Combined Linkage Mapping and Genome-Wide Association Study Identified QTLs Associated with Grain Shape and Weight in Rice (Oryza sativa L.)


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mapping Population, Growth Conditions, and Phenotypic Measurements
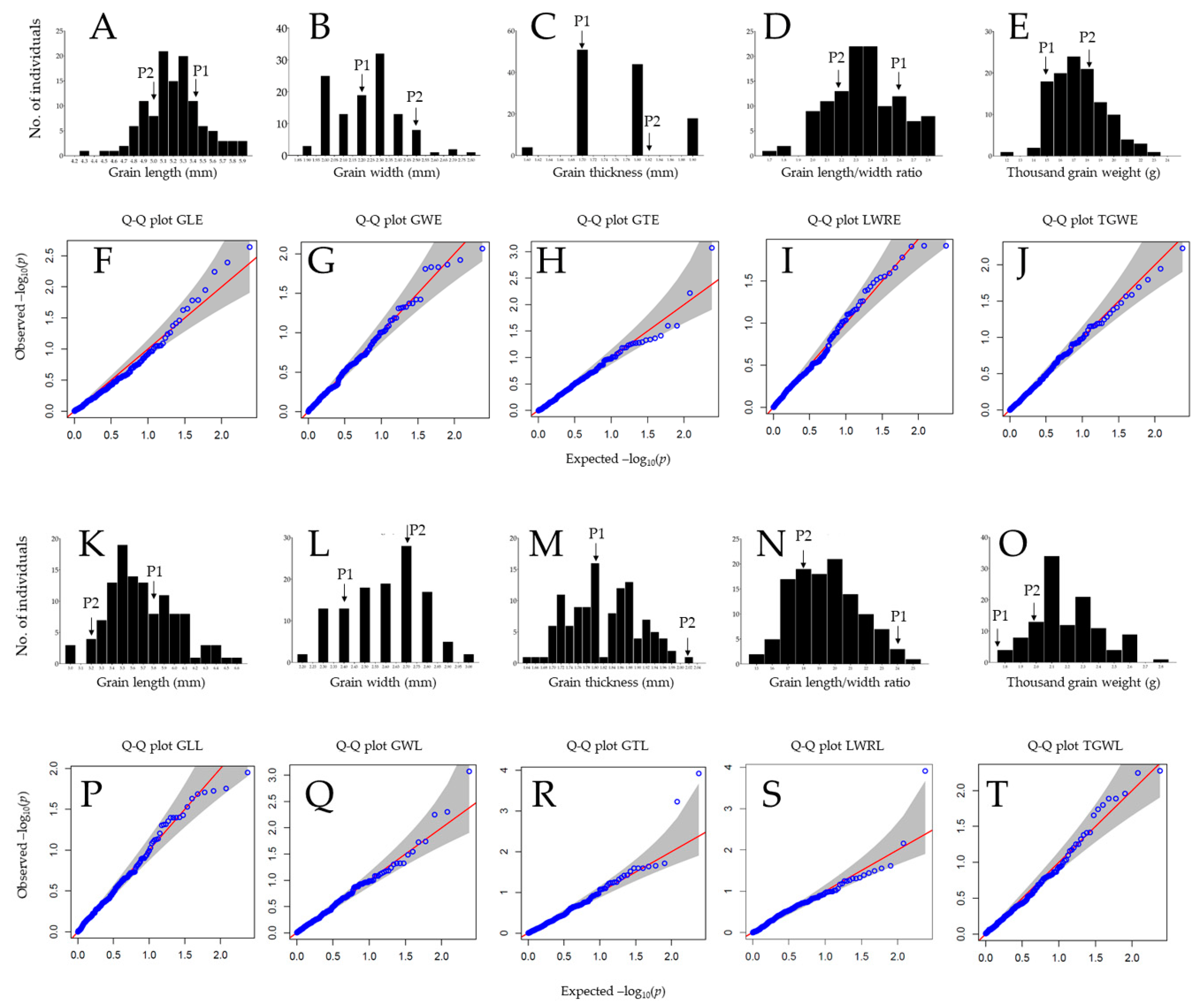
2.2. Frequency Distribution, Quantile–Quantile Plots, Correlation Analysis, and Principal Component Analysis
2.3. Genomic DNA Extraction and Molecular Markers Analysis
2.4. Construction of Linkage Maps, QTL Analysis, and GWAS
3. Results
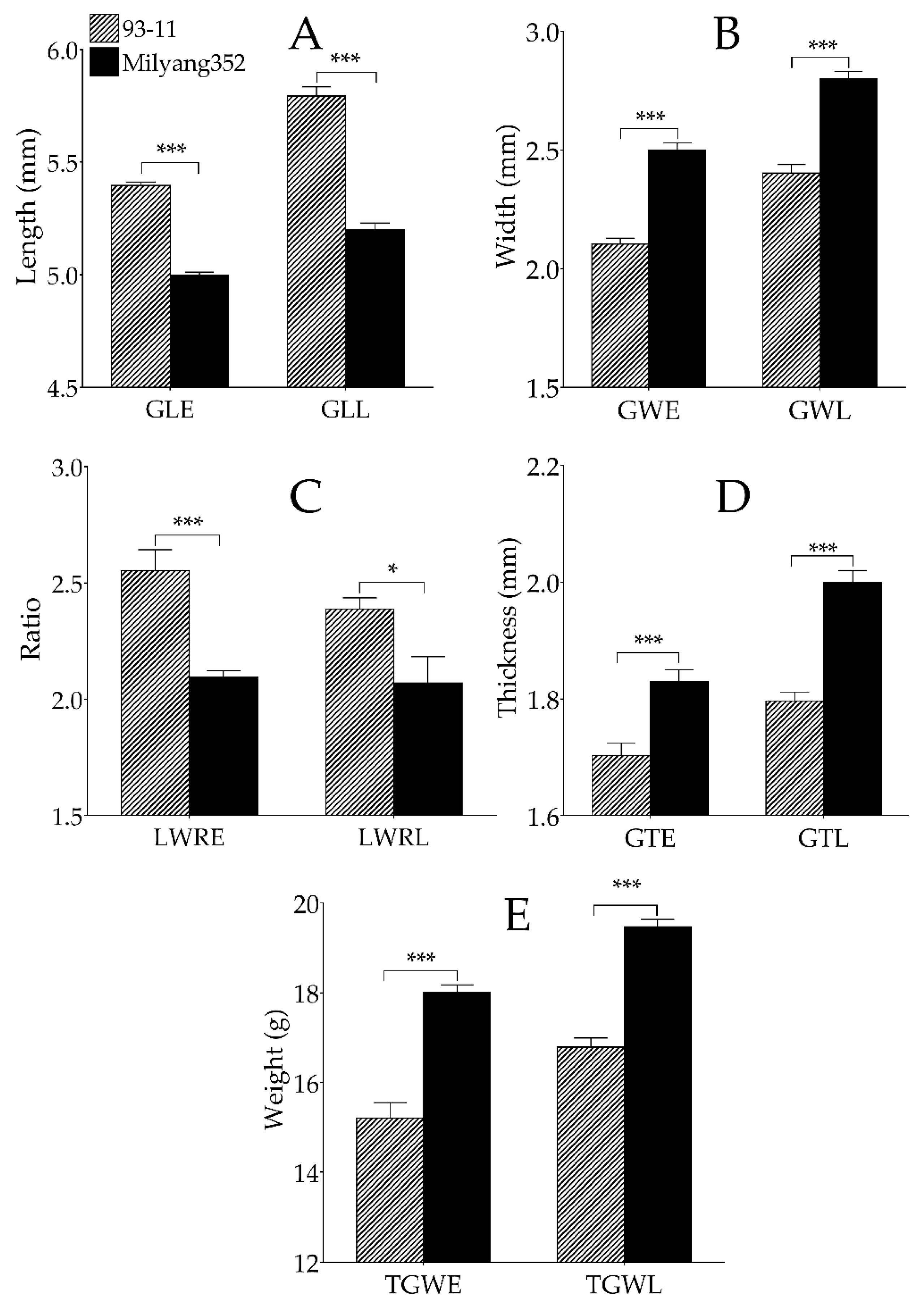
3.1. Differential Grain Traits of Parental Genotypes and DH Population
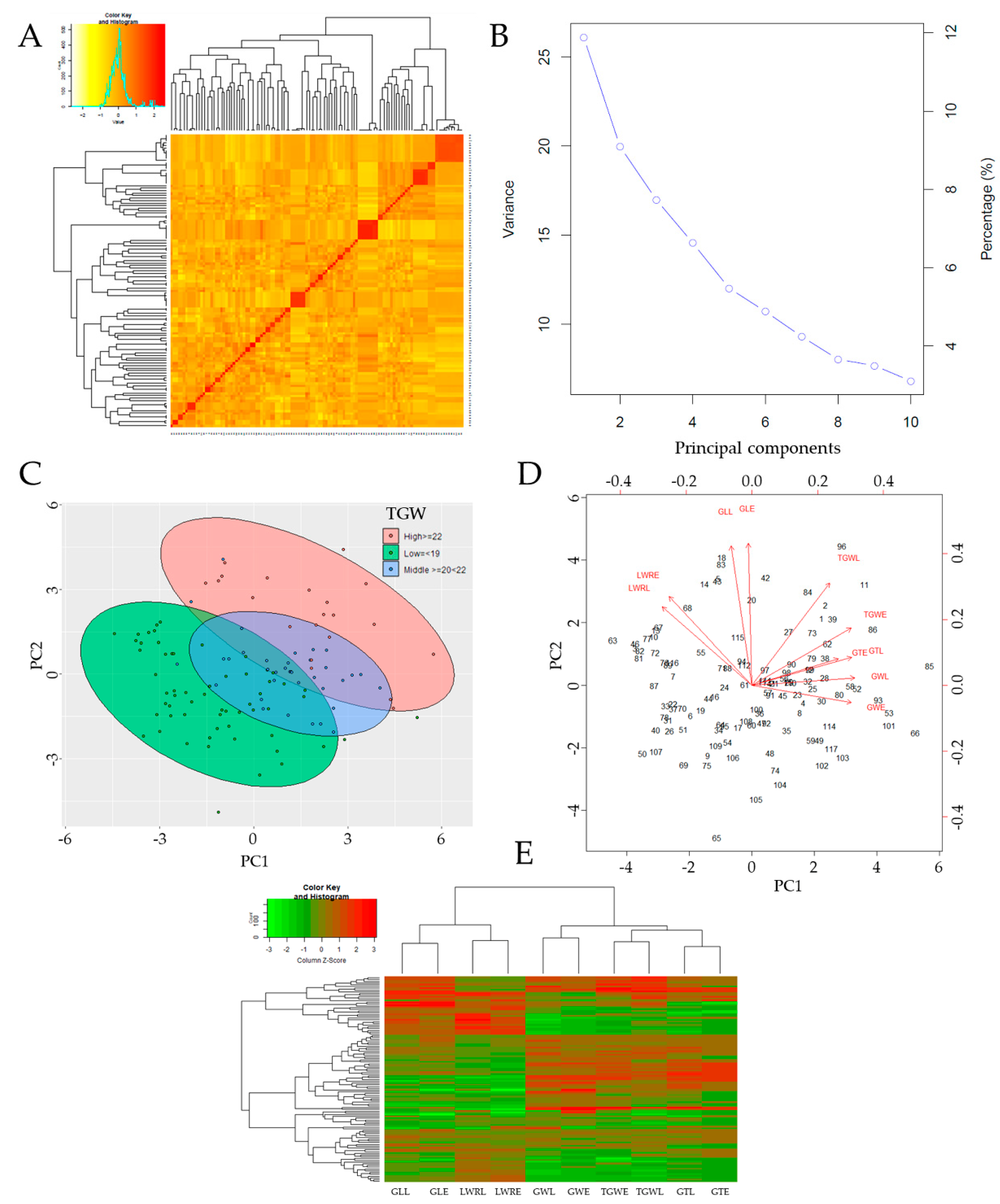
3.2. Relatedness and Principal Component Analysis (PCA)
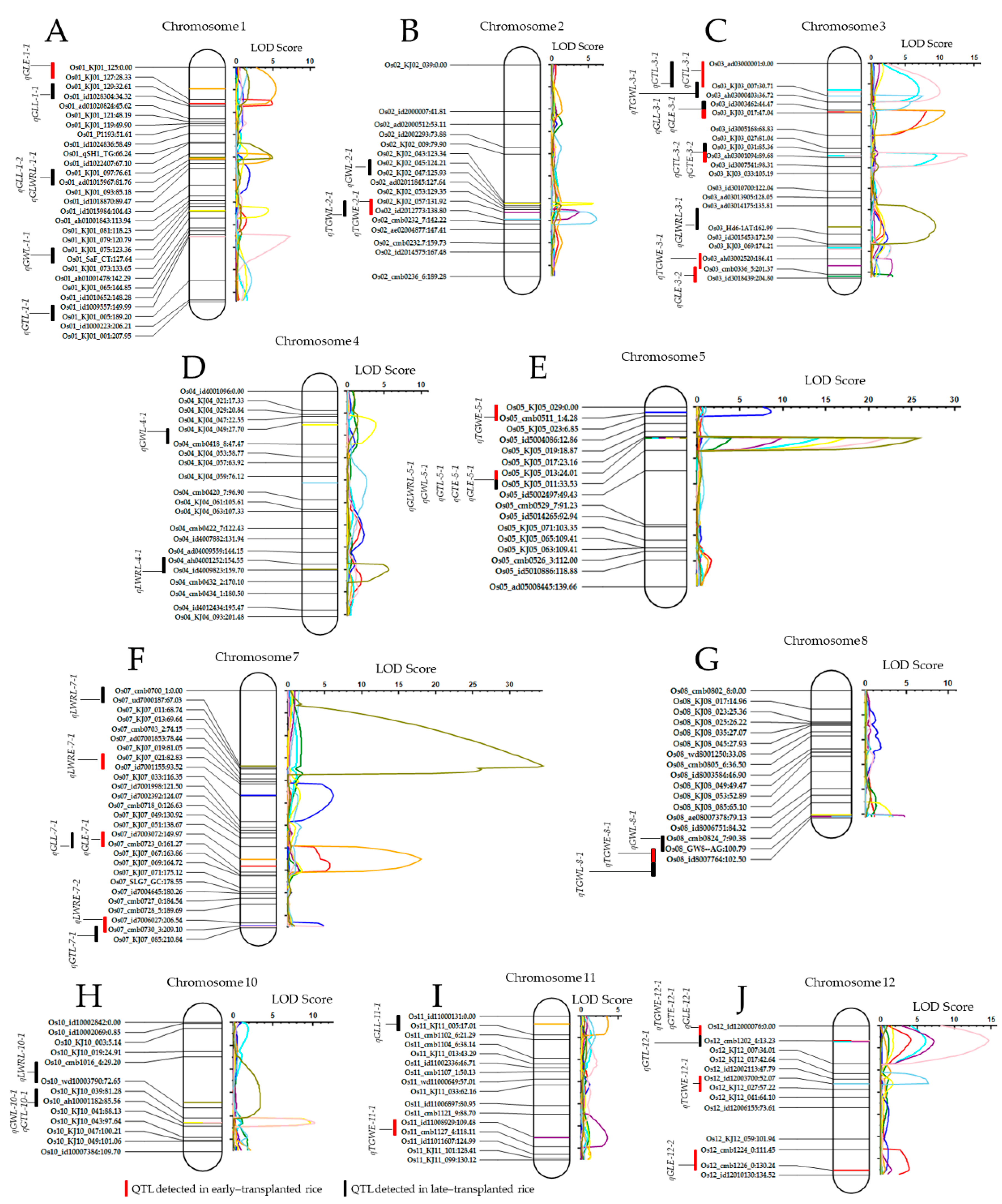
3.3. Detected Quantitative Traits Loci (QTLs) Associated with Rice Grain Traits Mapped on Different Chromosomes
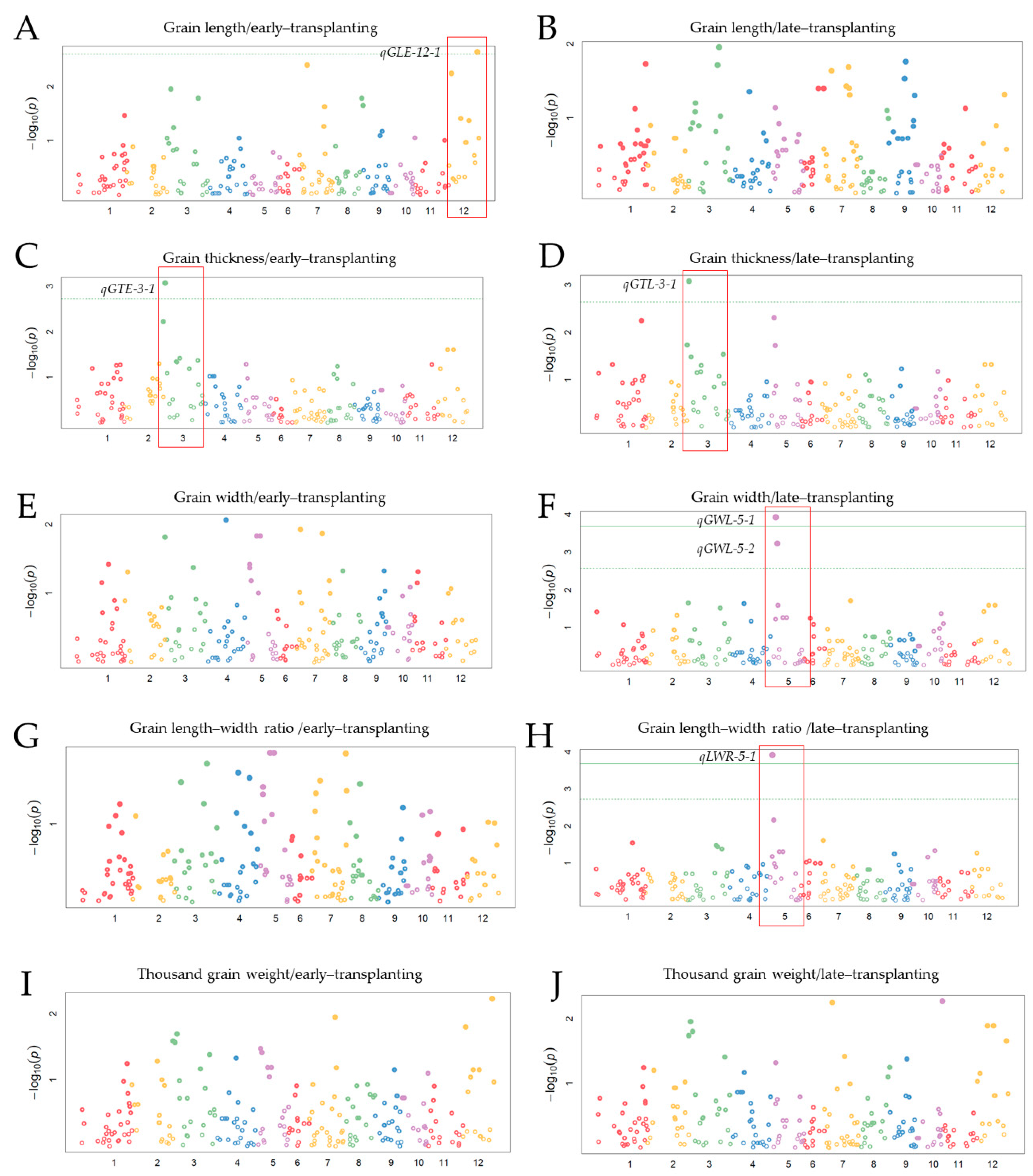
3.4. Detected QTLs by Genome-Wide Association Study (GWAS)
4. Discussion
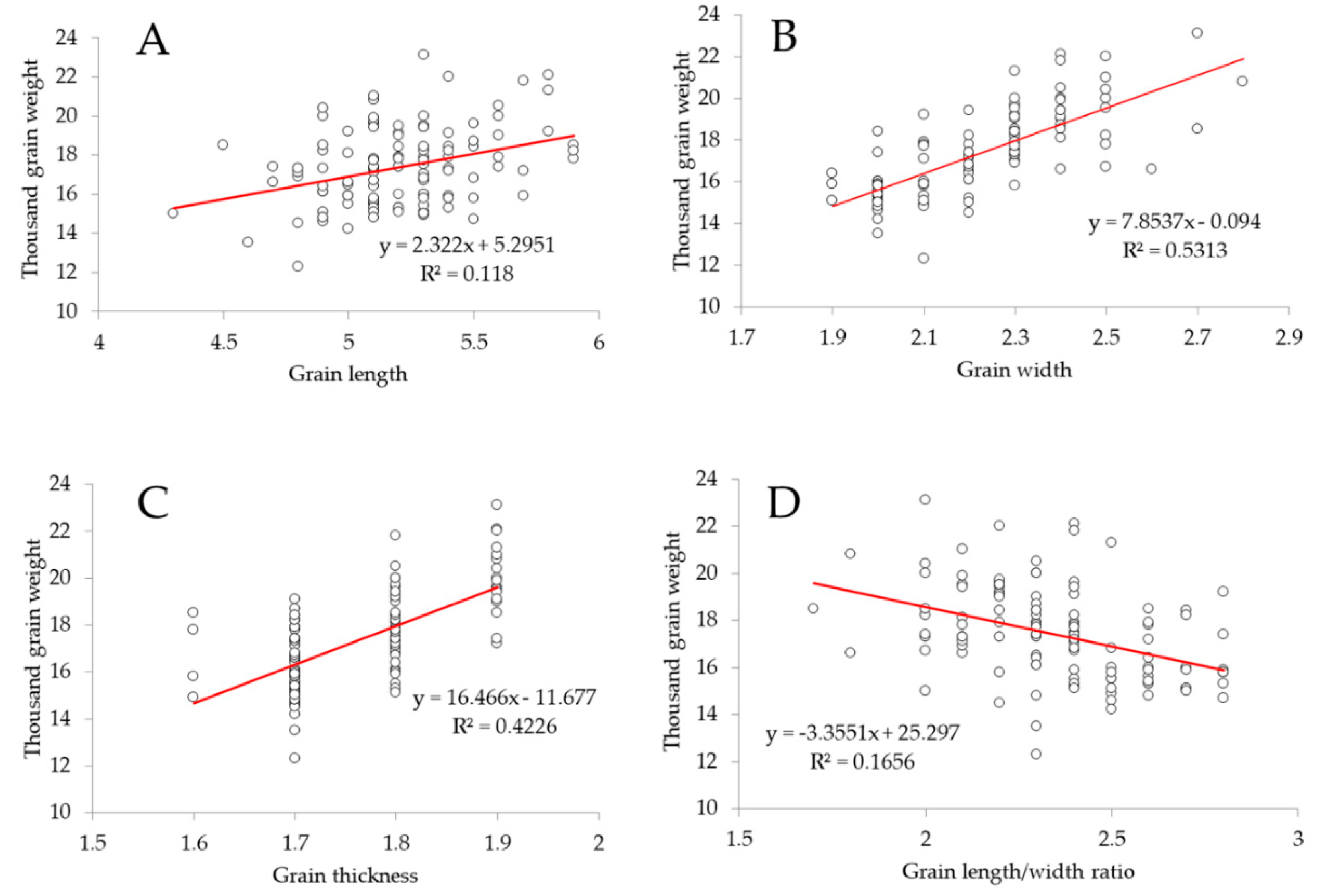
4.1. Grain Width and Thickness are Closely Related to Grain Weight in Rice
4.2. Combined Linkage Map and Genome-Wide Association Study (GWAS) Detected Similar Genetic Loci Associated with Grain Shape and Weight in Rice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yuan, L.; Denning, G.; Mew, T. Hybrid rice breeding for super high yield. In Proceedings of the China-IRRI Dialogue held in Beijing, Beijing, China, 7–8 November 1997; pp. 10–12. [Google Scholar]
- Kumar, A.; Dixit, S.; Ram, T.; Yadaw, R.; Mishra, K.; Mandal, N. Breeding high-yielding drought-tolerant rice: Genetic variations and conventional and molecular approaches. J. Exp. Bot. 2014, 65, 6265–6278. [Google Scholar] [CrossRef] [Green Version]
- Goto, A.; Sasahara, H.; Shigemune, A.; Miura, K. Hokuriku 193: A new high-yielding indica rice cultivar bred in Japan. Jpn. Agric. Res. Q. JARQ 2009, 43, 13–18. [Google Scholar] [CrossRef] [Green Version]
- UN DESA. World Population Projected to Reach 9.8 Billion in 2050, and 11.2 Billion in 2100. 2019. Available online: https://www.un.org/development/desa/en/news/population/world-population-prospects-2017.html (accessed on 9 October 2020).
- He, P.; Li, S.; Qian, Q.; Ma, Y.; Li, J.; Wang, W.; Chen, Y.; Zhu, L. Genetic analysis of rice grain quality. Theor. Appl. Genet. 1999, 98, 502–508. [Google Scholar] [CrossRef]
- Juliano, B.; Duff, B. Rice grain quality as an emerging priority in National rice breeding programmes. In Rice Grain Marketing and Quality Issues; IRRI: Laguna, Philippines, 1991; pp. 55–64. [Google Scholar]
- Hori, K. Genetic dissection and breeding for grain appearance quality in rice. In Rice Genomics, Genetics and Breeding; Springer: New York, NY, USA, 2018; pp. 435–451. [Google Scholar]
- Ahmar, S.; Gill, R.A.; Jung, K.-H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenaerts, B.; Collard, B.C.; Demont, M. Improving global food security through accelerated plant breeding. Plant. Sci. 2019, 287, 110207. [Google Scholar] [CrossRef]
- Bhargava, A.; Srivastava, S. Participatory Plant Breeding: Concept and Applications; Springer: New York, NY, USA, 2019. [Google Scholar]
- Campbell, M.T.; Bandillo, N.; Al Shiblawi, F.R.A.; Sharma, S.; Liu, K.; Du, Q.; Schmitz, A.J.; Zhang, C.; Véry, A.-A.; Lorenz, A.J. Allelic variants of OsHKT1; 1 underlie the divergence between indica and japonica subspecies of rice (Oryza sativa) for root sodium content. PLoS Genet. 2017, 13, e1006823. [Google Scholar] [CrossRef] [PubMed]
- Bush, W.S.; Moore, J.H. Genome-wide association studies. PLoS Comput. Biol. 2012, 8, e1002822. [Google Scholar] [CrossRef] [Green Version]
