1. Introduction
Cassava (
Manihot esculenta (L.) Crantz) is an important source of food in the tropics; its starchy roots are consumed fresh (for low toxic varieties) and as a processed starch. Cassava is also used for animal feed [
1]. Cassava is grown in tropical and sub-tropical areas (30° N to 30° S) including Brazil, South Africa and Southeast Asia countries [
1]. This crop is consumed by more than a billion people in 105 countries [
2]. Asia (29.4%) is the second largest cassava production area in the world after Africa (60.9%) [
3]. Under optimum conditions, cassava yield in Asia can reach 90 t ha
−1 [
1]. Yet, cassava average yields in Asia (23.6 t ha
−1) [
4] are usually less than the world average yield for many reasons, including biotic stress—such as insect and disease—and abiotic stress—such as lower soil fertility, drought, water flooding and extreme temperatures [
5,
6,
7].
Although cassava has a long crop cycle, ranging from 12 to 24 months, the planting date will determine the climatic conditions the crop will experience during critical growth stages including canopy establishment, root initiation and storage root accumulation [
8]. In Asia, the early growth stages of cassava planted during the early rainy season will be subjected to higher rainfall, temperature, RH and SR than those planted in the late rainy season. However, drought will often hit early rainy season plantings when they are in the later growth stages, reducing canopy size (CS) and yield [
9,
10,
11].
The size of the plant canopy is important for light interception and photosynthesis and plays a major role in photosynthesis capacity. Canopy size determined light interception and photosynthesis capacity [
12,
13]. Different varieties have different canopy architectures, which change light availability to the lower leaves in the canopy [
14,
15]. Cassava leaf photosynthesis, total biomass (BM) and root yield are known to be positively and significantly correlated with each other [
16,
17]. In addition, plant height is often found to have strong and positive correlations with leaf area, fresh root and dry matter yield [
18], as has leaf area index (LAI) [
1,
19,
20,
21,
22,
23]. Yet the highest yielding genotypes of cassava do not always have the highest LAI [
24], as leaf arrangement and appropriate leaf size may help increase photosynthetic efficiency and, thus, compensate for lower leaf area [
25]. Good canopy architecture, therefore, may help increase photosynthetic rates in the lower leaves of the canopy and increase cassava yield.
Cock et al. [
20] suggested that high yielding cassava should have late forking and branching and possess large leaves and long leaf life. However, the effects of the canopy components on light penetration in cassava genotypes have not been clearly defined because cassava has a long-life cycle. Throughout the life cycle of cassava, the crop encounters seasonal variations in climatic factors such as light intensity, temperature and humidity. These factors affect the change in canopy of cassava and cassava genotypes respond differently to environmental changes. While the responses of CS on seasonal variation under different water regimes had been reported, they generally use only one genotype [
11]. Therefore, the objective of this study was to determine canopy architecture, light penetration and photosynthesis of three cassava genotypes under different seasonal variations. The information obtained in this study will be useful for selection of cassava genotypes with good plant architecture, which will lead to improved photosynthesis and root yield.
4. Discussion
In this study, the research was focused on examining the effects of climatic factors on CS and yield of cassava genotypes. Planting dates (31.0–51.5%) contributed to a larger portion of variation in BM, storage root DW and SRY than other sources of variation. Similar results were reported by Mahakosee et al. [
11], who observed that planting date contributed the largest portion of variation in BM, storage root DW and storage root fresh weight (46.1–60.9%) compared to the effects of year and water regime. In this study, the interaction between genotype and planting date showed a much higher magnitude than genotype by year. This indicates that test of cassava genotypes for growth and yield should be evaluated under multiple seasons instead of years.
