Antifungal Effects of Rhizospheric Bacillus Species Against Bayberry Twig Blight Pathogen Pestalotiopsis versicolor

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Fungal Pathogen and Growth Medium
2.2. Isolation of Rhizospheric Bacteria
2.3. Screening for Antagonistic Activity Against P. versicolor XJ27
2.4. Identification of Potential Antagonistic Bacteria
2.5. Antagonistic Efficacy of Extracellular Culture Filtrates and Their Thermal Stability Assay
2.6. Hyphal Structures Observation Using SEM and TEM
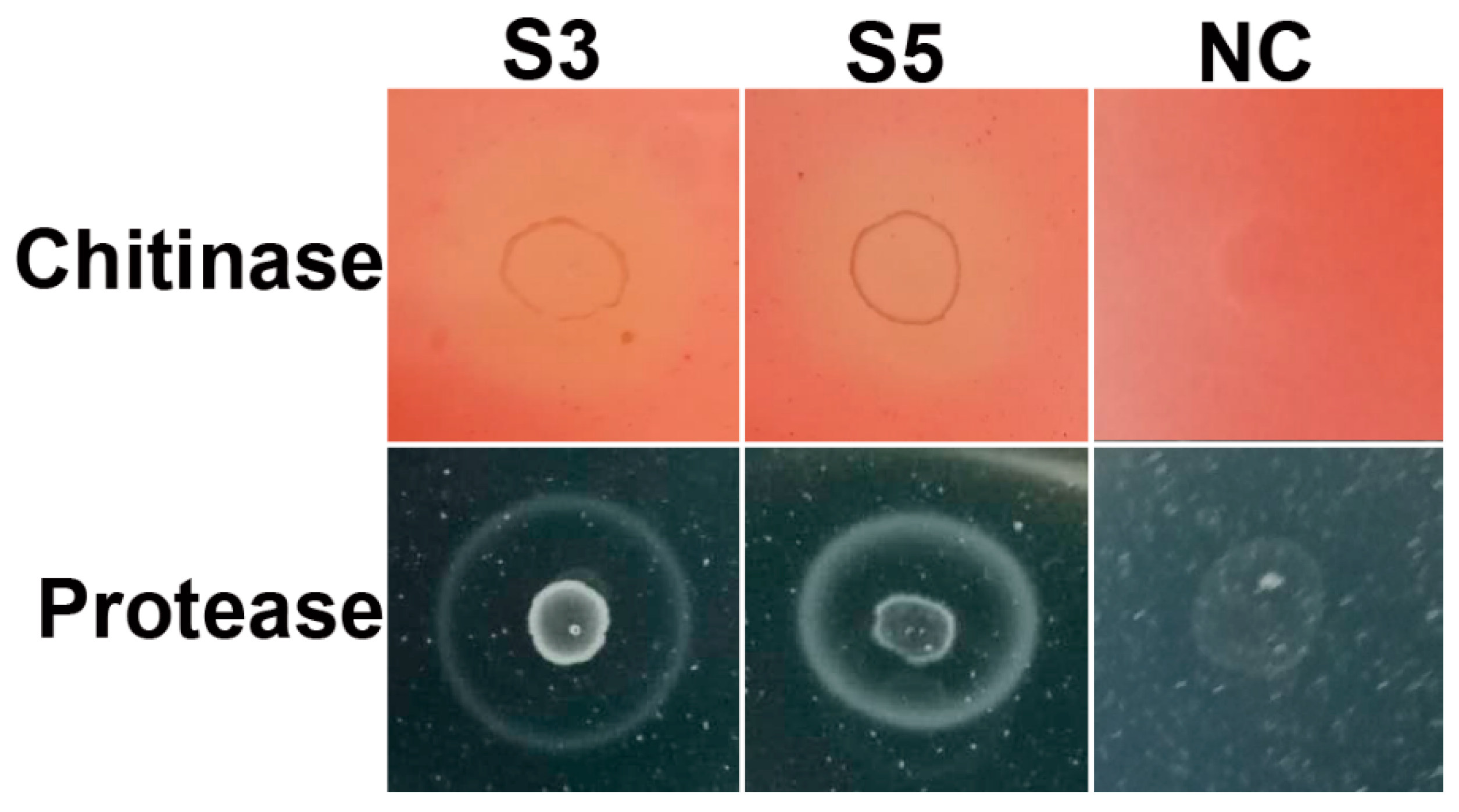
2.7. Hydrolytic Enzymes Activity Assay
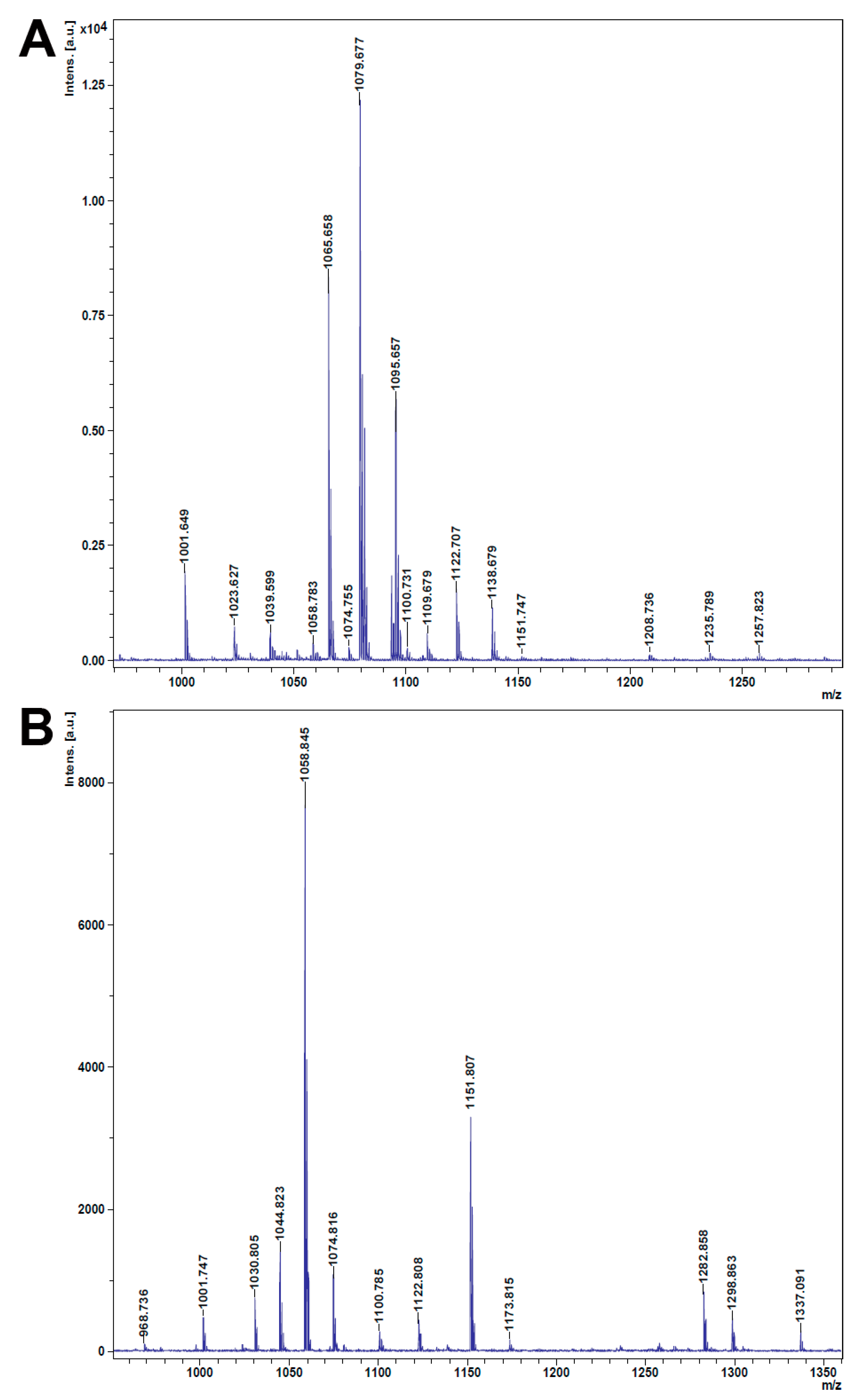
2.8. Detection of Lipopeptides in Antagonistic Bacteria by MALDI–TOF-MS Analysis
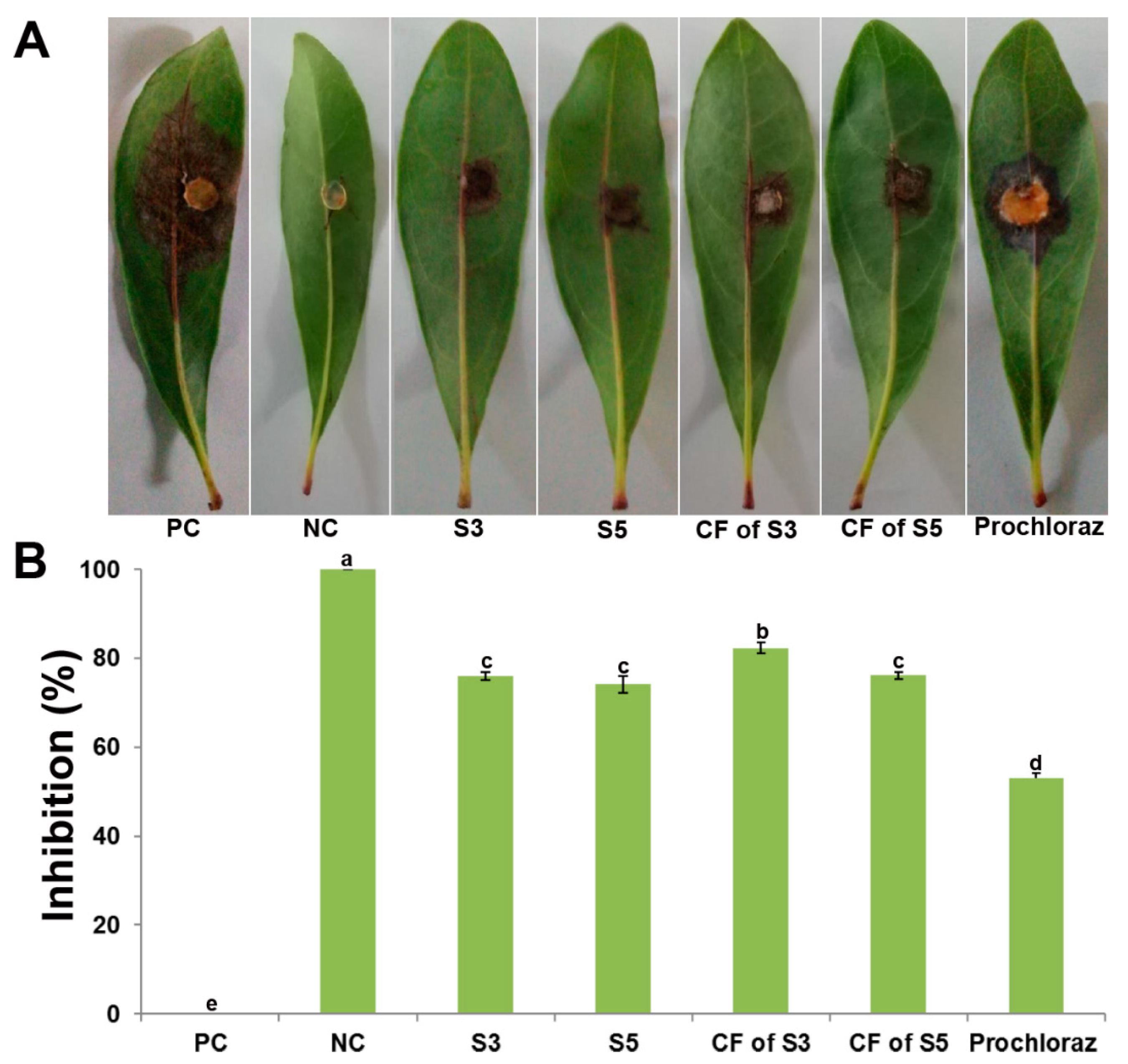
2.9. Antagonistic Assay on Detached Leaf Against P. versicolor XJ27
2.10. Statistical Analysis
3. Results
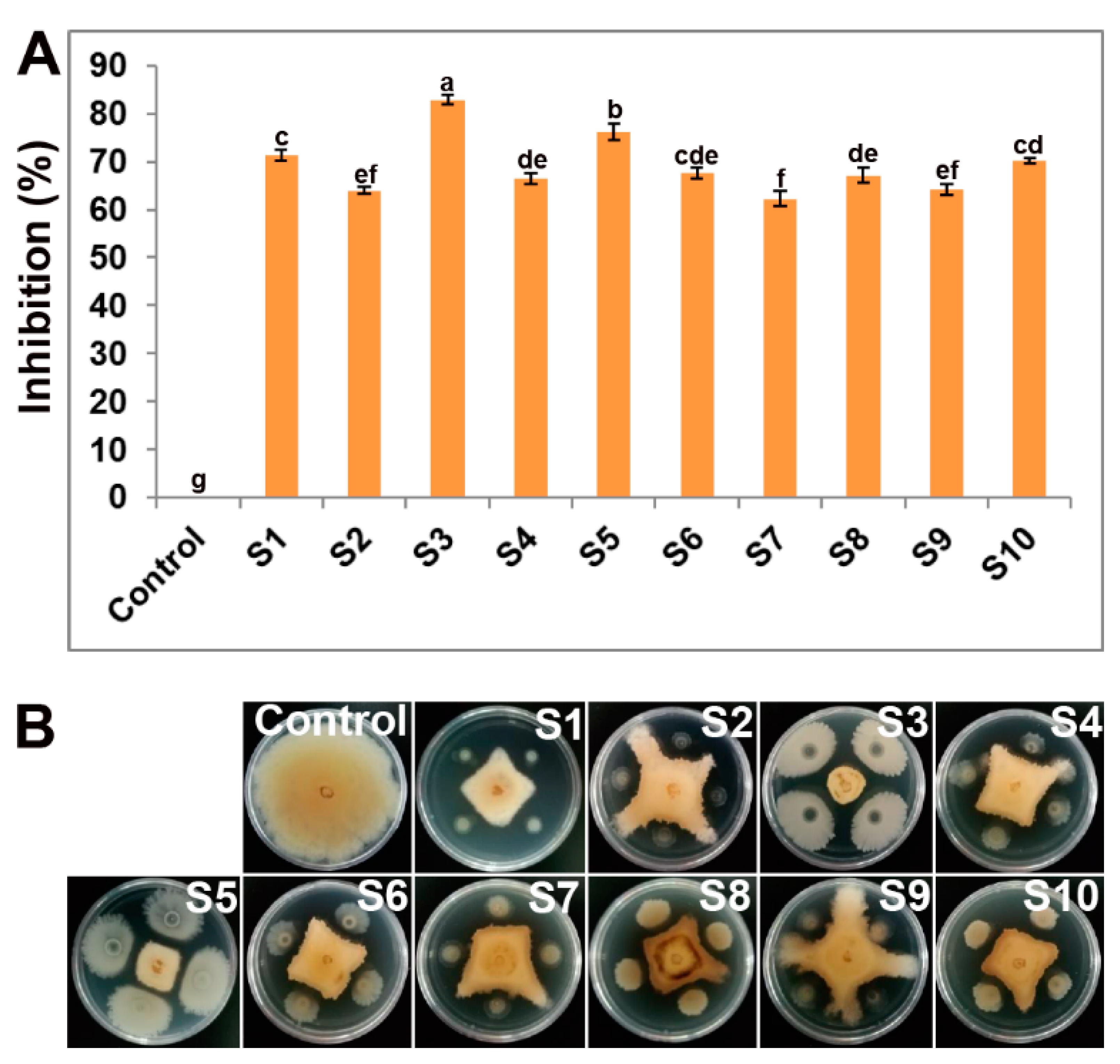
3.1. Rhizospheric Bacteria Showed XJ27 Inhibition In Vitro
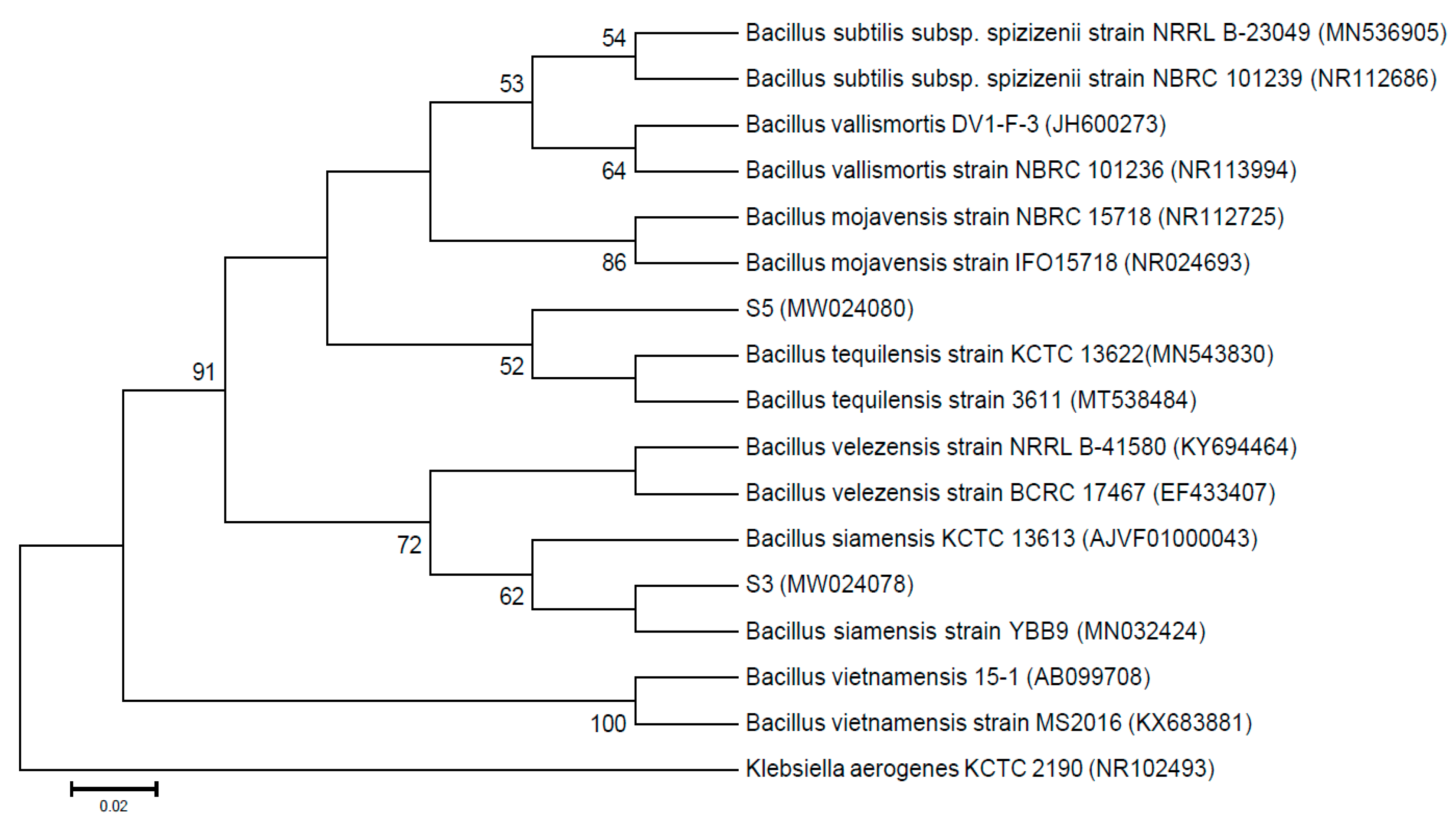
3.2. Antagonistic Bacteria were Identified as Bacillus
3.3. Extracellular Culture Filtrates Showed Antifungal Activity and Thermal Stability
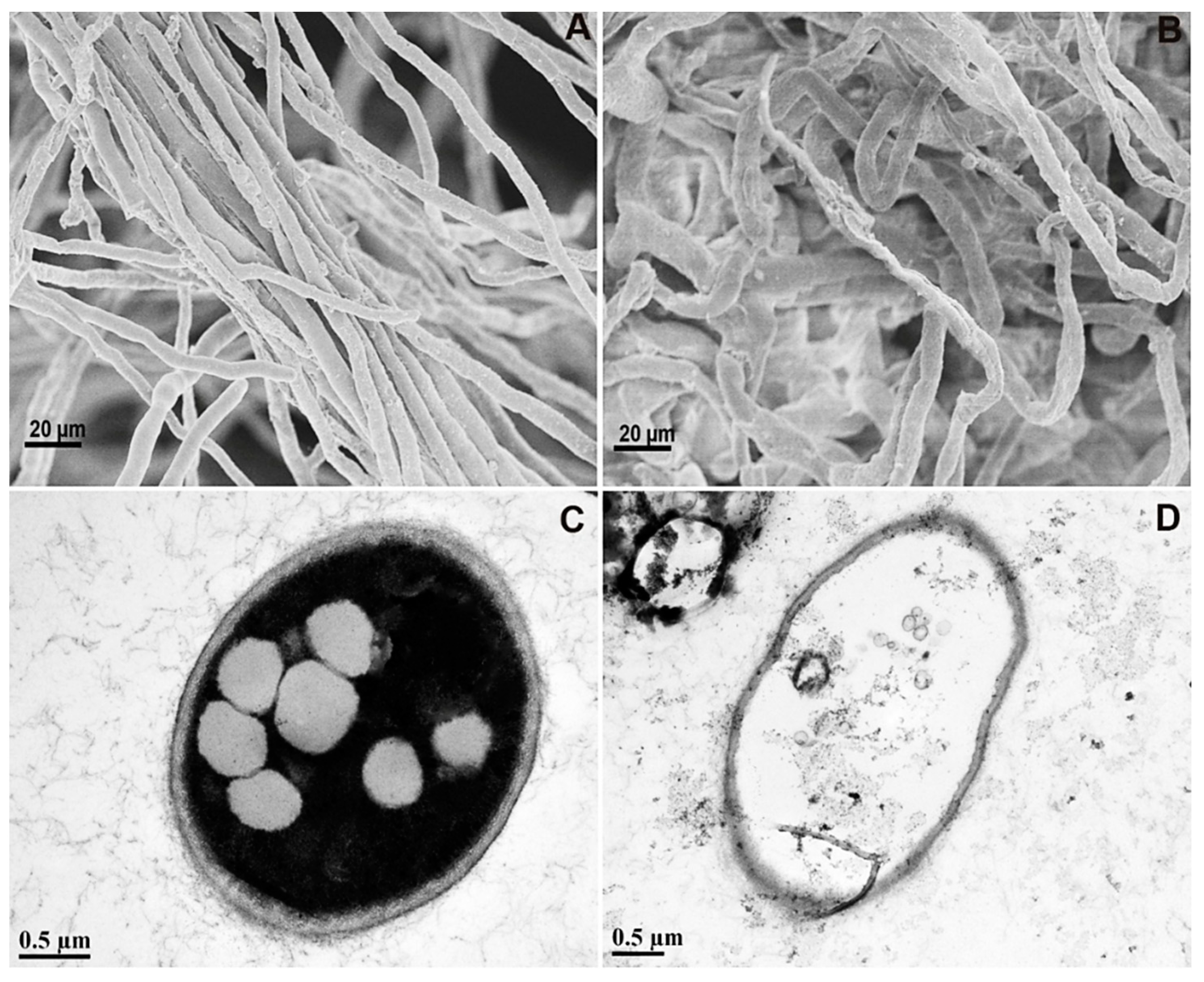
3.4. Damaged Hyphal Structures Were Observed by SEM and TEM
3.5. Hydrolytic Enzymes Activity Assay
3.6. Lipopeptides were Detected in S3 and S5
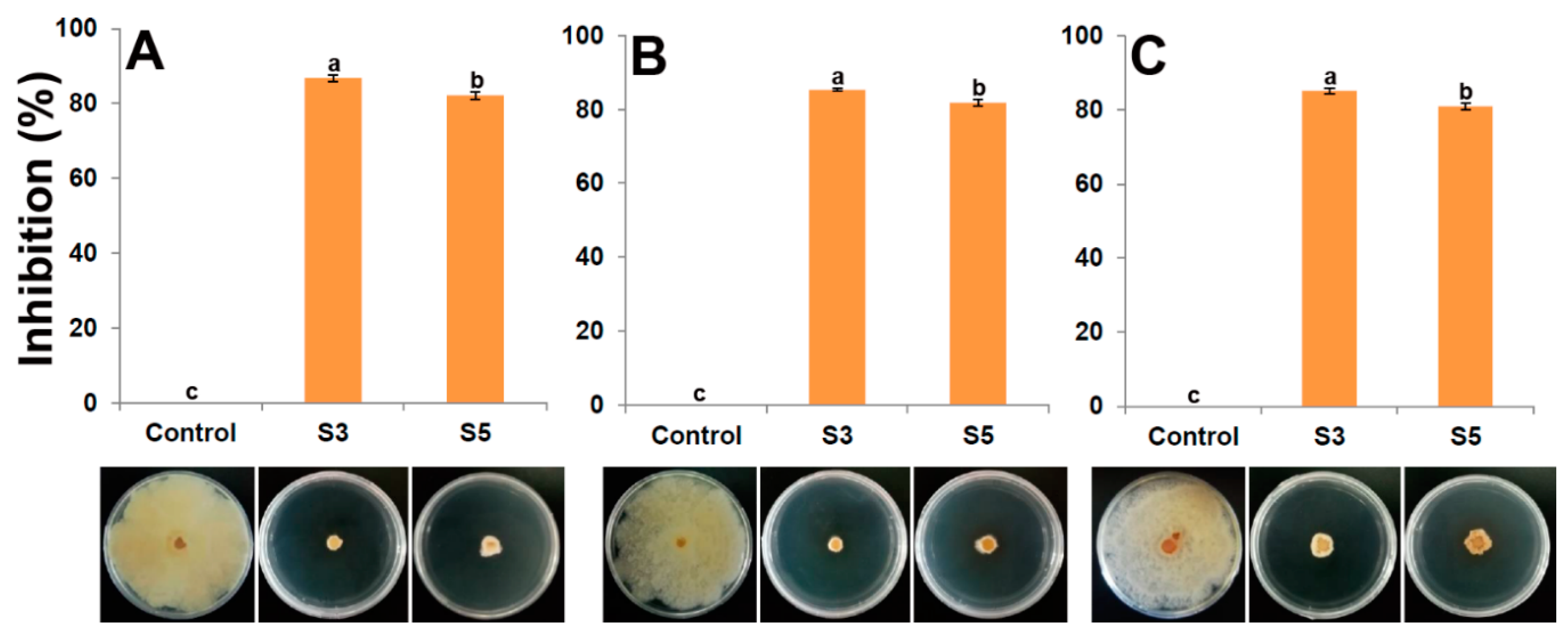
3.7. Inhibition of P. versicolor XJ27 on Bayberry Leaves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, K.; Xu, C.; Zhang, B.; Ferguson, I.B. Red bayberry: Botany and horticulture. Hortic. Rev. 2004, 30, 83–114. [Google Scholar]
- Shi, L.; Cao, S.; Chen, X.; Chen, W.; Zheng, Y.; Yang, Z. Proanthocyanidin Synthesis in Chinese Bayberry (Myrica rubra Sieb. Et Zucc.) Fruits. Front. Plant. Sci. 2018, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.L.; Zhang, S.L.; Chen, D.M. Red bayberry (Myrica rubra Seib & Zucc.): A valuable evergreen tree fruit for tropical and subtropical areas. Acta Hortic. 1992, 321, 112–121. [Google Scholar]
- Bao, J.; Cai, Y.; Sun, M.; Wang, G.; Corke, H. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem. 2005, 53, 2327–2332. [Google Scholar] [CrossRef]
- Fang, Z.; Zhang, M.; Tao, G.; Sun, Y.; Sun, J. Chemical Composition of Clarified Bayberry (Myrica rubra Sieb. Et Zucc.) Juice Sediment. J. Agric. Food Chem. 2006, 54, 7710–7716. [Google Scholar] [CrossRef]