- Hirschhorn, J.N.; Daly, M.J. Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 2005, 6, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Famoso, A.N.; Zhao, K.; Clark, R.T.; Tung, C.-W.; Wright, M.H.; Bustamante, C.; Kochian, L.V.; McCouch, S.R. Genetic architecture of aluminum tolerance in rice (Oryza sativa) determined through genome-wide association analysis and QTL mapping. PLoS Genet. 2011, 7, e1002221. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.M.; Rai, V. Plant Omics and Crop Breeding; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Kim, S.; Tang, H.; Mardis, E.R. Genome Sequencing Technology and Algorithms; Artech House, Inc.: Norwood, MA, USA, 2007. [Google Scholar]
- Kasahara, M.; Morishita, S. Large-Scale Genome Sequence Processing; Imperial College Press: London, UK, 2006. [Google Scholar]
- Huang, R.; Jiang, L.; Zheng, J.; Wang, T.; Wang, H.; Huang, Y.; Hong, Z. Genetic bases of rice grain shape: So many genes, so little known. Trends Plant Sci. 2013, 18, 218–226. [Google Scholar] [CrossRef]
- Gnanamanickam, S.S. Rice and its importance to human life. In Biological Control of Rice Diseases; Springer: New York, NY, USA, 2009. [Google Scholar]
- Liu, X.; Wei, X.; Sheng, Z.; Jiao, G.; Tang, S.; Luo, J.; Hu, P. Polycomb protein OsFIE2 affects plant height and grain yield in rice. PLoS ONE 2016, 11, e0164748. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Chen, K.; Dong, N.-Q.; Shi, C.-L.; Ye, W.-W.; Gao, J.-P.; Shan, J.-X.; Lin, H.-X. Grain size and number1 negatively regulates the OsMKKK10-OsMKK4-OsMPK6 cascade to coordinate the trade-off between grain number per panicle and grain size in rice. Plant Cell 2018, 30, 871–888. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Miao, J.; Gu, H.; Peng, X.; Leburu, M.; Yuan, F.; Gu, H.; Gao, Y.; Tao, Y.; Zhu, J. Natural variations in SLG7 regulate grain shape in rice. Genetics 2015, 201, 1591–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Xu, R.; Duan, P.; Li, Y. Control of grain size in rice. Plant Reprod. 2018, 31, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef]
- Tan, Y.-F.; Xing, Y.-Z.; Li, J.-X.; Yu, S.-B.; Xu, C.-G.; Zhang, Q. Genetic bases of appearance quality of rice grains in Shanyou 63, an elite rice hybrid. Theor. Appl. Genet. 2000, 101, 823–829. [Google Scholar] [CrossRef]
- Zeng, Y.; Shi, J.; Ji, Z.; Wen, Z.; Liang, Y.; Yang, C. Combination of twelve alleles at six quantitative trait loci determines grain weight in rice. PLoS ONE 2017, 12, e0181588. [Google Scholar] [CrossRef] [Green Version]
- Weng, J.; Gu, S.; Wan, X.; Gao, H.; Guo, T.; Su, N.; Lei, C.; Zhang, X.; Cheng, Z.; Guo, X.; et al. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 2008, 18, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Unnevehr, L.; Duff, B.; Juliano, B. Consumer demand for rice grain quality: Introduction and major findings. In Consumer Demand for Rice Grain Quality; Unnevehr, L.J., Duff, B., Juliano, B.O., Eds.; International Rice Research Institute; International Development Research Center: Manila, Philippines; Ottawa, ON, Canada, 1992; pp. 5–19. [Google Scholar]
- Juliano, B.O.; Villareal, C. Grain Quality Evaluation of World Rices; IRRI: Manila, Philippines, 1993. [Google Scholar]
- Juliano, B. Rice Chemistry and Technology, 2nd ed.; American Association of Cereal Chemists, Incorporated: Saint Paul, MN, USA, 1985. [Google Scholar]
- Webb, B. Rice quality and grades. In Rice: Production and Utilization; Luh, B.S., Ed.; Avi Publishing Co., Inc.: Westport, CT, USA, 1980. [Google Scholar]
- Xia, D.; Zhou, H.; Qiu, L.; Jiang, H.; Zhang, Q.; Gao, G.; He, Y. Mapping and verification of grain shape QTLs based on an advanced backcross population in rice. PLoS ONE 2017, 12, e0187553. [Google Scholar] [CrossRef]
