Simulation Models on the Ecology and Management of Arable Weeds: Structure, Quantitative Insights, and Applications

 , ,
, ,  , , , , and
, , , , and
Abstract
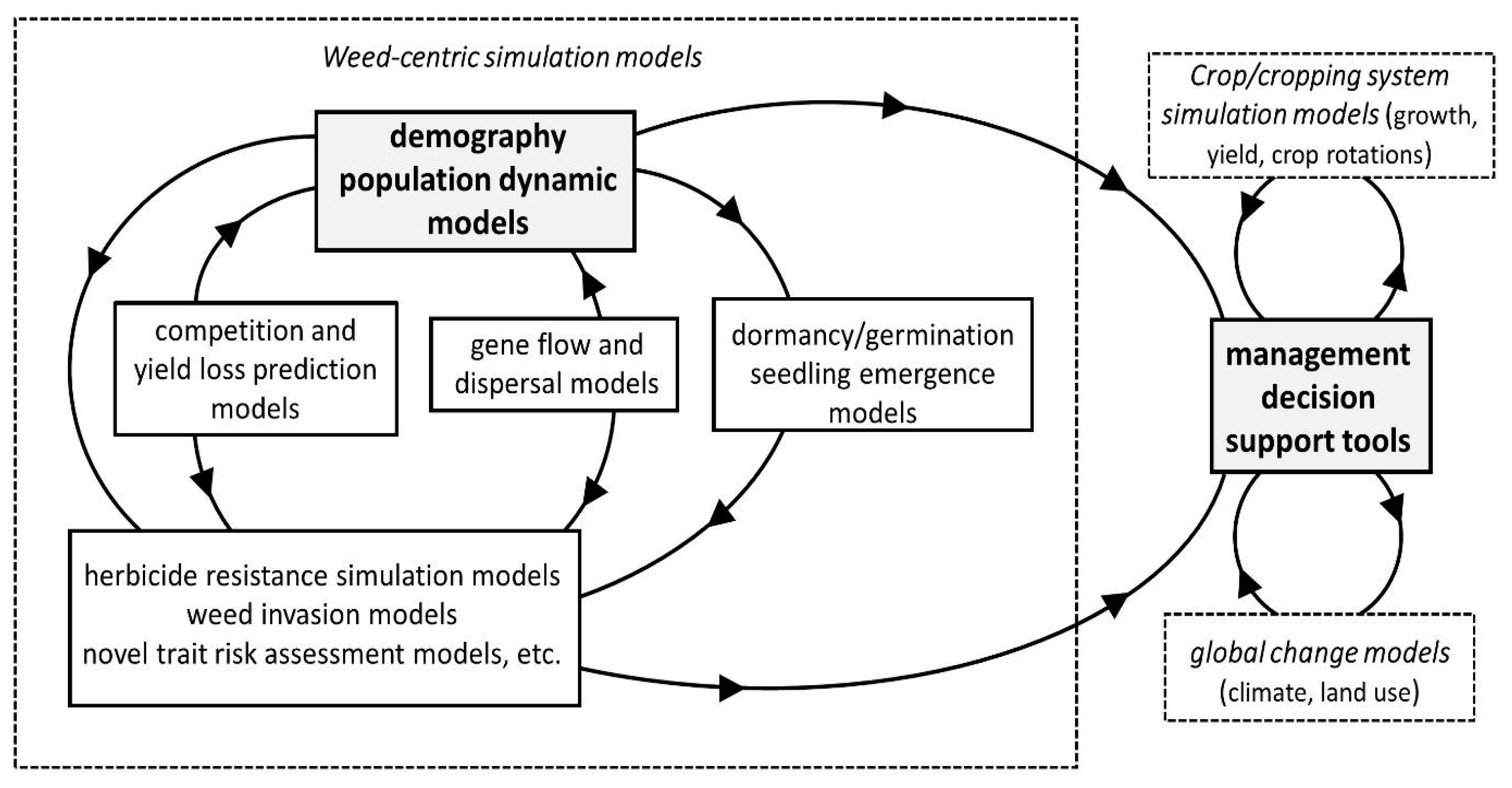
:1. Introduction
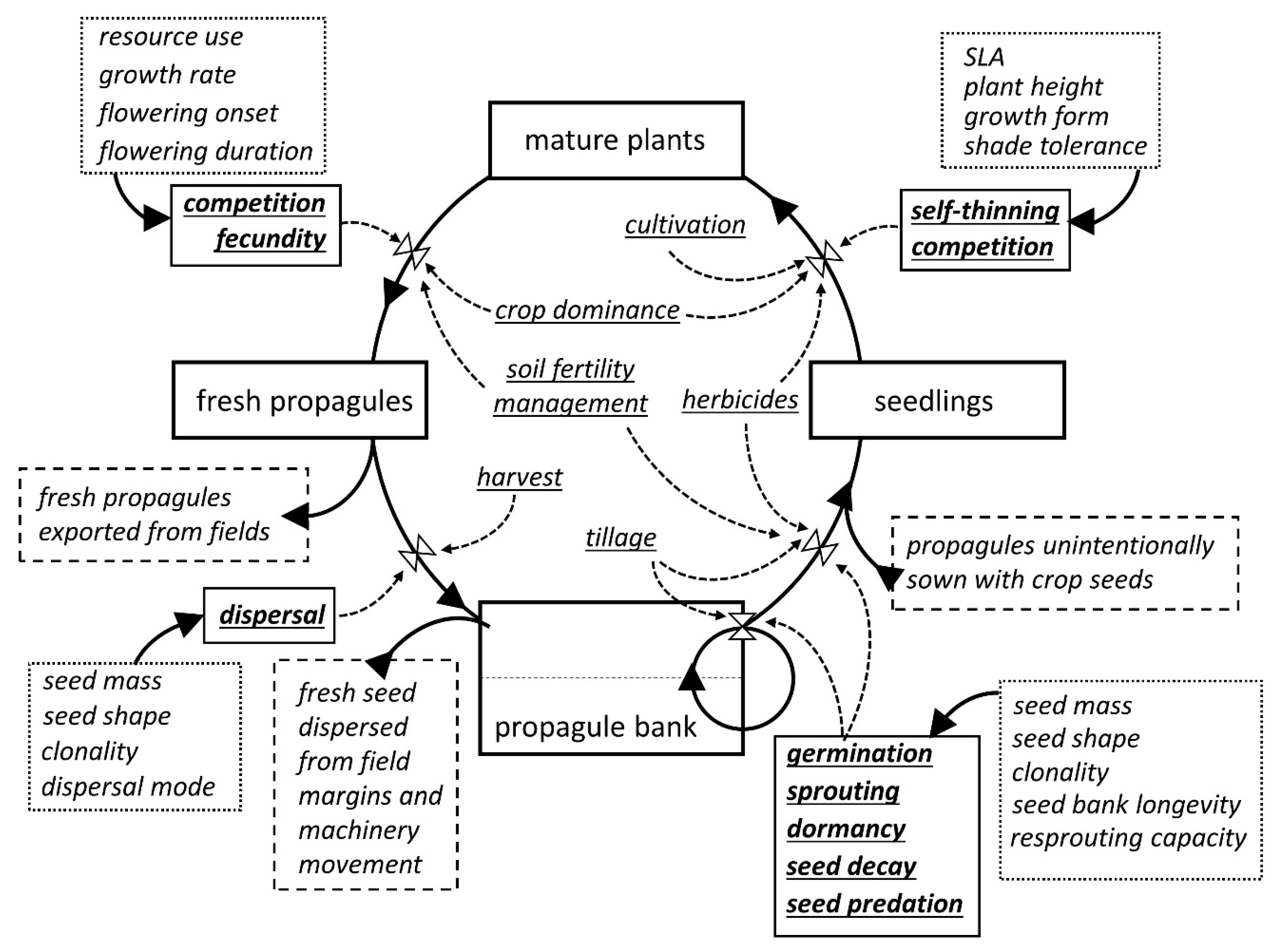
2. Modeling Weed Seed Dormancy, Germination, and Seedling Emergence
3. Weed–Crop Interference and Yield Loss Prediction Models
4. Gene Flow Models
4.1. Seed-Mediated Gene Flow
4.2. Pollen-Mediated Gene Flow
5. Weed Demography and Population Dynamic Models
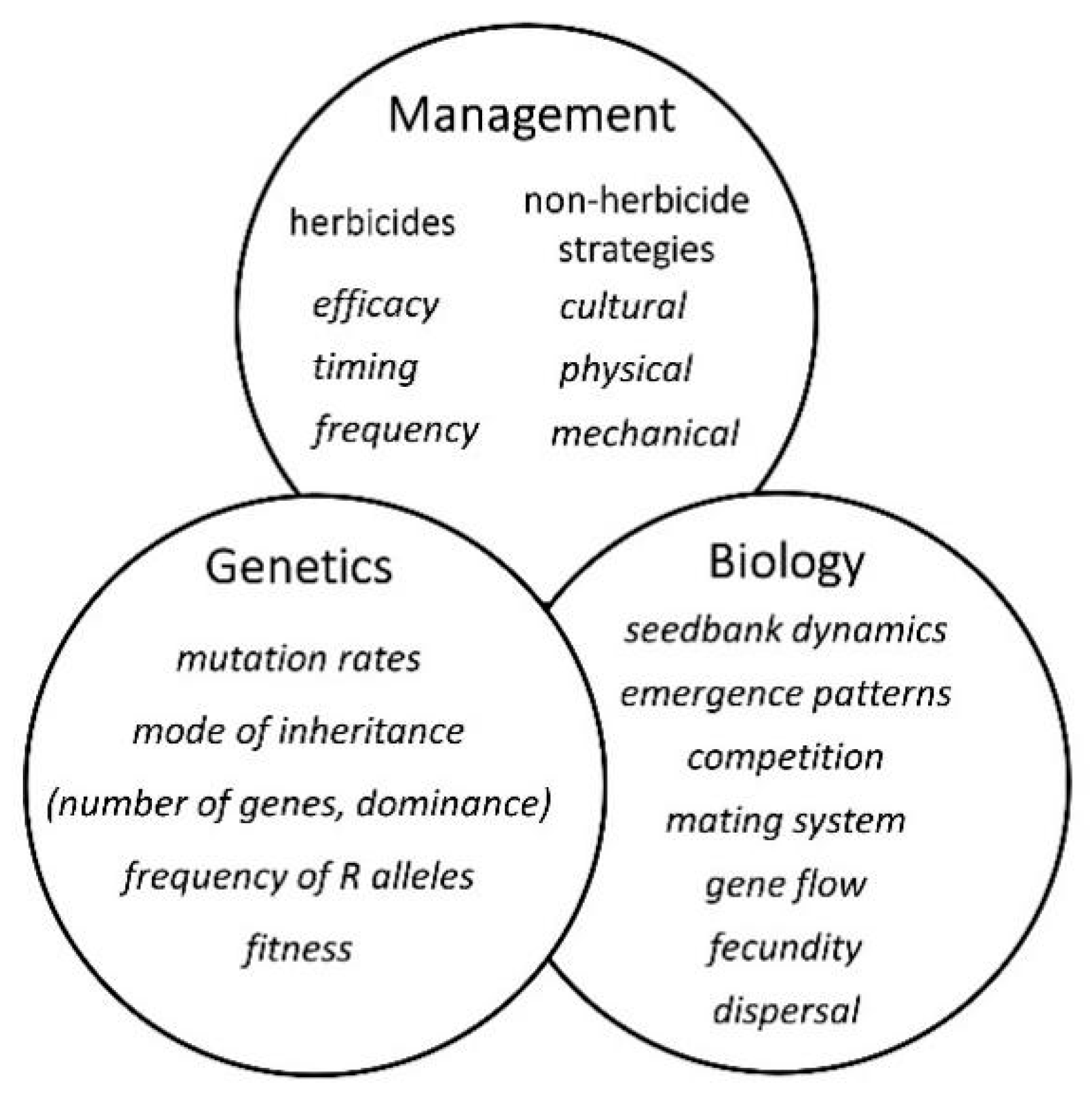
6. Herbicide Resistance Simulation Models
7. Management Decision-Support Tools
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Weed Crop Competition: A Review; International Plant Protection Center: Corvallis, OR, USA, 1980. [Google Scholar]
- Ballare, C.L.; Casal, J.J. Light signals perceived by crop and weed plants. Field Crops Res. 2000, 67, 149–160. [Google Scholar] [CrossRef]
- Page, E.R.; Tollenaar, M.; Lee, E.A.; Lukens, L.; Swanton, C.J. Shade avoidance: An integral component of crop-weed competition. Weed Res. 2010, 50, 281–288. [Google Scholar] [CrossRef]
- Bhowmik, P.C. Weed biology: Importance to weed management. Weed Sci. 1997, 45, 349–356. [Google Scholar] [CrossRef]
- Van Acker, R.C. Weed biology serves practical weed management. Weed Res. 2009, 49, 1–5. [Google Scholar] [CrossRef]
- Cousens, R. A simple model relating yield loss to weed density. Ann. Appl. Biol. 1985, 107, 239–252. [Google Scholar] [CrossRef]
- Radosevich, S.R. Methods to study interactions among crops and weeds. Weed Technol. 1987, 1, 190–198. [Google Scholar] [CrossRef]
- Kropff, M.J.; Lotz, L.A.P. Systems approaches to quantify crop-weed interactions and their application in weed management. Agric. Syst. 1992, 40, 265–282. [Google Scholar] [CrossRef]
- Kropff, M.J. Modelling the effects of weeds on crop production. Weed Res. 1988, 28, 465–471. [Google Scholar] [CrossRef]
- Holst, N.; Rasmussen, I.A.; Bastiaans, L. Field weed population dynamics: A review of model approaches and applications. Weed Res. 2007, 47, 1–14. [Google Scholar] [CrossRef]
- Haefner, J.W. Modeling Biological Systems: Principles and Applications, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Andrew, I.K.S.; Storkey, J. Using simulation models to investigate the cumulative effects of sowing rate, sowing date and cultivar choice on weed competition. Crop Prot. 2017, 95, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, C.J. Mathematical models in weed management. Crop Prot. 1991, 10, 432–444. [Google Scholar] [CrossRef]
- Lacoste, M.; Powles, S. Upgrading the RIM model for improved support of integrated weed management extension efforts in cropping systems. Weed Technol. 2014, 28, 703–720. [Google Scholar] [CrossRef] [Green Version]
- Neve, P. Simulation modelling to understand the evolution and management of glyphosate resistance in weeds. Pest Manag. Sci. 2008, 64, 392–401. [Google Scholar] [CrossRef]
- Bastiaans, L.; Storkey, J. Descriptive and Mechanistic Models of Crop-Weed Competition. In Weed Research: Expanding Horizons; Hatcher, P.E., Froud-Williams, R.J., Eds.; John Wiley & Sons: West Sussex, UK, 2017; Chapter 2; pp. 33–60. [Google Scholar]
- Colbach, N.; Collard, A.; Guyot, S.H.M.; Meziere, D.; Munier-Jolain, N. Assessing innovative sowing patterns for integrated weed management with a 3D crop: Weed competition model. Eur. J. Agron. 2014, 53, 74–89. [Google Scholar] [CrossRef]
- Kropff, M.J.; van Laar, H.H. Modelling Crop-weed Interactions; CAD International: Wallingford, UK, 1993. [Google Scholar]
- Park, E.S.; Laurence, R.B.; Andrew, R.W. The theory and application of plant competition models: An agronomic perspective. Ann. Bot. 2003, 92, 741–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renton, M.; Chauhan, B.S. Modelling crop-weed competition: Why, what, how and what lies ahead. Crop Prot. 2017, 95, 101–108. [Google Scholar] [CrossRef]
- Vleeshouwers, L.M.; Bouwmeester, H.J.; Karssen, C.M. Redefining seed dormancy: An attempt to integrate physiology and ecology. J. Ecol. 1995, 83, 1031–1037. [Google Scholar] [CrossRef] [Green Version]
- Benech-Arnold, R.L.; Sánchez, R.A.; Forcella, F.; Kruk, B.; Ghersa, C.M. Environmental control of dormancy in weed soil seed banks. Field Crops Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Vleeshouwers, L.M.; Kropff, M.J. Modelling field emergence patterns in arable weeds. New Phytol. 2000, 148, 445–457. [Google Scholar] [CrossRef]
- Batlla, D.; Benech-Arnold, R.L. Predicting changes in dormancy level in weed seed soil banks: Implications for weed management. Crop Prot. 2007, 26, 189–197. [Google Scholar] [CrossRef]