- Chen, Y.F.; Lu, L.M.; Ni, H.Z.; Wang, Y.; Wang, Y.G.; Li, G.Q. First Report of Pestalotiopsis mangiferae and P. vismiae Causing Twig Dieback of Myrica rubra in China. Plant. Dis. 2012, 96, 588. [Google Scholar] [CrossRef]
- Ren, -Y.H.; Li, G.; Qi, X.-J.; Fang, L.; Wang, H.-R.; Wei, J.-G.; Zhong, S. Identification and characterization of Pestalotiopsis spp. causing twig blight disease of bayberry (Myrica rubra Sieb. & Zucc) in China. Eur. J. Plant. Pathol. 2013, 137, 451–461. [Google Scholar]
- Li, W.; Hu, M.; Xue, Y.; Li, Z.; Zhang, Y.; Zheng, D.; Lu, G.; Wang, J.; Zhou, J. Five Fungal Pathogens Are Responsible for Bayberry Twig Blight and Fungicides Were Screened for Disease Control. Microorganisms 2020, 8, 689. [Google Scholar] [CrossRef]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical Pesticides and Human Health: The Urgent Need for a New Concept in Agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef] [Green Version]
- Cazorla, M.F.; Romero, D.; Pérez-García, A.; Lugtenberg, B.J.; Vicente, A.; Bloemberg, G. Isolation and characterization of antagonistic Bacillus subtilis strains from the avocado rhizoplane displaying biocontrol activity. J. Appl. Microbiol. 2007, 103, 1950–1959. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yan, F.; Chai, Y.; Liu, H.; Kolter, R.; Losick, R.; Guo, J.H. Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environ. Microbiol. 2013, 15, 848–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seleim, M.A.A.; Saead, F.A.; Abd-El-Mon, K.M.H.; Abo-ELyous, K.A.M. Biological control of bacterial wilt of tomato by plant growth promoting rhizobacteria. Plant. Pathol. J. 2011, 10, 146–153. [Google Scholar] [CrossRef]
- Law, -F.J.W.; Ser, H.-L.; Khan, T.M.; Chuah, L.-H.; Pusparajah, P.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. The Potential of Streptomyces as Biocontrol Agents against the Rice Blast Fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Maymon, M.; Hirsch, A.M. Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms 2017, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Mishra, A.; Nautiyal, C.S. Paenibacillus lentimorbus B-30488r controls early blight disease in tomato by inducing host resistance associated gene expression and inhibiting Alternaria solani. Biol. Control 2012, 62, 65–74. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; García, J.M.; Pozo, M.J. Induced systemic resistance against Botrytis cinerea by Micromonospora strains isolated from root nodules. Front. Microbiol. 2015, 6, 922. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.; Doty, S.L. Characterization of bacterial endophytes of sweet potato plants. Plant. Soil 2009, 322, 197–207. [Google Scholar] [CrossRef]