- Guo-Xin, Y.; Jin-Jie, L.; Zhang, Q.; Guang-Long, H.; Chao, C.; Bo, T.; Zhang, H.-L.; Zi-Chao, L. Mapping QTLs for Grain Weight and Shape Using Four Sister Near Isogenic Lines of Rice. Acta Agron. Sin. 2010, 36, 1310–1317. [Google Scholar]
- Xing, Y.; Tan, Y.-F.; Xu, C.-G.; Hua, J.-P.; Sun, X. Mapping quantitative trait loci for grain appearance traits of rice using a recombinant inbred line population. J. Integr. Plant Biol. 2001, 43, 721–726. [Google Scholar]
- Yoshida, S.; Ikegami, M.; Kuze, J.; Sawada, K.; Hashimoto, Z.; Ishii, T.; Nakamura, C.; Kamijima, O. QTL analysis for plant and grain characters of sake-brewing rice using a doubled haploid population. Breed. Sci. 2002, 52, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Rabiei, B.; Valizadeh, M.; Ghareyazie, B.; Moghaddam, M.; Ali, A. Identification of QTLs for rice grain size and shape of Iranian cultivars using SSR markers. Euphytica 2004, 137, 325–332. [Google Scholar] [CrossRef]
- Bazrkar-Khatibani, L.; Fakheri, B.-A.; Hosseini-Chaleshtori, M.; Mahender, A.; Mahdinejad, N.; Ali, J. Genetic mapping and validation of quantitative trait loci (QTL) for the grain appearance and quality traits in rice (Oryza sativa L.) by using recombinant inbred line (RIL) population. Int. J. Genom. 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhou, H.; Gu, Y.; Xia, D.; Wu, B.; Gao, G.; Zhang, Q.; He, Y. Mapping and verification of grain shape QTLs based on high-throughput SNP markers in rice. Mol. Breed. 2019, 39, 42. [Google Scholar] [CrossRef]
- Jianlong, X.; Qingzhong, X.; Lijun, L.; Zhikang, L. Genetic dissection of grain weight and its related traits in rice (Oryza sativa L.). Zhongguo Shuidao Kexue 2002, 16, 6–10. [Google Scholar]
- Yano, M.; Sasaki, T. Genetic and molecular dissection of quantitative traits in rice. In Oryza: From Molecule to Plant; Springer: New York, NY, USA, 1997; pp. 145–153. [Google Scholar]
- Yamamoto, T.; Yonemaru, J.; Yano, M. Towards the understanding of complex traits in rice: Substantially or superficially? DNA Res. 2009, 16, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Li, M.-M.; Lei, X.; Ren, J.-F.; Cao, G.-L.; Yu, L.-Q.; He, H.-H.; Han, L.-Z.; Koh, H.-j. Identification of quantitative trait loci for grain traits in japonica rice. Agric. Sci. China 2010, 9, 929–936. [Google Scholar] [CrossRef]
- Guangheng, Z.; Guoping, Z.; Qian, Q.; Luping, X.; Dali, Z.; Sheng, T.; Jingsong, B. QTL analysis of grain shape traits in different environments. Zhongguo Shuidao Kexue 2004, 18, 16–22. [Google Scholar]
- Hittalmani, S.; Shashidhar, H.; Bagali, P.G.; Huang, N.; Sidhu, J.; Singh, V.; Khush, G. Molecular mapping of quantitative trait loci for plant growth, yield and yield related traits across three diverse locations in a doubled haploid rice population. Euphytica 2002, 125, 207–214. [Google Scholar] [CrossRef]
- Yan, C.-J.; Liang, G.-H.; Chen, F.; Li, X.; Tang, S.-Z.; Yi, C.-D.; Tian, S.; Lu, J.-F.; Gu, M.-H. Mapping quantitative trait loci associated with rice grain shape based on an indica/japonica backcross population. Yi Chuan Xue Bao Acta Genet. Sinica 2003, 30, 711–716. [Google Scholar]
- Wacera, H.R.; Lee, H.-S.; Kim, K.-M. Biotechnology. Identification of Quantitative Trait Loci Associated with Grain Shape Using Cheongchenong/Nagdong Double Haploid Lines in Rice. Plant Breed. Biotechnol. 2016, 4, 188–197. [Google Scholar] [CrossRef]
- Yaobin, Q.; Peng, C.; Yichen, C.; Yue, F.; Derun, H.; Tingxu, H.; Xianjun, S.; Jiezheng, Y. QTL-Seq identified a major QTL for grain length and weight in rice using near isogenic F2 population. Rice Sci. 2018, 25, 121–131. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Huang, J.; Lan, H.; Wang, C.; Yin, C.; Wu, Y.; Tang, H.; Qian, Q.; Li, J.; et al. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice. Proc. Natl. Acad. Sci. USA 2012, 109, 21534–21539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.-J.; Huang, W.; Shi, M.; Zhu, M.-Z.; Lin, H.-X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet. 2007, 39, 623–630. [Google Scholar] [CrossRef]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef]