- Karssen, C.M. Seasonal patterns of dormancy in weed seeds. In The Physiology and Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier: Amsterdam, The Netherlands, 1982; pp. 243–270. [Google Scholar]
- Batlla, D.; Benech-Arnold, R.L. Predicting changes in dormancy level in natural seed soil banks. Plant Mol. Biol. 2010, 73, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Bradford, K.J. Threshold models applied to seed germination ecology. New Phytol. 2005, 165, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.J.; Karssen, C.M. The dual role of temperature in the regulation of the seasonal changes in dormancy and germination of seeds of Polygonum persicaria L. Oecologia 1992, 90, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.C.; Meyer, S.E.; Allen, P.S. A simulation model to predict seed dormancy loss in the field for Bromus tectorum L. J. Exp. Bot. 1998, 49, 1235–1244. [Google Scholar]
- Batlla, D.; Benech-Arnold, R.L. A quantitative analysis of dormancy loss dynamics in Polygonum aviculare L. seeds: Development of a thermal time model based on changes in seed population thermal parameters. Seed Sci. Res. 2003, 13, 55–68. [Google Scholar] [CrossRef]
- Gianinetti, A.; Cohn, M.A. Seed dormancy in red rice. XII: Population-based analysis of dry-after ripening with a hydrotime model. Seed Sci. Res. 2007, 17, 253–271. [Google Scholar] [CrossRef]
- Gardarin, A.; Dürr, C.; Colbach, N. Modeling the dynamics and emergence of a multispecies weed seed bank with species traits. Ecol. Model. 2012, 240, 123–138. [Google Scholar] [CrossRef]
- Bouwmeester, H.J.; Karssen, C.M. Annual changes in dormancy and germination in seeds of Sisymbrium officinale (L.) Scop. New Phytol. 1993, 124, 179–191. [Google Scholar] [CrossRef]
- Batlla, D.; Benech-Arnold, R.L. Seed dormancy loss assessed by changes in Polygonum aviculare L. population hydrotime parameters: Development of a predictive model. Seed Sci. Res. 2004, 14, 277–286. [Google Scholar] [CrossRef]
- Grundy, A.C. Predicting weed emergence: A review of approaches and future challenges. Weed Res. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Forcella, F.; Benech-Arnold, R.L.; Sanchez, R.E.; Ghersa, C.M. Modeling seedling emergence. Field Crop Res. 2000, 67, 123–139. [Google Scholar] [CrossRef]
- Izquierdo, J.; Bastida, F.; Lezaún, J.M.; Sánchez del Arco, M.J.; Gonzalez-Andujar, J.L. Development and evaluation of a model for predicting Lolium rigidum emergence in winter cereal crops in the Mediterranean area. Weed Res. 2013, 53, 269–278. [Google Scholar] [CrossRef]
- Werle, R.; Sandell, L.D.; Buhler, D.D.; Hartzler, R.G.; Lindquist, J.L. Predicting emergence of 23 summer annual weed species. Weed Sci. 2014, 62, 267–279. [Google Scholar] [CrossRef]
- Zambrano-Navea, C.; Bastida, F.; Gonzalez-Andujar, J.L. A hydrothermal seedling emergence model for Conyza bonariensis. Weed Res. 2013, 53, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Royo-Esnal, A.; Gesch, R.W.; Forcella, F.; Torra, J.; Recasens, J.; Necajeva, J. The role of light in the emergence of weeds: Using Camelina microcarpa as an example. PLoS ONE 2015, 10, e0146079. [Google Scholar] [CrossRef]
- Bewley, A.; BlackSeeds, M. Physiology of Development and Germination, 2nd ed.; Plenum Press: New York, NY, USA, 1994. [Google Scholar]
- Gonzalez-Andujar, J.L.; Chantre, G.R.; Morvillo, C.; Blanco, A.M.; Forcella, F. Predicting field weed emergence with empirical models and soft computing techniques. Weed Res. 2016, 56, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Francisco-Fernández, M.; Anand, A.; Bastida, F.; Gonzalez-Andujar, J.L. Computing statistical indices for hydrothermal times using weed emergence data. J. Agric. Sci. 2011, 149, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Kumar, A.; Das, B.; Burnwal, A. On soft computing techniques in various areas. Comp. Sci. Info. Tech. 2013, 3, 59–68. [Google Scholar]
- Onofri, A.; Piepho, H.P.; Kozak, M. Analyzing censored data in agricultural research: A review with examples and software tips. Ann. Appl. Biol. 2019, 174, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Onofri, A.; Gresta, F.; Tei, F. A new method for the analysis of germination and emergence data of weed species. Weed Res. 2010, 50, 187–198. [Google Scholar] [CrossRef]
- Hadi, M.R.H.S.; Gonzalez-Andujar, J.L. Comparison of fitting weed seedling emergence models with nonlinear regression and genetic algorithm. Comput. Elect. Agric. 2009, 65, 19–25. [Google Scholar] [CrossRef]
- Blanco, A.M.; Chantre, G.R.; Lodovichi, M.V.; Bandoni, J.A.; López, R.L.; Vigna, M.R.; Gigón, R.; Sabbatini, M.R. Modeling seed dormancy release and germination for predicting Avena fatua L. field emergence: A genetic algorithm approach. Ecol. Model. 2014, 272, 293–300. [Google Scholar] [CrossRef]