- Marques, M.J.; da Silva, T.F.; Vollu, R.E.; Blank, A.F.; Ding, G.C.; Seldin, L.; Smalla, K. Plant age and genotype affect the bacterial community composition in the tuber rhizosphere of field-grown sweet potato plants. FEMS Microbiol. Ecol. 2014, 88, 424–435. [Google Scholar] [CrossRef]
- Marques, M.J.; da Silva, T.F.; Vollú, R.E.; de Lacerda, J.R.M.; Blank, A.F.; Smalla, K.; Seldin, L. Bacterial endophytes of sweet potato tuberous roots affected by the plant genotype and growth stage. Appl. Soil Ecol. 2015, 96, 273–281. [Google Scholar] [CrossRef]
- Puri, R.; Dangi, S.; Dhungana, S.; Itoh, K. Diversity and Plant Growth Promoting Ability of Culturable Endophytic Bacteria in Nepalese Sweet Potato. Adv. Microbiol. 2018, 8, 734–761. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Yang, X.; Lu, D.; Jin, H.; Yan, Z.; Chen, J.; Li, X.; Qin, B. Isolation and characterization of bacteria from the rhizosphere and bulk soil of Stellera chamaejasme L. Can. J. Microbiol. 2015, 61, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xu, Y.; Lai, X. Antagonistic endophytic bacteria associated with nodules of soybean (Glycine max L.) and plant growth-promoting properties. Braz. J. Microbiol. 2018, 49, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Mukta, J.A.; Rahman, M.; Sabir, A.A.; Gupta, D.R.; Surovy, M.Z.; Rahman, M.; Islam, M.T. Chitosan and plant probiotics application enhance growth and yield of strawberry. Biocatal. Agric. Biotechnol. 2017, 11, 9–18. [Google Scholar] [CrossRef]
- Li, B.; Xu, L.H.; Lou, M.M.; Li, F.; Zhang, Y.D.; Xie, G.L. Isolation and characterization of antagonistic bacteria against bacterial leaf spot of Euphorbia pulcherrima. Lett. Appl. Microbiol. 2008, 46, 450–455. [Google Scholar] [CrossRef]
- Altschul, F.S.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Ohike, T.; Makuni, K.; Okanami, M.; Ano, T. Screening of endophytic bacteria against fungal plant pathogens. J. Environ. Sci. 2013, 25, S122–S126. [Google Scholar] [CrossRef]