- Tanabe, S.; Ashikari, M.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Yano, M.; Yoshimura, A.; Kitano, H.; Matsuoka, M.; Fujisawa, Y.; et al. A Novel Cytochrome P450 Is Implicated in Brassinosteroid Biosynthesis via the Characterization of a Rice Dwarf Mutant, dwarf11, with Reduced Seed Length. Plant Cell 2005, 17, 776–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.; Wang, J.; Zhu, X.; Hao, W.; Wang, L.; Li, Q.; Zhang, L.; He, W.; Lu, B.; Lin, H.; et al. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat. Genet. 2008, 40, 1370–1374. [Google Scholar] [CrossRef]
- She, K.-C.; Kusano, H.; Koizumi, K.; Yamakawa, H.; Hakata, M.; Imamura, T.; Fukuda, M.; Naito, N.; Tsurumaki, Y.; Yaeshima, M.; et al. A Novel Factor FLOURY ENDOSPERM2 Is Involved in Regulation of Rice Grain Size and Starch Quality. Plant Cell 2010, 22, 3280–3294. [Google Scholar] [CrossRef] [Green Version]
- Shomura, A.; Izawa, T.; Ebana, K.; Ebitani, T.; Kajiya-Kanegae, H.; Konishi, S.; Yano, M. Deletion in a gene associated with grain size increased yields during rice domestication. Nat. Genet. 2008, 40, 1023–1028. [Google Scholar] [CrossRef]
- Li, Y.; Fan, C.; Xing, Y.; Jiang, Y.; Luo, L.; Sun, L.; Shao, D.; Xu, C.; Li, X.; Xiao, J.; et al. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet. 2011, 43, 1266–1269. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, K.; Kurinami, S.; Oki, K.; Abe, Y.; Ando, T.; Kono, I.; Yano, M.; Kitano, H.; Iwasaki, Y. A Novel Kinesin 13 Protein Regulating Rice Seed Length. Plant Cell Physiol. 2010, 51, 1315–1329. [Google Scholar] [CrossRef] [Green Version]
- Ashikari, M.; Wu, J.; Yano, M.; Sasaki, T.; Yoshimura, A. Rice gibberellin-insensitive dwarf mutant gene Dwarf 1 encodes the α-subunit of GTP-binding protein. Proc. Natl. Acad. Sci. USA 1999, 96, 10284–10289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, Y.; Mieda, K.; Ando, T.; Kono, I.; Yano, M.; Kitano, H.; Iwasaki, Y. The SMALL AND ROUND SEED1 (SRS1/DEP2) gene is involved in the regulation of seed size in rice. Genes Genet. Syst. 2010, 85, 327–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Li, S.; Liu, Q.; Wu, K.; Zhang, J.; Wang, S.; Wang, Y.; Chen, X.; Zhang, Y.; Gao, C.; et al. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat. Genet. 2015, 47, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, G.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.; Zeng, L.; Xu, E.; Xu, J.; et al. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef]
- Si, L.; Chen, J.; Huang, X.; Gong, H.; Luo, J.; Hou, Q.; Zhou, T.; Lu, T.; Zhu, J.; Shangguan, Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q.; et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef]
- Li, S.; Qian, Q.; Fu, Z.; Zeng, D.; Meng, X.; Kyozuka, J.; Maekawa, M.; Zhu, X.; Zhang, J.; Li, J.; et al. Short panicle1encodes a putative PTR family transporter and determines rice panicle size. Plant J. 2009, 58, 592–605. [Google Scholar] [CrossRef]
- Segami, S.; Kono, I.; Ando, T.; Yano, M.; Kitano, H.; Miura, K.; Iwasaki, Y. Small and round seed 5 gene encodes alpha-tubulin regulating seed cell elongation in rice. Rice 2012, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Yang, C.-D.; Fan, Y.; Zhuang, J.; Li, X. Genetic dissection of a thousand-grain weight quantitative trait locus on rice chromosome. Sci. Bull. 2008, 53, 2326–2332. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Singh, A.K.; Sharma, T.R.; Singh, A.; Singh, N.K. Fine mapping of grain length QTLs on chromosomes 1 and 7 in Basmati rice (Oryza sativa L.). J. Plant Biochem. Biotechnol. 2011, 21, 157–166. [Google Scholar] [CrossRef]