- Chantre, G.R.; Blanco, A.M.; Forcella, F.; Van Acker, R.C.; Sabbatini, M.R.; Gonzalez-Andujar, J.L. A comparative study between nonlinear regression and artificial neural network approaches for modeling wild oat (Avena fatua) field emergence. J. Agric. Sci. 2014, 52, 254–262. [Google Scholar] [CrossRef]
- Chantre, G.R.; Vigna, M.R.; Renzi, J.P.; Blanco, A.M. A flexible and practical approach for real-time weed emergence prediction based on Artificial Neural Networks. Biosyst. Eng. 2018, 170, 51–60. [Google Scholar] [CrossRef]
- Cao, R.; Francisco-Fernández, M.; Anand, A.; Bastida, F.; Gonzalez-Andujar, J.L. Modeling Bromus diandrus seedling emergence using nonparametric estimation. J. Agric. Biol. Environ. Stat. 2013, 18, 64–86. [Google Scholar] [CrossRef] [Green Version]
- Barrerios-Ures, D.; Francisco-Fernández, M.; Cao, R.; Fraguela, B.; Doallo, R.; Gonzalez-Andujar, J.L.; Reyes, M. Analysis of interval-grouped data in weed science: The binnednp Rcpp package. Ecol. Evol. 2019, 9, 10903–10915. [Google Scholar] [CrossRef]
- Sattin, M.; Zanin, G.; Berti, A. Case history for weed competition/population ecology: Velvetleaf (Abutilon theophrasti) in corn (Zea mays). Weed Technol. 1992, 6, 213–219. [Google Scholar] [CrossRef]
- Spitters, C.J.T.; Kropff, M.; de Groot, W. Competition between maize and Echinochloa crus-galli analysed by a hyperbolic regression model. Ann. Appl. Biol. 1989, 115, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Cousens, R. An empirical model relating crop yield to weed and crop density and a statistical comparison with other models. J. Agric. Sci. 1985, 105, 513–521. [Google Scholar] [CrossRef]
- Swinton, S.M.; Buhler, D.D.; Forcella, F.; Gunsolus, J.L.; King, R.P. Estimation of crop yield loss due to interference by multiple weed species. Weed Sci. 1994, 42, 103–109. [Google Scholar] [CrossRef]
- Christensen, S. Crop weed competition and herbicide performance in cereal species and varieties. Weed Res. 1994, 34, 29–36. [Google Scholar] [CrossRef]
- Kropff, M.J.; Spitters, J.T. A simple model of crop loss by weed competition from early observations on relative leaf area of the weeds. Weed Res. 1991, 31, 97–105. [Google Scholar] [CrossRef]
- Florez, J.A.; Fischer, A.J.; Ramirez, H.; Duque, M.C. Predicting rice yield losses caused by multispecies weed competition. Agron. J. 1999, 91, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Nieto, H.J.; Brondo, M.A.; Gonzales, M.A. Critical periods of the crop growth cycle for competition from weeds. PANS (C) 1968, 14, 159–166. [Google Scholar] [CrossRef]
- Kim, D.S.; Brain, P.; Marshall, E.J.P.; Caseley, J.C. Modelling herbicide dose and weed density effects on crop: Weed competition. Weed Res. 2002, 42, 1–13. [Google Scholar] [CrossRef]
- Weaver, S.E.; Kropff, M.J.; Groeneveld, R.M.W. Use of ecophysiological models for crop-weed interference: The critical period of weed interference. Weed Sci. 1992, 40, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Kropff, M.J.; Lotz, L.A.P.; Weaver, S.E.; Bos, H.J.; Wallinga, J.; Migo, T. A two-parameter model for prediction of crop loss by weed competition from early observations of relative leaf area of the weeds. Ann. Appl. Biol. 1995, 126, 329–346. [Google Scholar] [CrossRef] [Green Version]
- Deen, W.; Cousens, R.; Warringa, J.; Bastiaans, L.; Carberry, P.; Rebel, K.; Riha, S.; Murphy, C.; Benjamin, L.R.; Cloughley, C.; et al. An evaluation of four crop: Weed competition models using a common data set. Weed Res. 2003, 43, 116–129. [Google Scholar] [CrossRef] [Green Version]
- Snaydon, R.W. Replacement or additive designs for competition studies. J. Appl. Ecol. 1991, 28, 930–994. [Google Scholar] [CrossRef]
- Rejmanek, M.; Robinson, G.R.; Rejmankova, E. Weed-crop competition: Experimental designs and models for data analysis. Weed Sci. 1989, 37, 276–284. [Google Scholar] [CrossRef]
- Keller, M.; Gutjahr, C.; Mohring, J.; Weis, M.; Sokefeld, M.; Gerhards, R. Estimating economic thresholds for site-specific weed control using manual weed counts and sensor technology: An example based on three winter wheat trials. Pest Manag. Sci. 2014, 70, 200–211. [Google Scholar] [CrossRef]
- Bennett, A.C.; Price, A.J.; Sturgill, M.C.; Buol, G.S.; Wilkerson, G.C. HADSS™, Pocket HERB™, and Web HADSS™: Decision aids for field crops. Weed Technol. 2003, 17, 412–420. [Google Scholar] [CrossRef]
- Wilkerson, G.G.; Modena, S.A.; Coble, H.D. HERB: Decision model for postemergence weed control in soybean. Agron. J. 1991, 83, 413–417. [Google Scholar] [CrossRef]