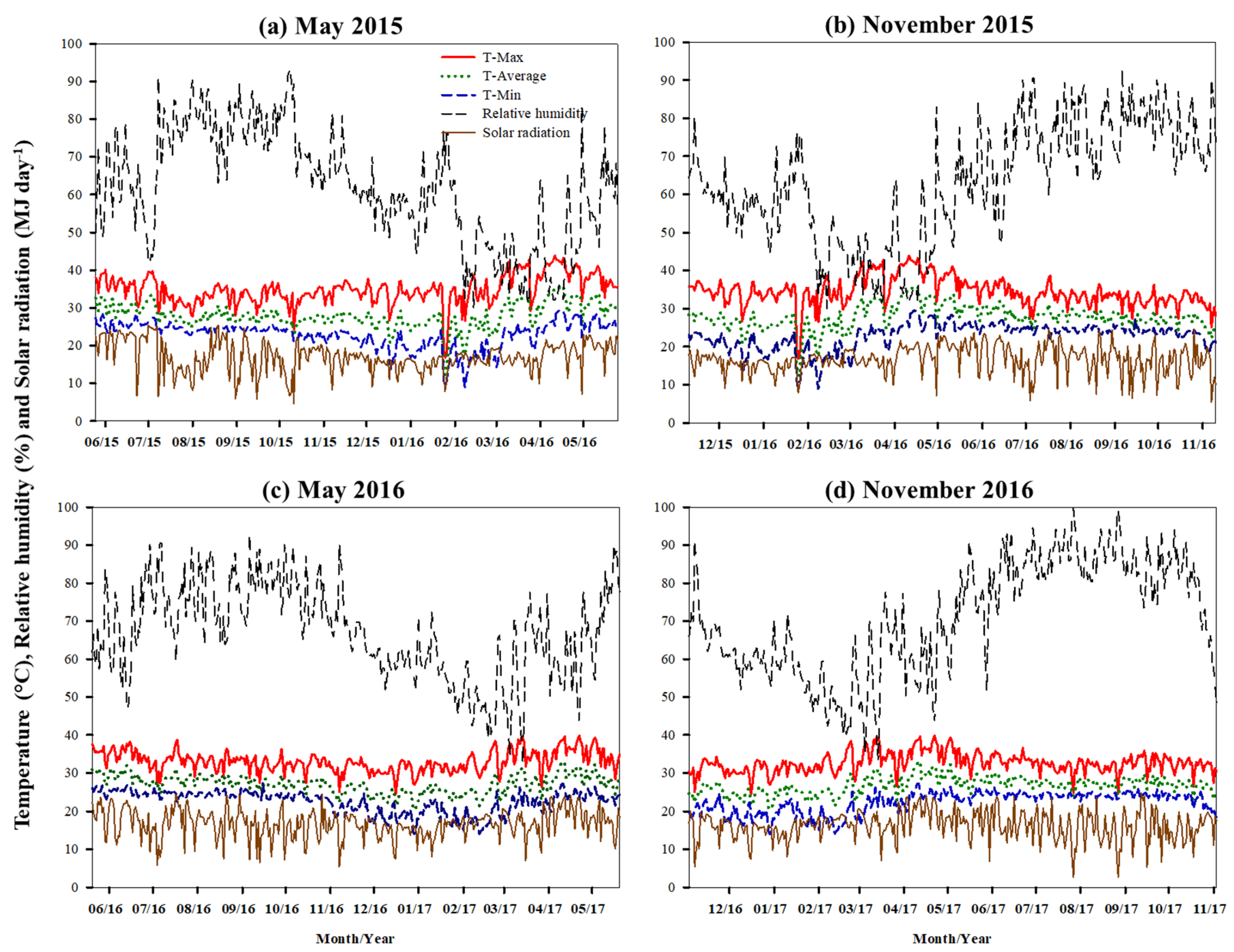
Differences in climatic factors during the growing seasons, including RH, maximum and minimum temperatures, SR and DL impacted CS. When planted in May in the early rainy season with high RH (63.6–79.0%), temperature (32.6–38.2 °C), SR (5.8–25.4 MJ m−2 day−1, average 17.6 MJ m−2 day−1) and long DL (12.08–13.10 h), CS rapidly increased in the early growth stages (1–4 MAP). Low temperature (16.3–22.6 °C), RH (40.0–61.1%), SR (4.6–24.5 MJ m−2 day−1, average 15.4 MJ m−2 day−1) and short DL (11.18–11.97 h) decreased CS in the mid and late growth stages for all cassava genotypes. However, at the mid and late growth stages, Rayong 11 maintained higher CS than KU50 and CMR38–125-77. For November plantings, the cassava genotypes were subjected to low temperatures (17.7–21.8 °C), low RH (42.9–68.1%), SR (5.4–27.6 MJ m−2 day−1, average 15.8 MJ m−2 day−1) and short DL (11.16–11.74 h), in the early growth stage (1–4 MAP), and therefore, canopy development was slower than the May plantings. These findings were supported by stepwise regression analysis. The analysis with high coefficient of determination (R2) indicated that climatic factors explained a large part of the variation of CS in the three cassava genotypes. In May 2015, SR and RH both strongly contributed to CH and LAI for all three cassava genotypes, while SR and minimum temperature were important factors for CW. However, RH and DL explained a large part of the variation for CS of the three genotypes planted in November 2015. For the crop planted in May 2016, RH and DL were identified as the main factors affecting CS of KU50. SR and DL were important factors for CS of Rayong 11. However, SR, DL, maximum temperature and minimum temperature described CS of CMR38-125-77. For the November 2016 planting date, SR, RH and DL were the main factors determining CS of Rayong 11 and CMR38-125-77, whereas the CS of KU50 could be explained by minimum and maximum temperature, RH and DL.