- Zhang, H.-W.; Fan, Y.-Y.; Zhu, Y.-J.; Chen, J.-Y.; Yu, S.; Zhuang, J. Dissection of the qTGW1.1 region into two tightly-linked minor QTLs having stable effects for grain weight in rice. BMC Genet. 2016, 17, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Sun, P.; He, Q.; Shu, F.; Wang, J.; Deng, H. Fine mapping of GS2, a dominant gene for big grain rice. Crop. J. 2013, 1, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.Y.; Wan, J.M.; Jiang, L.; Wang, J.K.; Zhai, H.Q.; Weng, J.F.; Wang, H.L.; Lei, C.L.; Zhang, X.; Cheng, Z.J.; et al. QTL analysis for rice grain length and fine mapping of an identified QTL with stable and major effects. Theor. Appl. Genet. 2006, 112, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Thomson, M.; McCouch, S.R. Fine Mapping of a Grain-Weight Quantitative Trait Locus in the Pericentromeric Region of Rice Chromosome. Genetics 2004, 168, 2187–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Ma, L.; Jiang, H.; Zeng, D.; Hu, J.; Wu, L.; Gao, Z.; Zhang, G.; Qian, Q. Genetic Analysis and Fine Mapping of Two Genes for Grain Shape and Weight in Rice. J. Integr. Plant Biol. 2009, 51, 45–51. [Google Scholar] [CrossRef]
- Liu, T.; Shao, D.; Kovi, M.R.; Xing, Y. Mapping and validation of quantitative trait loci for spikelets per panicle and 1,000-grain weight in rice (Oryza sativa L.). Theor. Appl. Genet. 2009, 120, 933–942. [Google Scholar] [CrossRef]
- Zhang, Q.; Yao, G.-X.; Hu, G.-L.; Chen, C.; Tang, B.; Zhang, H.; Li, Z.-C. Fine Mapping of qTGW3-1, a QTL for 1000-Grain Weight on Chromosome 3 in Rice. J. Integr. Agric. 2012, 11, 879–887. [Google Scholar] [CrossRef]
- Tang, S.-Q.; Shao, G.-N.; Wei, X.; Chen, M.-L.; Sheng, Z.-H.; Luo, J.; Jiao, G.-A.; Xie, L.-H.; Hu, P. QTL mapping of grain weight in rice and the validation of the QTL qTGW3. Gene 2013, 527, 201–206. [Google Scholar] [CrossRef]
- Kato, T.; Segami, S.; Toriyama, M.; Kono, I.; Ando, T.; Yano, M.; Kitano, H.; Miura, K.; Iwasaki, Y. Detection of QTLs for grain length from large grain rice (Oryza sativa L.). Breed. Sci. 2011, 61, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.-X.; Zhu, M.-Z.; Shi, M.; Gao, J.-P.; Lin, H.-X. Fine mapping and candidate gene analysis of spd6, responsible for small panicle and dwarfness in wild rice (Oryza rufipogon Griff.). Theor. Appl. Genet. 2009, 119, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Luo, L.; Yan, W.; Kovi, M.R.; Zhan, W.; Xing, Y. Genetic dissection of rice grain shape using a recombinant inbred line population derived from two contrasting parents and fine mapping a pleiotropic quantitative trait locus qGL7. BMC Genet. 2010, 11, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, G.; Wei, X.; Chen, M.; Tang, S.; Luo, J.; Jiao, G.; Xie, L.; Hu, P. Allelic variation for a candidate gene for GS7, responsible for grain shape in rice. Theor. Appl. Genet. 2012, 125, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Gong, R.; Tan, Y.; Yu, S. Mapping and characterization of the major quantitative trait locus qSS7 associated with increased length and decreased width of rice seeds. Theor. Appl. Genet. 2012, 125, 1717–1726. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Jin, F.; Song, M.-H.; Suh, J.-P.; Hwang, H.-G.; Kim, Y.-G.; McCouch, S.R.; Ahn, S.-N. Fine mapping of a yield-enhancing QTL cluster associated with transgressive variation in an Oryza sativa × O. rufipogon cross. Theor. Appl. Genet. 2007, 116, 613–622. [Google Scholar] [CrossRef]