- Gao, T.; Zhou, H.; Zhou, W.; Hu, L.; Chen, J.; Shi, Z. The Fungicidal Activity of Thymol against Fusarium graminearum via Inducing Lipid Peroxidation and Disrupting Ergosterol Biosynthesis. Molecules 2016, 21, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krithika, S.; Chellaram, C. Isolation, Screening and Characterization of Chitinase producing Bacteriafrom marine wastes. Int. J. Pharm. Pharm. Sci. 2016, 8, 34–36. [Google Scholar]
- Oberai, M.; Khanna, V. Mung Bean Rhizobacteria Antagonist to Fusarium oxysporum and Rhizoctonia solani. Acta Sci. Microbiol. 2019, 2, 82–90. [Google Scholar] [CrossRef]
- Mahasneh, A.M.; Stewart, D.J. A Medium for detecting β-(1→3) Glucanase Activity in Bacteria. J. Appl. Bacteriol. 1980, 48, 457–458. [Google Scholar] [CrossRef]
- Masum, I.M.M.; Liu, L.; Yang, M.; Hossain, M.M.; Siddiqa, M.M.; Supty, M.E.; Ogunyemi, S.O.; Hossain, A.; An, Q.; Li, B. Halotolerant bacteria belonging to operational group Bacillus amyloliquefaciens in biocontrol of the rice brown stripe pathogen Acidovorax oryzae. J. Appl. Microbiol. 2018, 125, 1852–1867. [Google Scholar] [CrossRef]
- Vater, J.; Niu, B.; Dietel, K.; Borriss, R. Characterization of Novel Fusaricidins Produced by Paenibacillus polymyxa-M1 Using MALDI-TOF Ms Spectrometry. J. Am. Soc. Mass Spectrom. 2015, 26, 1548–1558. [Google Scholar] [CrossRef] [Green Version]
- Vater, J.; Herfort, S.; Doellinger, J.; Weydmann, M.; Dietel, K.; Faetke, S.; Lasch, P. Fusaricidins from PaenibacilluspolymyxaM-1, a family of lipohexapeptides of unusual complexity-a mass spectrometric study. J. Mass Spectrom. 2017, 52, 7–15. [Google Scholar] [CrossRef]
- Vater, J.; Kablitz, B.; Wilde, C.; Franke, P.; Mehta, N.; Cameotra, S.S. Matrix-assisted laser desorption ionization--time of flight mass spectrometry of lipopeptide biosurfactants in whole cells and culture filtrates of Bacillus subtilis C-1 isolated from petroleum sludge. Appl. Environ. Microbiol. 2002, 68, 6210–6219. [Google Scholar] [CrossRef] [Green Version]
- Vater, J.; Xuewen, G.; Gabriele, H.; Christopher, W.; Peter, F. “Whole Cell” Matrix-Assisted Laser Desorption Ionization-Time of Flight-Mass Spectrometry, an Emerging Technique for Efficient Screening of Biocombinatorial Libraries of Natural Compounds–Present State of Research. Comb. Chem. High. Throughput Screen. 2003, 6, 557–567. [Google Scholar] [CrossRef]
- Pueyo, T.M.; Mutafci, B.A.; Soto-Arriaza, M.A.; di Mascio, P.; Carmona-Ribeiro, A.M. The Self-Assembly of a Cyclic Lipopeptides Mixture Secreted by a B. megaterium Strain and Its Implications on Activity against a Sensitive Bacillus Species. PLoS ONE 2014, 9, e97261. [Google Scholar] [CrossRef] [Green Version]