- Barroso, J.; Fernandez-Quintanilla, C.; Maxwell, B.D.; Rew, L.J. Simulating the effects of weed spatial pattern and resolution of mapping and spraying on economics of site-specific management. Weed Res. 2004, 44, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Onyango, C.; Marchant, J.; Grundy, A.; Phelps, K.; Rader, R. Image processing performance assessment using crop weed competition models. Precis. Agric. 2005, 6, 183–192. [Google Scholar] [CrossRef]
- Apolo-Apolo, O.E.; Perez-Ruiz, M.; Martinez-Guanter, J.; Egea, G. A mixed data-based deep neural network to estimate leaf area index in wheat breeding trials. Agronomy 2020, 10, 175. [Google Scholar] [CrossRef] [Green Version]
- Bastiaans, L.; Kropff, M.J.; van Laar, H.H. Design of weed management systems with a reduced reliance on herbicides poses new challenges and prerequisites for modeling crop-weed interactions. Field Crops Res. 2000, 67, 161–179. [Google Scholar] [CrossRef]
- Gerhards, R.; Gutjahr, C.; Weis, M.; Keller, M.; Sökefeld, M.; Möhring, J.; Piepho, H.P. Using precision farming technology to quantify yield effects attributed to weed competition and herbicide application. Weed Res. 2012, 52, 6–15. [Google Scholar] [CrossRef]
- Colbach, N.; Forcella, F.; Johnson, G.A. Spatial and temporal stability of weed populations over five years. Weed Sci. 2000, 48, 366–377. [Google Scholar] [CrossRef]
- Graf, B.; Gutierrez, A.P.; Rakotobe, O.; Zahner, P.; Delucchi, V. A simulation model for the dynamics of rice growth and development: Part II-The competition with weeds for nitrogen and light. Agric. Syst. 1990, 32, 367–392. [Google Scholar] [CrossRef]
- Afifi, M.; Swanton, C. Early physiological mechanisms of weed competition. Weed Sci. 2012, 60, 542–551. [Google Scholar] [CrossRef]
- Ziska, L.H. Could recent increases in atmospheric CO2 have acted as a selection factor in Avena fatua populations A case study of cultivated and wild oat competition. Weed Res. 2017, 57, 399–405. [Google Scholar] [CrossRef]
- Ziska, L.H.; Tomecek, M.B.; Gealy, D.R. Competitive interactions between cultivated and red rice as a function of recent and projected increases in atmospheric carbon dioxide. Agron. J. 2010, 102, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Bravo, W.; Leon, R.G.; Ferrell, J.A.; Mulvaney, M.J.; Wood, C.W. Evolutionary adaptations of Palmer amaranth populations (Amaranthus palmeri) to nitrogen fertilization and crop rotation history affect morphology and nutrient-use efficiency. Weed Sci. 2018, 66, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Bravo, W.; Leon, R.G.; Ferrell, J.A.; Mulvaney, M.J.; Wood, C.W. Differentiation of life-history traits among Palmer amaranth populations (Amaranthus palmeri) and its relation to cropping systems and glyphosate sensitivity. Weed Sci. 2017, 65, 339–349. [Google Scholar] [CrossRef]
- Beckie, H.J.; Busi, R.; Bagavathiannan, M.V.; Martin, S.L. Herbicide resistance gene flow in weeds: Under-estimated and under-appreciated. Agric. Ecosyst. Environ. 2019, 283, e106566. [Google Scholar] [CrossRef]
- Beckie, H.J.; Hall, L.M. Simple to complex: Modelling crop pollen-mediated gene flow. Plant Sci. 2008, 175, 615–628. [Google Scholar] [CrossRef]
- Cousens, R.; Dytham, C.; Law, R. Dispersal in Plants: A Population Perspective; Oxford University Press: Oxford, UK, 2008; 240p. [Google Scholar]
- Dauer, J.T.; Mortensen, D.A.; Vangessel, M.J. Temporal and spatial dynamics of long-distance Conyza canadensis seed dispersal. J. Appl. Ecol. 2006, 44, 105–114. [Google Scholar] [CrossRef]
- Andersen, M.C. An analysis of variability in seed settling velocities of several wind-dispersed Asteraceae. Am. J. Bot. 1992, 79, 1087–1091. [Google Scholar] [CrossRef]
- Stephenson, C.M.; Kohn, D.D.; Park, K.J.; Atkinson, R.; Edwards, C.; Travis, J.M. Testing mechanistic models of seed dispersal for the invasive Rhododendron ponticum (L.). Perspect. Plant Ecol. Evol. Syst. 2007, 9, 15–28. [Google Scholar] [CrossRef]
- Nathan, R.; Katul, G.G.; Bohrer, G.; Kuparinen, A.; Soons, M.B.; Thompson, S.E.; Trakhtenbrot, A.; Horn, H.S. Mechanistic models of seed dispersal by wind. Theor. Ecol. 2011, 4, 113–132. [Google Scholar] [CrossRef]
- Ganie, Z.A.; Jhala, A.J. Modeling pollen-mediated gene flow from glyphosate-resistant to –susceptible giant ragweed (Ambrosia trifida L.) under field conditions. Sci. Rep. 2017, 7, e17067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, E.K.; Lavigne, C.; Picault, H.; Renard, M.; Gouyon, P.H. Pollen dispersal of oilseed rape: Estimation of the dispersal function and effects of field dimension. J. Appl. Ecol. 2006, 43, 141–151. [Google Scholar] [CrossRef]