- Onishi, K.; Horiuchi, Y.; Ishigoh-Oka, N.; Takagi, K.; Ichikawa, N.; Maruoka, M.; Sano, Y. A QTL Cluster for Plant Architecture and Its Ecological Significance in Asian Wild Rice. Breed. Sci. 2007, 57, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef]
- Zhu, Q.; Ge, S. Phylogenetic relationships among A-genome species of the genus Oryza revealed by intron sequences of four nuclear genes. New Phytol. 2005, 167, 249–265. [Google Scholar] [CrossRef]
- Kato, S. On the affinity of rice varieties as shown by fertility of hybrid plants. Bull. Sci. Fac. Agric. Kyushu Univ. 1928, 3, 132–147. [Google Scholar]
- Lee, S.-M.; Kang, J.-W.; Lee, J.-Y.; Seo, J.; Shin, D.; Cho, J.-H.; Jo, S.; Song, Y.-C.; Park, D.-S.; Ko, J.-M.; et al. QTL Analysis for Fe and Zn Concentrations in Rice Grains Using a Doubled Haploid Population Derived from a Cross between Rice (Oryza sativa) Cultivar 93-11 and Milyang. Plant Breed. Biotechnol. 2020, 8, 69–76. [Google Scholar] [CrossRef]
- Kabange, N.R.; Park, S.-Y.; Shin, D.; Lee, S.-M.; Jo, S.-M.; Kwon, Y.; Cha, J.-K.; Song, Y.-C.; Ko, J.-M.; Lee, J.-H. Identification of a Novel QTL for Chlorate Resistance in Rice (Oryza sativa L.). Agriculture 2020, 10, 360. [Google Scholar] [CrossRef]
- Thorpe, R.S. Multiple group principal component analysis and population differentiation. J. Zool. 1988, 216, 37–40. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Keb-Llanes, M.; Gonzalez, G.; Chi-Manzanero, B.; Infante, D. A rapid and simple method for small-scale DNA extraction inAgavaceae and other tropical plants. Plant Mol. Boil. Rep. 2002, 20, 299. [Google Scholar] [CrossRef]
- Cheon, K.-S.; Baek, J.; Cho, Y.-I.; Jeong, Y.-M.; Lee, Y.-Y.; Oh, J.; Won, Y.J.; Kang, D.-Y.; Oh, H.; Kim, S.L.; et al. Single Nucleotide Polymorphism (SNP) Discovery and Kompetitive Allele-Specific PCR (KASP) Marker Development with Korean Japonica Rice Varieties. Plant Breed. Biotechnol. 2018, 6, 391–403. [Google Scholar] [CrossRef]
- Seo, J.; Lee, G.; Jin, Z.; Kim, B.; Chin, J.H.; Koh, H.-J. Development and application of indica–japonica SNP assays using the Fluidigm platform for rice genetic analysis and molecular breeding. Mol. Breed. 2020, 40, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop. J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Nietzsche, M.; Schießl, I.; Börnke, F. The complex becomes more complex: Protein-protein interactions of SnRK1 with DUF581 family proteins provide a framework for cell-and stimulus type-specific SnRK1 signaling in plants. Front. Plant Sci. 2014, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Seung, D.; Schreier, T.B.; Bürgy, L.; Eicke, S.; Zeeman, S.C. Two Plastidial Coiled-Coil Proteins Are Essential for Normal Starch Granule Initiation in Arabidopsis. Plant Cell 2018, 30, 1523–1542. [Google Scholar] [CrossRef] [Green Version]
- Tsai, A.Y.-L.; Gazzarrini, S. Trehalose-6-phosphate and SnRK1 kinases in plant development and signaling: The emerging picture. Front. Plant Sci. 2014, 5, 119. [Google Scholar] [CrossRef] [Green Version]
- Krasnoperova, E.E.; Buy, D.D.; Goriunova, I.I.; Isayenkov, S.; Karpov, P.A.; Blume, Y.B.; Yemets, A.I. The Potential Role of SnRK1 Protein Kinases in the Regulation of Cell Division in Arabidopsis thaliana. Cytol. Genet. 2019, 53, 185–191. [Google Scholar] [CrossRef]
- Xie, X.; Song, M.-H.; Jin, F.; Ahn, S.-N.; Suh, J.-P.; Hwang, H.-G.; McCouch, S.R. Fine mapping of a grain weight quantitative trait locus on rice chromosome 8 using near-isogenic lines derived from a cross between Oryza sativa and Oryza rufipogon. Theor. Appl. Genet. 2006, 113, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-M.; Balkunde, S.; Yang, P.; Yoon, D.-B.; Ahn, S.-N. Fine mapping of grain weight QTL, tgw11 using near isogenic lines from a cross between Oryza sativa and O. grandiglumis. Genes Genom. 2011, 33, 259–265. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits (a) | QTL (b) | Chr (c) | Position (cM) (d) | Left Marker (e) | Right Marker (f) | LOD (g) | PVE (%) (h) | Add (i) |