The effect of temperature on growth and development of cassava such as root initiation, leaf formation, leaf size and storage root formation had been reported [
7]. Cassava can be grown under 16–38 °C [
8,
20] and the most favorable temperature has been reported to be between 25–29 °C by Alves [
8]. In this study, the crops planted in May of both years were subjected to minimum temperatures > 25 °C from planting to 5 MAP, but in the crops planted in November, the minimum temperatures were lower than 25 °C from planting until 4 MAP. Therefore, early season canopy growth of crops planted in May was larger than the planting in November. In contrast, the crops planted in November had a more optimum temperatures (>25 °C) during mid and late growth stages. The crops planted in November also maintained CS until harvest; however, the KU50 and CMR38-125-77 maintained higher CS than Rayong 11. These results were supported by Lahai et al. [
24], who observed that cassava genotypes with better maintenance of canopy parameters during a mid-season drought could produce higher SRY. Lenis et al. [
35] also reported that cassava genotypes with leaf retention traits gave higher BM and yield than the genotypes without those traits; additionally, Lenis et al. [
35] mentioned that cassava yields were positively correlated with leaf longevity, leaf retention and LAI. However, leaf longevity, leaf retention and LAI of cassava depends on genotypes, shade level and environmental factors such as temperature. Irikura et al. [
36] found that cassava grown under cool conditions (20 °C) had longer leaf life than warm conditions (24 °C and 28 °C). Phosaengsri et al. [
37] found that KU50 had the longest leaf life and leaf area duration during 3–6 and 6–9 MAP in all dates studied, and CMR38-125-77 had the highest LAI at 3, 6 and 9 MAP; therefore, both of KU50 and CMR38-125-77 gave a higher biomass and yield in all planting dates. However, the optimum LAI for storage root bulking in cassava was 3–3.5 [
20]. In this study, the crop planted in November maintained LAI greater than 3 at the storage root bulking stage (6–9 MAP), whereas the crop planted in May had an LAI lower than 3. The lower LAI for the crop planted in May was caused mainly by low temperature and low RH at mid and late growth stages, resulting in falling of old leaves and delaying the formation of new leaves. However, Rayong 11 in the May planting date seemed to maintain higher LAI and lower light transmission than CMR38-125-77 and KU50 during 6–9 MAP. Lower light availability in the lower leaves in the canopy resulted in low leaf photosynthesis in the lower leaves in the canopy of Rayong 11.
Increasing SR may also increase the net photosynthesis and is correlated to yield of cassava [
1,
38,
39]. DL, along with increasing the photosynthetically active period, is known to affect assimilates partitioning, flowering and tuberization [
8]. In this study, crops planted in May of both years were subjected to long days in early growth, while a November planting date resulted in short days in early growth stages. The effect of the photoperiod on storage root initiation in early growth stages was reported by Lowe et al. [
40]. Short days (8 h) induced storage root initiation earlier than long days (14 and 20 h), and storage root dry weight of shorts day was greater than long days. A study by Veltkamp [
41] indicated that long days induce shoot growth and also increase the amount of forking with the first fork occurring earlier, while short days increase storage root growth and reduce shoot growth, without influencing total BM. A current report pointed out that low temperature and low SR reduced BM accumulation rates in both early growth and late growth stages of cassava, whereas high temperature and high SR often resulted in higher BM accumulation rates in both early and late growth stages [
29].
In this paper’s study most, genotypes planted in November had higher BM and storage root DW than the crops planted in May, except for Rayong 11. In May plantings, BM and yield were not significant different among cassava genotypes. Rayong 11 had the lowest BM and yield in November plantings for both years. Rayong 11 seemed to have smaller CS than the others, although it was an early forking type [
42]. The forking of cassava affects the canopy development and increases LAI and yield [
1,
36]. Forking in cassava produced more apex and more leaves than non-forking types [
36,
43]. However, the new leaves in forking branches are often smaller than the main stem leaves [
36]. More leaf formation, short petioles and short internode length produced a dense canopy. In this study, Rayong 11 had higher LAI, lower light penetration and lower leaf photosynthesis in all leaf layers, resulting in lower yields. According to Lahai et al. [
24], TMS 4 (2)1425 forks early, about 2 MAP and continued branching until harvest. Due to this branching habit, it produced the highest number of leaves compared to other cultivars. Our report found that Rayong 11 produced higher shoot DW, but it had lower storage root DW. This might be due to competition between shoots and storage roots for photo assimilate in cassava [
44].
Although KU50 and CMR38-125-77 were forking type genotypes similar to Rayong 11, the KU50 and CMR38-125-77 had higher storage root DW and yield than Rayong 11 in the November plantings. In this study, the first forking of KU50 and CMR38-125-77 were higher than Rayong 11. Moreover, the KU50 and CMR38-125-77 had a wider angle of forking than Rayong 11. It was due to greater light penetration in canopy of KU50 and CMR38-125-77. However, the variation of forking in cassava was depended on genotypes and strongly correlated with climatic factors. Irikura et al. [
35] reported that increasing temperature from 20 to 24 °C resulted in early branching and increased leaf formation and size. Phoncharean et al. [
41] reported that Rayong 11 had earlier first forking (57 days after planting) than KU50 and CMR38-125-77. SR, minimum temperature and DL were the main factors inducing first forking of Rayong 11 and CMR38-125-77. The results of our study supported Cock et al. [
20], who suggested that high yielding cassava genotypes should have late forking and branching and possess large leaves with long leaf life. In conclusion, KU50 and CMR38-125-77 had a better canopy architecture and could maintain CS under cool temperatures and low RH conditions. Moreover, both KU50 and CMR38-125-77 had higher leaf photosynthetic capacity and better leaf spatial arrangement for a higher light interception at mid and late growth stages.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}