- Devaux, C.; Lavigne, C.; Austerlitz, F.; Klein, E.K. Modelling and estimating pollen movement in oilseed rape (Brassica napus) at the landscape scale using genetic markers. Mol. Ecol. 2007, 16, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.W.; Harwood, T.D.; Wilkinson, M.J.; Elliott, L. Assembling spatially explicit landscape models of pollen and spore dispersal by wind for risk assessment. Proc. Roy. Soc. B 2006, 273, 1705–1713. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, G. Pollen dispersal vectored by wind or insects. In Gene Flow from GM Plants; Poppy, G.M., Wilkinson, M.J., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2005. [Google Scholar]
- Jackson, S.T.; Lyford, M.E. Pollen dispersal models in quaternary plant ecology: Assumptions, parameters, and prescriptions. Bot Rev. 1999, 65, 39–75. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Germination ecophysiology of herbaceous plant species in a temperature region. Am. J. Bot. 1988, 75, 286–305. [Google Scholar] [CrossRef]
- Burnside, O.C.; Wilson, R.G.; Weisberg, S.; Hubbard, K.G. Seed longevity of 41 weed species buried 17 years in eastern and western Nebraska. Weed Sci. 1996, 44, 74–86. [Google Scholar] [CrossRef]
- Buhler, D.D.; Hartzler, R.G. Emergence and persistence of seed of velvetleaf, common waterhemp, woolly cupgrass, and giant foxtail. Weed Sci. 2001, 49, 230–235. [Google Scholar] [CrossRef]
- Forcella, F.; Wilson, R.G.; Dekker, J.; Kremer, R.; Cardina, J.; Anderson, R.L.; Alm, D.; Renner, K.A.; Harvey, R.G.; Clay, S.; et al. Weed seed bank emergence across the Corn Belt. Weed Sci. 1997, 67, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Bagavathiannan, M.V.; Norsworthy, J.K. Late-Season seed production in arable weed communities: Management implications. Weed Sci. 2012, 60, 325–334. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Korres, N.E.; Bagavathiannan, M.V. Weed seedbank management: Revisiting how herbicides are evaluated. Weed Sci. 2018, 66, 415–417. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.S.; Dixon, P.M.; Liebman, M. Cropping system effects on giant foxtail demography: II. retrospective perturbation analysis. Weed Sci. 2003, 51, 930–939. [Google Scholar] [CrossRef]
- Liebman, M.; Nichols, V.A. Cropping system redesign for improved weed management: A modeling approach illustrated with giant ragweed (Ambrosia trifida). Agronomy 2020, 10, 262. [Google Scholar] [CrossRef] [Green Version]
- Jordan, N.; Mortensen, D.A.; Prenzlow, D.M.; Cox, K.C. Simulation analysis of crop rotation effects on weed seedbanks. Am. J. Bot. 1995, 82, 390–398. [Google Scholar] [CrossRef]
- Bohan, D.A.; Powers, S.J.; Champion, G.; Haughton, A.J.; Hawes, C.; Squire, G.; Cussans, J.; Mertens, S.K. Modelling rotations: Can crop sequences explain arable weed seedbank abundance. Weed Res. 2011, 51, 422–432. [Google Scholar] [CrossRef]
- Munier-Jolain, N.M.; Guyot, S.H.M.; Colbach, N. A 3D model for light interception in heterogeneous crop: Weed canopies. Model structure and evaluation. Ecol. Model. 2013, 250, 101–110. [Google Scholar] [CrossRef]
- Storkey, J.; Moss, S.R.; Cussans, J.W. Using assembly theory to explain changes in a weed flora in response to agricultural intensification. Weed Sci. 2010, 58, 39–46. [Google Scholar] [CrossRef]
- Colbach, N.; Gardarin, A.; Munier-Jolain, N.M. FLORSYS: A mechanistic model of cropping system effects on weed flora based on functional relationships with species traits. In Proceedings of the 15th International EWRS Symposium, Kaposvar, Hungary, 12–15 July 2010. [Google Scholar]
- Gressel, J.; Segel, L.A. The paucity of plants evolving genetic resistance to herbicides: Possible reasons and implications. J. Theoret. Biol. 1978, 75, 349–371. [Google Scholar] [CrossRef]
- Maxwell, B.D.; Roush, M.L.; Radosevich, S.R. Predicting the evolution and dynamics of herbicide resistance in weed populations. Weed Technol. 1990, 4, 2–13. [Google Scholar] [CrossRef]
- Jasieniuk, M.; Maxwell, B.D. Population genetics and the evolution of herbicide resistance in weeds. Phytoprotection 1994, 75, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Renton, M.; Diggle, A.; Manalil, S.; Powles, S. Does cutting herbicide rates threaten the sustainability of weed management in cropping systems. J. Theor. Biol. 2011, 283, 14–27. [Google Scholar] [CrossRef]
- Neve, J.K.; Norsworthy, K.L.; Smith, I.A. Modelling evolution and management of glyphosate resistance in Amaranthus palmeri. Weed Res. 2011, 51, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Bagavathiannan, M.V.; Norsworthy, J.K.; Smith, K.L.; Neve, P. Modeling the evolution of glyphosate resistance in barnyardgrass (Echinochloa crus-galli) in cotton-based production systems of the Mid southern United States. Weed Technol. 2013, 27, 475–487. [Google Scholar] [CrossRef]
- Bagavathiannan, M.V.; Norsworthy, J.K.; Smith, K.L.; Neve, P. Modeling the simultaneous evolution of resistance to ALS- and ACCase-inhibiting herbicides in barnyardgrass (Echinochloa crus-galli) in Clearfield® rice. Weed Technol. 2014, 28, 89–103. [Google Scholar] [CrossRef]