|---|---|---|---|---|---|---|---|---|
| GLE | qGLE-1-1 | 1 | 32 | Os01_KJ01_127 | Os01_KJ01_129 | 4.8451 | 10.808 | −0.087 |
| qGLE-3-1 | 3 | 46 | Os03_id3003462 | Os03_KJ03_017 | 8.0562 | 19.294 | 0.116 | |
| qGLE-3-2 | 3 | 203 | Os03_cmb0336_5 | Os03_id3018439 | 3.1436 | 13.146 | −0.058 | |
| qGLE-5-1 | 5 | 24 | Os05_KJ05_017 | Os05_KJ05_013 | 3.9426 | 15.579 | −0.064 | |
| qGLE-7-1 | 7 | 156 | Os07_id7003072 | Os07_cmb0723_0 | 5.6943 | 13.751 | 0.1019 | |
| qGLE-12-1 | 12 | 13 | Os12_id12000076 | Os12_cmb1202_4 | 4.0984 | 9.1872 | 0.0801 | |
| qGLE-12-2 | 12 | 130 | Os12_cmb1224_0 | Os12_cmb1226_0 | 3.8695 | 8.4563 | −0.077 | |
| GLL | qGLL-1-1 | 1 | 19 | Os01_KJ01_125 | Os01_KJ01_127 | 5.3501 | 9.0539 | −0.107 |
| qGLL-1-2 | 1 | 81 | Os01_KJ01_097 | Os01_ad01015967 | 4.5891 | 6.2337 | 0.091 | |
| qGLL-3-1 | 3 | 46 | Os03_id3003462 | Os03_KJ03_017 | 10.676 | 16.413 | 0.136 | |
| qGLL-7-1 | 7 | 150 | Os07_id7003072 | Os07_cmb0723_0 | 18.017 | 32.151 | 0.1963 | |
| qGLL-11-1 | 11 | 7 | Os11_id11000131 | Os11_KJ11_005 | 3.6033 | 5.1524 | 0.0766 | |
| GTE | qGTE-3-1 | 3 | 25 | Os03_ad03000001 | Os03_KJ03_007 | 6.9497 | 10.081 | 0.032 |
| qGTE-3-2 | 3 | 88 | Os03_KJ03_031 | Os03_ah03001094 | 9.4914 | 12.644 | −0.036 | |
| qGTE-5-1 | 5 | 24 | Os05_KJ05_017 | Os05_KJ05_013 | 14.188 | 20.25 | −0.046 | |
| qGTE-12-1 | 12 | 14 | Os12_cmb1202_4 | Os12_KJ12_007 | 6.0808 | 7.5265 | 0.0277 | |
| GTL | qGTL-1-1 | 1 | 150 | Os01_id1009557 | Os01_KJ01_005 | 7.2095 | 5.5653 | −0.025 |
| qGTL-3-1 | 3 | 27 | Os03_ad03000001 | Os03_KJ03_007 | 9.9909 | 9.027 | 0.0291 | |
| qGTL-3-2 | 3 | 88 | Os03_KJ03_031 | Os03_ah03001094 | 13.934 | 12.367 | −0.034 | |
| qGTL-5-1 | 5 | 24 | Os05_KJ05_017 | Os05_KJ05_013 | 21.996 | 23.347 | −0.047 | |
| qGTL-7-1 | 7 | 210 | Os07_cmb0730_3 | Os07_KJ07_085 | 4.9114 | 3.7314 | −0.019 | |
| qGTL-10-1 | 10 | 85 | Os10_KJ10_039 | Os10_ah10001182 | 10.159 | 8.4954 | −0.029 | |
| qGTL-12-1 | 12 | 13 | Os12_id12000076 | Os12_cmb1202_4 | 14.613 | 13.242 | 0.0353 | |
| GWL | qGWL-1-1 | 1 | 127 | Os01_KJ01_075 | Os01_SaF_CT | 4.2995 | 6.2645 | 0.0568 |
| qGWL-2-1 | 2 | 124 | Os02_KJ02_043 | Os02_KJ02_045 | 5.5625 | 8.0429 | −0.05 | |
| qGWL-4-1 | 4 | 30 | Os04_KJ04_049 | Os04_cmb0418_8 | 3.8361 | 5.7157 | −0.042 | |
| qGWL-5-1 | 5 | 24 | Os05_KJ05_017 | Os05_KJ05_013 | 17.213 | 31.664 | −0.098 | |
| qGWL-8-4 | 8 | 100 | Os08_cmb0824_7 | Os08_GW8--AG | 3.0747 | 4.332 | 0.0377 | |
| qGWL-10-1 | 10 | 85 | Os10_KJ10_039 | Os10_ah10001182 | 9.597 | 15.122 | −0.07 | |
| LWRE | qLWRE-5-1 | 5 | 24 | Os05_KJ05_017 | Os05_KJ05_013 | 9.8132 | 18.992 | −0.894 |
| qLWRE-7-1 | 7 | 93 | Os07_KJ07_021 | Os07_id7001155 | 6.1326 | 15.95 | 0.0883 | |
| qLWRE-7-2 | 7 | 209 | Os07_id7006027 | Os07_cmb0730_3 | 4.8302 | 12.15 | 0.0797 | |
| LWRL | qLWRL-1-1 | 1 | 80 | Os01_KJ01_097 | Os01_ad01015967 | 4.8679 | 2.7977 | 0.0496 |
| qLWRL-3-1 | 3 | 156 | Os03_ad03014175 | Os03_Hd6-1AT | 9.3938 | 6.6007 | 0.0717 | |
| qLWRL-4-1 | 4 | 159 | Os04_ah04001252 | Os04_id4009823 | 5.6156 | 3.272 | −0.053 | |
| qLWRL-5-1 | 5 | 24 | Os05_KJ05_017 | Os05_KJ05_013 | 25.747 | 22.935 | 0.1315 | |
| qLWRL-7-1 | 7 | 67 | Os07_cmb0700_1 | Os07_ud7000187 | 34.456 | 37.665 | 0.1677 | |
| qLWRL-10-1 | 10 | 68 | Os10_cmb1016_4 | Os10_wd10003790 | 3.3679 | 2.0534 | 0.0422 | |
| TGWE | qTGWE-2-1 | 2 | 132 | Os02_KJ02_057 | Os02_id2012773 | 3.7903 | 6.4953 | −0.525 |
| qTGWE-3-1 | 3 | 193 | Os03_ah03002520 | Os03_cmb0336_5 | 3.0986 | 5.6929 | −0.488 | |