- Hossain, A.; Masum, M.M.I.; Wu, X.; Abdallah, Y.; Ogunyemi, S.O.; Wang, Y.; Sun, G.; Li, B.; An, Q. Screening of Bacillus strains in biocontrol of pathogen Dickeya dadantii causing stem and root rot disease of sweet potato. Biocontrol Sci. Technol. 2020, 30. [Google Scholar] [CrossRef]
- Sarwar, A.; Hassan, M.N.; Imran, M.; Iqbal, M.; Majeed, S.; Brader, G.; Sessitsch, A.; Hafeez, F.Y. Biocontrol activity of surfactin A purified from Bacillus NH-100 and NH-217 against rice bakanae disease. Microbiol. Res. 2018, 209, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, L.; Su, C.X.; Gong, G.H.; Wang, P.; Yu, Z.L. Isolation and characterization of lipopeptide antibiotics produced by Bacillus subtilis. Lett. Appl. Microbiol. 2008, 47, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Perez, J.K.; Viana, J.D.; Lopes, F.C.; Pereira, J.Q.; Santos, D.M.D.; Oliveira, J.S.; Velho, R.V.; Crispim, S.M.; Nicoli, J.R.; Brandelli, A.; et al. Bacillus spp. Isolated from Puba as a Source of Biosurfactants and Antimicrobial Lipopeptides. Front. Microbiol. 2017, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Pathak, V.K.; Bose, A.; Keharia, H. Characterization of Novel Lipopeptides Produced by Bacillus tequilensis P15 Using Liquid Chromatography Coupled Electron Spray Ionization Tandem Mass Spectrometry (Lc ESI MS/MS). Int. J. Pept. Res. Ther. 2013, 20, 133–143. [Google Scholar] [CrossRef]
- Pathak, K.V.; Keharia, H. Identification of surfactins and iturins produced by potent fungal antagonist, Bacillus subtilis K1 isolated from aerial roots of banyan (Ficus benghalensis) tree using mass spectrometry. 3 Biotech 2014, 4, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Stein, T. Whole-cell matrix-assisted laser desorption/ionization mass spectrometry for rapid identification of bacteriocin/lantibiotic-producing bacteria. Rapid Commun. Mass Spectrom. 2008, 22, 1146–1152. [Google Scholar] [CrossRef]
- Wang, X.; Li, Q.; Sui, J.; Zhang, J.; Liu, Z.; Du, J.; Xu, R.; Zhou, Y.; Liu, X. Isolation and Characterization of Antagonistic Bacteria Paenibacillus jamilae HS-26 and Their Effects on Plant Growth. Biomed. Res. Int. 2019. [Google Scholar] [CrossRef] [Green Version]