- Werle, R.; Brigitte, T.; Lindquist, J.L. Modeling shattercane dynamics in herbicide-tolerant grain sorghum cropping systems. Ecol. Model. 2017, 343, 131–141. [Google Scholar] [CrossRef]
- Heap, I. International Survey of Herbicide Resistant Weeds. 2020. Available online: www.herbicideresistance.org (accessed on 28 September 2020).
- Mortimer, A.M.; Ulf-Hansen, P.F.; Putwain, P.D. Modelling herbicide resistance-a study of ecological fitness. In Achievements and Developments in Combating Pesticide Resistance; Denholm, I., Devonshire, A.L., Hollomons, D.W., Eds.; Elsevier Science Publishers: Essex, UK, 1992; pp. 283–306. [Google Scholar]
- Neve, P.; Diggle, A.J.; Smith, F.P.; Powles, S.B. Simulating evolution of glyphosate resistance in Lolium rigidum I: Population biology of a rare resistance trait. Weed Res. 2003, 43, 404–417. [Google Scholar] [CrossRef]
- Renton, M.; Busi, R.; Neve, P.; Thornby, D.; Vila-Aiub, M. Herbicide resistance modelling: Past, present and future. Pest Manag. Sci. 2014, 70, 1394–1404. [Google Scholar] [CrossRef]
- Diggle, A.J.; Neve, P.B.; Smith, F.P. Herbicides used in combination can reduce the probability of herbicide resistance in finite weed populations. Weed Res. 2003, 43, 371–382. [Google Scholar] [CrossRef]
- Powles, S.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annl. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [Green Version]
- Pannell, D.J.; Stewart, V.; Bennett, A.; Monjardino, M.; Schmidt, C.; Powles, S.B. RIM: A bioeconomic model for integrated weed management of Lolium rigidum in Western Australia. Agric. Syst. 2004, 79, 305–325. [Google Scholar] [CrossRef]
- Somerville, G.J.; Powles, S.B.; Walsh, M.J.; Renton, M. Modeling the impact of harvest weed seed control on herbicide resistance evolution. Weed Sci. 2018, 66, 395–403. [Google Scholar] [CrossRef]
- Liu, C.; Bridges, M.E.; Kaundun, S.S.; Glasgow, L.; Owen, M.D.K.; Neve, P. A generalized individual-based algorithm for modelling the evolution of quantitative herbicide resistance in arable weed populations. Pest Manag. Sci. 2017, 73, 462–474. [Google Scholar] [CrossRef]
- Gardner, S.; Gressel, J.; Mangel, M. A revolving dose strategy to delay the evolution of both quantitative vs. major monogene resistances to pesticides and drugs. Int. J. Pest. Manag. 1998, 44, 161–180. [Google Scholar] [CrossRef]
- Richter, O.; Zwerger, P.; Bottcher, U. Modelling spatio-temporal dynamics of herbicide resistance. Weed Res. 2002, 42, 52–64. [Google Scholar] [CrossRef]
- Somerville, G.J.; Powles, S.B.; Walsh, M.J.; Renton, M. How do spatial heterogeneity and dispersal in weed population models affect predictions of herbicide resistance evolution? Ecol. Model. 2017, 362, 37–53. [Google Scholar] [CrossRef]
- Vila-Aiub, M.; Neve, P.; Powles, S. Fitness costs associated with evolved herbicide resistance alleles in plants. New Phytol. 2009, 184, 751–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somerville, G.J.; Sønderskov, M.; Mathiassen, S.K.; Metcalfe, H. Spatial modelling of within-field weed populations: A review. Agronomy 2020, 10, 1044. [Google Scholar] [CrossRef]
- Busi, R.; Gaines, T.A.; Powles, S. Phorate can reverse P450 metabolism-based herbicide resistance in Lolium rigidum. Pest Manag. Sci. 2017, 73, 410–417. [Google Scholar] [CrossRef]
- Wiles, L.J.; King, R.P.; Sweizer, E.E.; Lybecker, D.W.; Swinton, S.M. GWM: General weed management model. Agric. Syst. 1996, 50, 355–376. [Google Scholar] [CrossRef]
- Parsons, D.J.; Benjamin, L.R.; Clarke, J.; Ginsburg, D.; Mayes, A.; Milne, A.E.; Wilkinson, D.J. Weed Manager—A model-based decision support system for weed management in arable crops. Comput. Electron. Agric. 2009, 65, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Oriade, C.; Forcella, F. Maximizing efficacy and economics of mechanical weed control in row crops through forecasts of weed emergence. In Expanding the Context of Weed Management; Food Products Press: Binghamton, NY, USA, 1999; pp. 189–206. [Google Scholar]
- Scursoni, J.A.; Forcella, F.; Gunsolus, J. Weed escapes and delayed weed emergence in glyphosate-resistant soybean. Crop Prot. 2007, 26, 212–218. [Google Scholar] [CrossRef]
- Schutte, B.J.; Hager, A.C.; Davis, A.S. Respray requests on custom-applied, glyphosate-resistant soybeans in Illinois: How many and why. Weed Technol. 2010, 24, 590–598. [Google Scholar] [CrossRef]
- Nowell, L.H.; Moran, P.W.; Schmidt, T.S.; Norman, J.E.; Nakagaki, N.; Shoda, M.E.; Mahler, B.J.; Van Metre, P.C.; Stone, W.W.; Sandstrom, M.W.; et al. Complex mixtures of dissolved pesticides show potential aquatic toxicity in a synoptic study of Midwestern U.S. streams. Sci. Total Environ. 2018, 613–614, 1469–1488. [Google Scholar] [CrossRef]
