef]
- Scarcelli, N.; Tostain, S.; Vigouroux, Y.; Agbangla, C.; Daïnou, O.; Pham, J.-L. Farmers’ use of wild relative and sexual reproduction in a vegetatively propagated crop. The case of yam in Benin. Mol. Ecol. 2006, 15, 2421–2431. [Google Scholar] [CrossRef] [PubMed]
- Chaïr, H.; Cornet, D.; Deu, M.; Baco, M.N.; Agbangla, A.; Duval, M.F.; Noyer, J.L. Impact of farmer selection on yam genetic diversity. Conserv. Genet 2010, 11, 2255–2265. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Primer Sequences | Tm | Cycles | Size (bp) |
|---|---|---|---|---|
| Dr-Actine | F: 5′-CAGGGAAAAGATGACCCAAATC-3 R: 5′-CCATCACCAGAATCCAGCAC-3 | 55 °C | 35 | ~250 bp |
| sp1 | F: 5′-GATCTGGCTTCCTCCATCTTG-3′ R: 5′-GCTTGGGTGGTTAGTTTATTGTTTG-3′ | 52 °C | 40 | 854 bp |
| sp16 | F: 5′-AATGTGTTTAACAGGGTGAATTC-3′ R: 5′-GAATTCAGCCGAATATACTTATTC-3′ | 52 °C | 40 | 262 bp |
| Male (M+) | Female (F+) | |||
|---|---|---|---|---|
| sp1OA absent | sp1OA present | sp1OA absent | sp1OA present | |
| sp16 absent | 12.50% | 16.67% | 5.26% | 0.00% |
| sp16 present | 16.67% | 57.17% | 31.58% | 63.16% |
| Cultivar | Ploidy Level | Sex1 | sp16 Marker | sp1 Marker |
|---|---|---|---|---|
| Baridjo | 3X | M | Present | AB |
| Doundoua | 4X | M | Present | AB |
| Owompeotina | 3X | N | Present | AB |
| Alakissa | 4X | M | Present | AB |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denadi, N.; Gandonou, C.; Missihoun, A.A.; Zoundjihékpon, J.; Quinet, M. Plant Sex Prediction Using Genetic Markers in Cultivated Yams (Dioscorea rotundata Poir.) in Benin. Agronomy 2020, 10, 1521. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101521
Denadi N, Gandonou C, Missihoun AA, Zoundjihékpon J, Quinet M. Plant Sex Prediction Using Genetic Markers in Cultivated Yams (Dioscorea rotundata Poir.) in Benin. Agronomy. 2020; 10(10):1521. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101521
Chicago/Turabian StyleDenadi, Narcisse, Christophe Gandonou, Antoine Abel Missihoun, Jeanne Zoundjihékpon, and Muriel Quinet. 2020. "Plant Sex Prediction Using Genetic Markers in Cultivated Yams (Dioscorea rotundata Poir.) in Benin" Agronomy 10, no. 10: 1521. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101521