- Wardle, A.D.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological Linkages Between Aboveground and Belowground Biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Collins, P.D.; Jacobsen, B.J. Optimizing a Bacillus subtilis isolate for biological control of sugar beet cercospora leaf spot. Biol. Control 2003, 26, 153–161. [Google Scholar] [CrossRef]
- Gardener, B.M.M. Ecology of Bacillus and Paenibacillus spp. in Agricultural Systems. Phytopathology 2004, 94, 1252–1258. [Google Scholar] [CrossRef] [Green Version]
- Raupach, S.G.; Kloepper, J.W. Mixtures of Plant Growth-Promoting Rhizobacteria Enhance Biological Control of Multiple Cucumber Pathogens. Phytopathology 1998, 88, 1158–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoda, M. Bacterial control of plant diseases. J. Biosci. Bioeng. 2000, 89, 515–521. [Google Scholar] [CrossRef]
- Romero, D.; Pérez-García, A.; Rivera, M.E.; Cazorla, F.M.; de Vicente, A. Isolation and evaluation of antagonistic bacteria towards the cucurbit powdery mildew fungus Podosphaerafusca. Appl. Microbiol. Biotechnol. 2004, l64, 263–269. [Google Scholar] [CrossRef]
- Raaijmakers, M.J.; Vlami, M.; de Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Leeuwenhoek 2002, 81, 37. [Google Scholar] [CrossRef]
- Schisler, A.D.; Slininger, P.J.; Behle, R.W.; Jackson, M.A. Formulation of Bacillus spp. for Biological Control of Plant Diseases. Phytopathology 2004, 94, 1267–1271. [Google Scholar] [CrossRef] [Green Version]
- Ongena, M.; Jacques, P.; Touré, Y.; Destain, J.; Jabrane, A.; Thonart, P. Involvement of fengycin-type lipopeptides in the multifaceted biocontrol potential of Bacillus subtilis. Appl. Microbiol. Biotechnol. 2005, 69, 29–38. [Google Scholar] [CrossRef]
- Sellem, I.; Triki, M.A.; Elleuch, L.; Cheffi, M.; Chakchouk, A.; Smaoui, S.; Mellouli, L. The use of newly isolated Streptomyces strain TN258 as potential biocontrol agent of potato tubers leak caused by Pythium ultimum. J. Basic Microbiol. 2017, 57, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 2004, 150, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Sultana, R.; Chae, J.-C.; Kim, K.; Lee, K.-J. Bacillus thuringiensis C25 which is rich in cell wall degrading enzymes efficiently controls lettuce drop caused by Sclerotinia minor. Eur. J. Plant. Pathol. 2015, 142, 577–589. [Google Scholar] [CrossRef]