- Pandey, S.; Medd, R.W. A stochastic dynamic programming framework for weed control decision making: An application to Avena fatua L. Agric. Econ. 1991, 6, 115–128. [Google Scholar] [CrossRef]
- Lodovichi, M.V.; Blanco, A.M.; Chantre, G.R.; Bandoni, J.A.; Sabbatini, M.R.; Vigna, M.; López, R.; Gigón, R. Operational planning of herbicide-based weed management. Agric. Syst. 2013, 121, 117–129. [Google Scholar] [CrossRef]
- Berti, A.; Bravin, F.; Zanin, G. Application of decision-support software for postemergence weed control. Weed Sci. 2003, 51, 618–627. [Google Scholar] [CrossRef]
- Wilkerson, G.C.; Wiles, L.J.; Bennett, A.C. Weed management decision models: Pitfalls, perceptions, and possibilities of the economic threshold approach. Weed Sci. 2002, 50, 411–424. [Google Scholar] [CrossRef]
- Lindsay, K.; Popp, M.; Norsworthy, J.; Bagavathiannan, M.; Powles, S.; Lacoste, M. PAM: Decision support for long-term Palmer amaranth (Amaranthus palmeri) control. Weed Technol. 2017, 31, 915–927. [Google Scholar] [CrossRef]
- Lacoste, M.; Powles, S. Beyond modeling: Considering user-centered and post-development aspects to ensure the success of a decision support system. Comput. Electron. Agric. 2016, 121, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Kragt, M.E.; Llewellyn, R.S. Using a choice experiment to improve decision support tool design. Appl. Econ. Perspect. Policy 2014, 36, 351–371. [Google Scholar] [CrossRef]
- Colas, F.; Cordeau, S.; Granger, S.; Jeuffroy, M.-H.; Pointurier, O.; Queyrel, W.; Rodriguez, A.; Villerd, J.; Colbach, N. Co-development of a decision support system for integrated weed management: Contribution from future users. Eur. J. Agron. 2020, 114, 126010. [Google Scholar] [CrossRef]
- Bessette, D.; Wilson, R.; Beaudrie, C.; Schroeder, C. An online decision support tool to evaluate ecological weed management strategies. Weed Sci. 2019, 67, 463–473. [Google Scholar] [CrossRef]
- Kristensen, K.; Rasmussen, I.A. The use of Bayesian network in the design of a decision support system for growing malting barley without use of pesticides. Comput. Electron. Agric. 2002, 32, 197–217. [Google Scholar] [CrossRef]
- Neuhoff, D.; Schulz, D.; Köpke, U. Potential of decision support systems for organic crop production: WECOF-DSS, a tool for weed control in winter wheat. In Proceedings of the International Scientific Conference on Organic Agriculture, Adelaide, Australia, 21–23 September 2005. [Google Scholar]
- Zambrano-Navea, C.; Bastida, F.; Gonzalez-Andujar, J.L. A cohort-based stochastic model of the population dynamic and long-term management of Conyza bonariensis in fruiting tree crops. Crop Prot. 2016, 80, 15–20. [Google Scholar] [CrossRef]
- Sarangi, D.; Jhala, A.J. Biologically effective rates of a new premix (atrazine, bicyclopyrone, mesotrione, and S-metolachlor) for preemergence or postemergence control of common waterhemp [Amaranthus tuberculatus (Moq.) Sauer var. rudis] in corn. Can. J. Plant Sci. 2017, 97, 1075–1089. [Google Scholar]
- Dieleman, J.A.; Mortensen, D.A.; Martin, A.R. Influence of velvetleaf (Abutilon theophrasti) and common sunflower (Helianthus annuus) density variation on weed management outcomes. Weed Sci. 1999, 47, 81–89. [Google Scholar] [CrossRef]
- Schutte, B.J. Measuring interference from midseason tall morning glory (Ipomoea purpurea) to develop a model for teaching weed seedbank effects on chile pepper. Weed Technol. 2017, 31, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Renner, K.A.; Black, J.R. SOYHERB-A computer program for soybean herbicide decision making. Agron. J. 1991, 83, 921–925. [Google Scholar] [CrossRef]
- Lyon, L.L.; Keeling, J.W.; Dotry, P.A. Evaluation and adaptation of the HADSS® computer program in Texas Southern High Plains cotton. Weed Technol. 2004, 18, 315–324. [Google Scholar] [CrossRef]
- Ford, A.J.; Dotray, P.A.; Keeling, J.W.; Wilkerson, J.B.; Wilcut, J.W.; Gilbert, L.V. Site-specific weed management in cotton using WebHADSS™. Weed Technol. 2011, 25, 107–112. [Google Scholar] [CrossRef]
- Gonzalez-Andujar, J.L.; Fernandez-Quintanilla, C.; Bastida, F.; Calvo, R.; Izquierdo, J.; Lezaun, J.A. Assessment of a decision support system for chemical control of annual ryegrass (Lolium rigidum) in winter cereals. Weed Res. 2011, 51, 304–309. [Google Scholar] [CrossRef]
- Gonzalez-Andujar, J.L.; Fernandez-Quintanilla, C.; Bastida, F.; Calvo, R.; Gonzalez-Diaz, L.; Izquierdo, J.; Lezaun, J.A.; Perea, F.; Sanchez del Arco, M.J.; Urbano, J.M. Field evaluation of a decision support system for herbicidal control of Avena sterilis ssp. ludoviciana in winter wheat. Weed Res. 2010, 50, 83–88. [Google Scholar] [CrossRef]