| qTGWE-5-1 | 5 | 4 | Os05_KJ05_029 | Os05_cmb0511_1 | 8.5874 | 23.405 | 0.1099 | |
| qTGWE-8-1 | 8 | 102 | Os08_GW8--AG | Os08_id8007764 | 4.8611 | 8.5632 | 0.6181 | |
| qTGWE-11-1 | 11 | 110 | Os11_id11008929 | Os11_cmb1127_4 | 3.5075 | 6.0367 | −0.526 | |
| qTGWE-12-1 | 12 | 14 | Os12_cmb1202_4 | Os12_KJ12_007 | 7.2136 | 13.525 | 0.7519 | |
| TGWL | qTGWL-2-1 | 2 | 138 | Os02_KJ02_057 | Os02_id2012773 | 6.0649 | 12.667 | −0.886 |
| qTGWL-3-1 | 3 | 31 | Os03_KJ03_007 | Os03_ah03000403 | 7.4325 | 14.514 | 0.9248 | |
| qTGWL-8-1 | 8 | 102 | Os08_GW8--AG | Os08_id8007764 | 3.4418 | 6.7419 | 0.6509 | |
| qTGWL-12-1 | 12 | 52 | Os12_id12002113 | Os12_id12003700 | 6.4314 | 13.021 | 0.9115 |
| No. | Locus Name | Gene Name | Description | Similar Papers |
|---|---|---|---|---|
| 1 | Os05g08800 | DUF581 | DUF581 domain containing protein, expressed | [93] |
| 2 | Os05g08790 | MFP1 | MAR-binding filament-like protein 1, putative, expressed | [94] |
| 3 | Os05g08810 | PPL3-K | phosphatidylinositol 3-kinase, root isoform, putative, expressed | - |
| 4 | Os05g08840 | DnaK | DnaK family protein, putative, expressed | - |
| 5 | Os05g08850 | CP450 | cytochrome P450, putative, expressed | [51] |
| 6 | Os05g08870 | Ty1-copia | retrotransposon protein, putative, Ty1-copia subclass, expressed | - |
| 7 | Os05g08920 | LEA | Late embryogenesis abundant protein, putative, expressed, LEA, TF | - |
| 8 | Os05g08930 | Clo-lumen | chloroplast lumen common family protein, putative, expressed | - |
| 9 | Os05g08940 | Mariner | Transposon protein, putative, Mariner sub-class, expressed | - |
| 10 | Os05g08960 | Ubiquitin E2 | Ubiquitin-conjugating enzyme, E2 putative, expressed | [49] |
| 11 | Os05g08970 | SSRP1 | SSRP1-like FACT complex subunit, putative, expressed, TF | - |
| 12 | Os05g09020 | WRKY67 | WRKY67 Transcription factor, expressed | - |
| 13 | Os05g09050 | MLO | MLO domain containing protein, putative, expressed | - |
| 14 | Os05g09080 | Ty3-gyspsy | Retrotransposon protein, putative, Ty3-gypsy subclass, expressed | - |
| 15 | Os05g10210 | HAD | HAD superfamily phosphatase, putative, expressed | - |
| 16 | Os05g10230 | En-Spm | Transposon protein, putative, CACTA, En/Spm sub-class, expressed | - |
| 17 | Os05g10380 | RWP-RK | RWP-RK, putative, expressed | - |
| 18 | Os05g10580 | Cullin | Cullin family domain containing protein, putative, expressed | - |
| 19 | Os05g10620 | NAM | no apical meristem protein, putative, expressed | - |
| 20 | Os05g10650 | PFK6 | 6-phosphofructokinase, putative, expressed | - |
| 21 | Os05g10670 | Zinc finger | Zinc finger CCCH type family protein, putative, expressed | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.-W.; Kabange, N.R.; Phyo, Z.; Park, S.-Y.; Lee, S.-M.; Lee, J.-Y.; Shin, D.; Cho, J.H.; Park, D.-S.; Ko, J.-M.; et al. Combined Linkage Mapping and Genome-Wide Association Study Identified QTLs Associated with Grain Shape and Weight in Rice (Oryza sativa L.). Agronomy 2020, 10, 1532. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101532
Kang J-W, Kabange NR, Phyo Z, Park S-Y, Lee S-M, Lee J-Y, Shin D, Cho JH, Park D-S, Ko J-M, et al. Combined Linkage Mapping and Genome-Wide Association Study Identified QTLs Associated with Grain Shape and Weight in Rice (Oryza sativa L.). Agronomy. 2020; 10(10):1532. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101532
Chicago/Turabian StyleKang, Ju-Won, Nkulu Rolly Kabange, Zarchi Phyo, So-Yeon Park, So-Myeong Lee, Ji-Yun Lee, Dongjin Shin, Jun Hyeon Cho, Dong-Soo Park, Jong-Min Ko, and et al. 2020. "Combined Linkage Mapping and Genome-Wide Association Study Identified QTLs Associated with Grain Shape and Weight in Rice (Oryza sativa L.)" Agronomy 10, no. 10: 1532. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101532