- Khan, N.; Martínez-Hidalgo, P.; Ice, T.A.; Maymon, M.; Humm, E.A.; Nejat, N.; Sanders, E.R.; Kaplan, D.; Hirsch, A.M. Antifungal Activity of Bacillus Species against Fusarium and Analysis of the Potential Mechanisms Used in Biocontrol. Front. Microbiol. 2018, 9, 2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Guan, Y.; Dong, Y.; Zhao, L.; Rong, S.; Chen, W.; Lv, M.; Xu, H.; Gao, X.; Chen, R.; et al. Isolation and evaluation of endophytic Bacillus tequilensis GYLH001 with potential application for biological control of Magnaporthe oryzae. PLoS ONE 2018, 13, e0203505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazarika, J.D.; Goswami, G.; Gautom, T.; Parveen, A.; Das, P.; Barooah, M.; Boro, R.C. Lipopeptide mediated biocontrol activity of endophytic Bacillus subtilis against fungal phytopathogens. BMC Microbiol. 2019, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Asaka, O.; Shoda, M. Biocontrol of Rhizoctonia solani Damping-Off of Tomato with Bacillus subtilis RB14. Appl. Environ. Microbiol. 1996, 62, 4081–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koumoutsi, A.; Chen, X.-H.; Henne, A.; Liesegang, H.; Hitzeroth, G.; Franke, P.; Vater, J.; Borriss, R. Structural and functional characterization of gene clusters directing nonribosomal synthesis of bioactive cyclic lipopeptides in Bacillus amyloliquefaciens strain FZB42. J. Bacteriol. 2004, 186, 1084–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, D.; de Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.W.; Arrebola, E.; Cazorla, F.M.; Kuipers, O.P.; Paquot, M.; Pérez-García, A. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol. Plant. Microbe Interact. 2007, 20, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Gao, L.; Bie, X.; Lu, Z.; Liu, H.; Zhang, C.; Lu, F.; Zhao, H. Identification of novel surfactin derivatives from NRPS modification of Bacillus subtilis and its antifungal activity against Fusarium moniliforme. BMC Microbiol. 2016, 16, 31. [Google Scholar] [CrossRef] [Green Version]
- Gong, -D.A.; Li, H.-P.; Yuan, Q.-S.; Song, X.-S.; Yao, W.; He, W.-J.; Zhang, J.-B.; Liao, Y.-C. Antagonistic Mechanism of Iturin A and Plipastatin A from Bacillus amyloliquefaciens S76-3 from Wheat Spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar]