- Meyer, C.J.; Norsworthy, J.K. Influence of weed size on herbicide interactions for Enlist™ and Roundup Ready® Xtend® technologies. Weed Technol. 2019, 33, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Knezevic, S.Z.; Evans, S.P.; Blankenship, E.E.; Van Acker, R.C. Critical period for weed control: The concept and data analysis. Weed Sci. 2002, 50, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, L.R.; Milne, A.E.; Parsons, D.J.; Cussans, J.; Lutman, P.J.W. Using stochastic dynamic programming to support weed management decisions over a rotation. Weed Res. 2009, 49, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Lacoste, M.; Powles, S. RIM: Anatomy of a weed management decision support system for adaptation and wider application. Weed Sci. 2015, 63, 676–689. [Google Scholar] [CrossRef]
- Cousens, R.; Doyle, C.J.; Wilson, B.J.; Cussans, G.W. Modelling the economics of controlling Avena fatua in winter wheat. Pestic. Sci. 1986, 17, 1–12. [Google Scholar] [CrossRef]
- Gonzalez-Diaz, L.; Bastida, F.; Gonzalez-Andujar, J.L. A bioeconomic model for the analysis of control strategies for Lolium rigidum and Avena sterilis ssp. ludoviciana in winter wheat. Int. J. Plant Prod. 2020, 14, 37–42. [Google Scholar] [CrossRef]
- Stanton, R.A.; Pratley, J.E.; Hudson, D.; Dill, G.M. A risk calculator for glyphosate resistance in Lolium rigidum (Gaud.). Pest Manag. Sci. 2008, 64, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Beckie, H.J.; Harker, N.K.; Hall, L.M.; Holm, F.A.; Gulden, R.H. Risk assessment of glyphosate resistance in western Canada. Weed Technol. 2011, 25, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Borger, C.P.D.; Riethmuller, G.P.; Renton, M. Weed Seed Wizard: A tool that demonstrates the value of integrated weed management tactics such as harvest weed seed destruction. Comput. Electron. Agric. 2018, 147, 27–33. [Google Scholar] [CrossRef]
- Llewellyn, R.S.; Pannell, D.J.; Lindner, R.K.; Powles, S.B. Targeting key perceptions when planning and evaluating extension. Aust. J. Exp. Agric. 2005, 45, 1627–1633. [Google Scholar] [CrossRef] [Green Version]
- Attonaty, J.M.; Chatelin, M.H.; Garcia, F. Interactive simulation modeling in farm decision-making. Comput. Electron. Agric. 1999, 22, 157–170. [Google Scholar] [CrossRef]
- Neve, P.; Norsworthy, J.K.; Smith, K.L.; Zelaya, I.A. Modeling glyphosate resistance management strategies for Palmer amaranth (Amaranthus palmeri) in cotton. Weed Technol. 2011, 25, 335–343. [Google Scholar] [CrossRef]
- Jørgensen, L.N.; Noe, E.; Langvad, A.M.; Jensen, J.E.; Ørum, J.E.; Rydahl, P. Decision support systems: Barriers and farmers’ need for support. Bull. OEPP 2007, 37, 374–377. [Google Scholar] [CrossRef]
- Kanatas, P.; Travlos, I.S.; Gazoulis, I.; Tataridas, A.; Tsekoura, A.; Antonopoulos, N. Benefits and limitations of decision support systems (DSS) with a special emphasis on weeds. Agronomy 2020, 10, 548. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Model Characteristic | DST for Curative Tactics | DST for Preventive Management |
|---|---|---|
| Projected output | Short-term control outcomes and current season profit responses for possible tactics for controlling weeds | Weed infestation and revenue responses to different combinations of tactics occurring over multiple years |
| Time frame | Single growing season | Multiple growing seasons |
| Information needed for development |
|
|
| Primary service to farmers and farm advisors | Prescriptive recommendations for specific weed problems | Means for investigating novel, multiyear strategies for managing weed communities or problematic weed biotypes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagavathiannan, M.V.; Beckie, H.J.; Chantre, G.R.; Gonzalez-Andujar, J.L.; Leon, R.G.; Neve, P.; Poggio, S.L.; Schutte, B.J.; Somerville, G.J.; Werle, R.; et al. Simulation Models on the Ecology and Management of Arable Weeds: Structure, Quantitative Insights, and Applications. Agronomy 2020, 10, 1611. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101611
Bagavathiannan MV, Beckie HJ, Chantre GR, Gonzalez-Andujar JL, Leon RG, Neve P, Poggio SL, Schutte BJ, Somerville GJ, Werle R, et al. Simulation Models on the Ecology and Management of Arable Weeds: Structure, Quantitative Insights, and Applications. Agronomy. 2020; 10(10):1611. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101611
Chicago/Turabian StyleBagavathiannan, Muthukumar V., Hugh J. Beckie, Guillermo R. Chantre, Jose L. Gonzalez-Andujar, Ramon G. Leon, Paul Neve, Santiago L. Poggio, Brian J. Schutte, Gayle J. Somerville, Rodrigo Werle, and et al. 2020. "Simulation Models on the Ecology and Management of Arable Weeds: Structure, Quantitative Insights, and Applications" Agronomy 10, no. 10: 1611. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101611