- Leclère, V.; Béchet, M.; Adam, A.; Guez, J.-S.; Wathelet, B.; Ongena, M.; Thonart, P.; Gancel, F.; Chollet-Imbert, M.; Jacques, P. Mycosubtilin Overproduction by Bacillus subtilis BBG100 Enhances the Organisms Antagonistic and Biocontrol Activities. Appl. Environ. Microbiol. 2005, 71, 4577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munimbazi, C.; Bullerman, L.B. Isolation and partial characterization of antifungal metabolites of Bacillus pumilus. J. Appl. Microbiol. 1998, 84, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Nagórska, K.; Bikowski, M.; Obuchowski, M. Multicellular behaviour and production of a wide variety of toxic substances support usage of Bacillus subtilis as a powerful biocontrol agent. Acta Biochim. Pol. 2007, 54, 495–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed Mass Peaks (m/z) | Lipopeptides | References |
|---|---|---|
| Strain: S3 | ||
| 1023.627 | Leu/Ile-7 C15 Surfactin, [M + H]+ | [41] |
| 1039.599 | C15 Surfactin, [M + Na − H2O]+ | [42] |
| 1058.783 | C15 Surfactin, [M + Na]+ | [43] |
| 1065.658 | C14 Iturin, [M + Na]+ | [43] |
| 1074.755 | C15 Surfactin, [M + K]+ | [44] |
| 1079.677 | C15 Iturin, [M + Na]+ | [43] |
| 1095.657 | C15 Iturin, [M + K]+ | [43] |
| 1100.731 | C18 Iturin, [M + H]+ | [45] |
| 1109.679 | C16 Iturin, [M + K]+ | [46] |
| 1122.707 | C17 Iturin, [M + K]+ | [38] |
| 1138.679 | C18 Mycosubtilin, [M + K]+ | [47] |
| 1151.747 | C19 Mycosubtilin, [M + K]+ | [47] |
| 1257.823 | Polymyxin B6, [M + K]+ | [41] |
| Strain: S5 | ||
| 1030.805 | C13 Surfactin, [M + Na]+ | [43] |
| 1044.823 | C14 Surfactin, [M + Na]+ | [43] |
| 1058.845 | C15 Surfactin, [M + Na]+ | [43] |
| 1074.816 | C15 Surfactin, [M + K]+ | [44] |
| 1100.785 | C18 Iturin, [M + H]+ | [45] |
| 1122.808 | C17 Iturin, [M + K]+ | [38] |
| 1151.807 | C19 Mycosubtilin [M + K]+ | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.A.; Ren, H.; Ahmed, T.; Luo, J.; An, Q.; Qi, X.; Li, B. Antifungal Effects of Rhizospheric Bacillus Species Against Bayberry Twig Blight Pathogen Pestalotiopsis versicolor. Agronomy 2020, 10, 1811. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10111811
Ali MA, Ren H, Ahmed T, Luo J, An Q, Qi X, Li B. Antifungal Effects of Rhizospheric Bacillus Species Against Bayberry Twig Blight Pathogen Pestalotiopsis versicolor. Agronomy. 2020; 10(11):1811. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10111811
Chicago/Turabian StyleAli, Md. Arshad, Haiying Ren, Temoor Ahmed, Jinyan Luo, Qianli An, Xingjiang Qi, and Bin Li. 2020. "Antifungal Effects of Rhizospheric Bacillus Species Against Bayberry Twig Blight Pathogen Pestalotiopsis versicolor" Agronomy 10, no. 11: 1811. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10111811