Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Treatments, and Growth Conditions
2.2. Measurements
2.2.1. Growth and Yield Attributes
2.2.2. Anatomical Characteristics
2.2.3. Leaf Pigments
2.2.4. Total Soluble Sugar and Proline
2.2.5. Total Protein
2.2.6. Total Alkaloid
2.3. Statistical Analysis
3. Results
3.1. Growth Characteristics
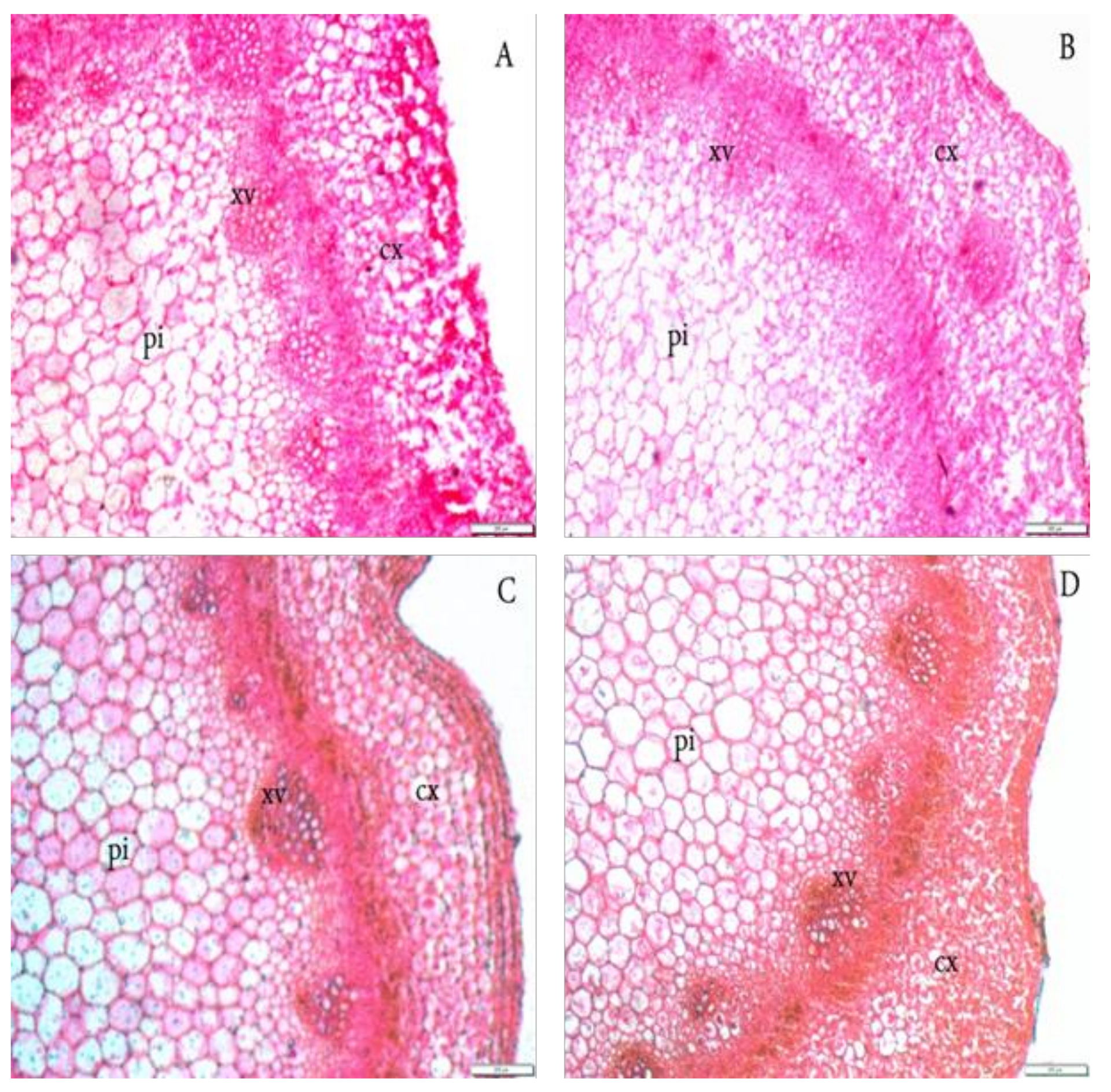
3.2. Anatomical Structure
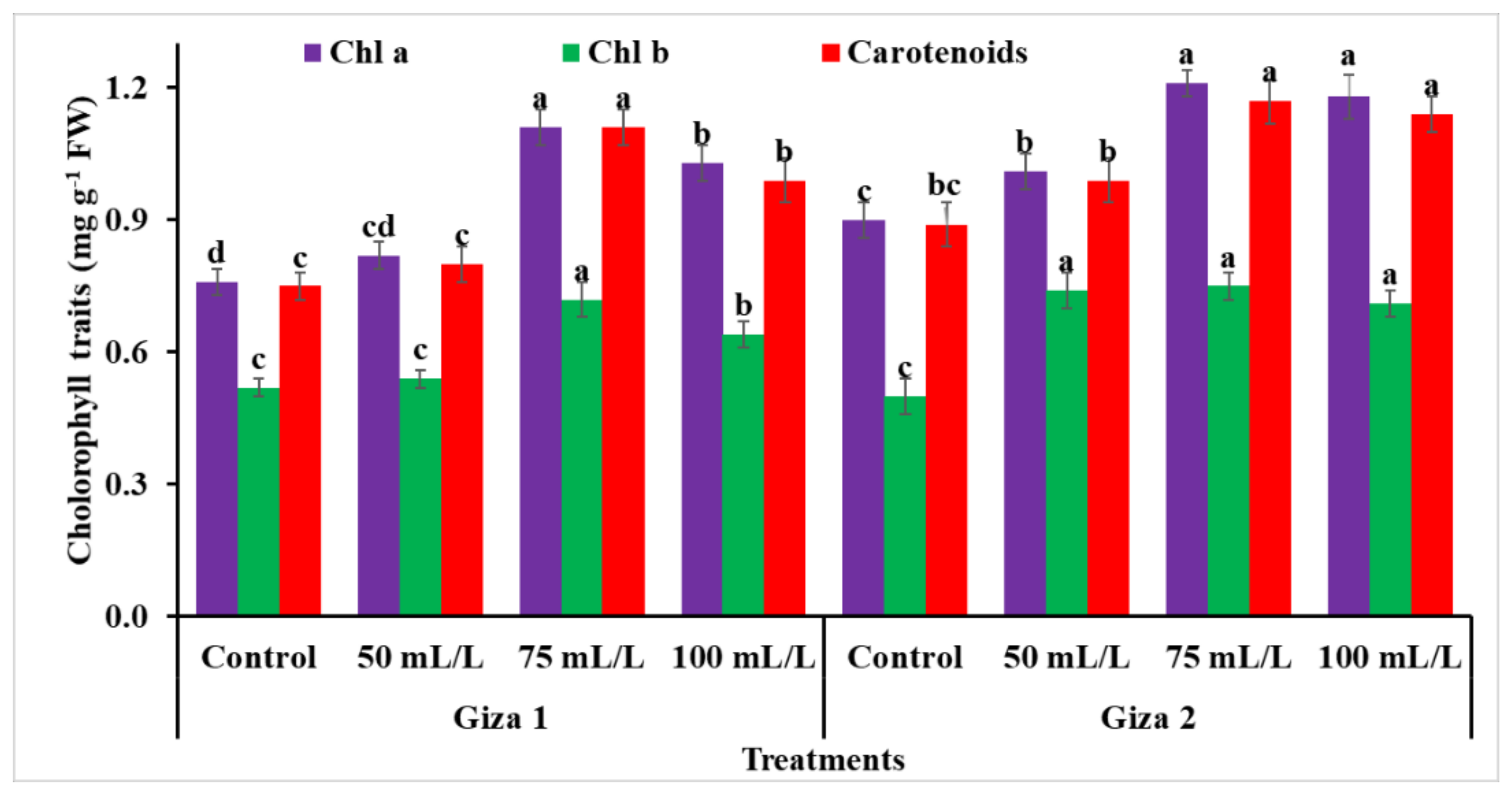
3.3. Chlorophyll a, b and Carotenoids
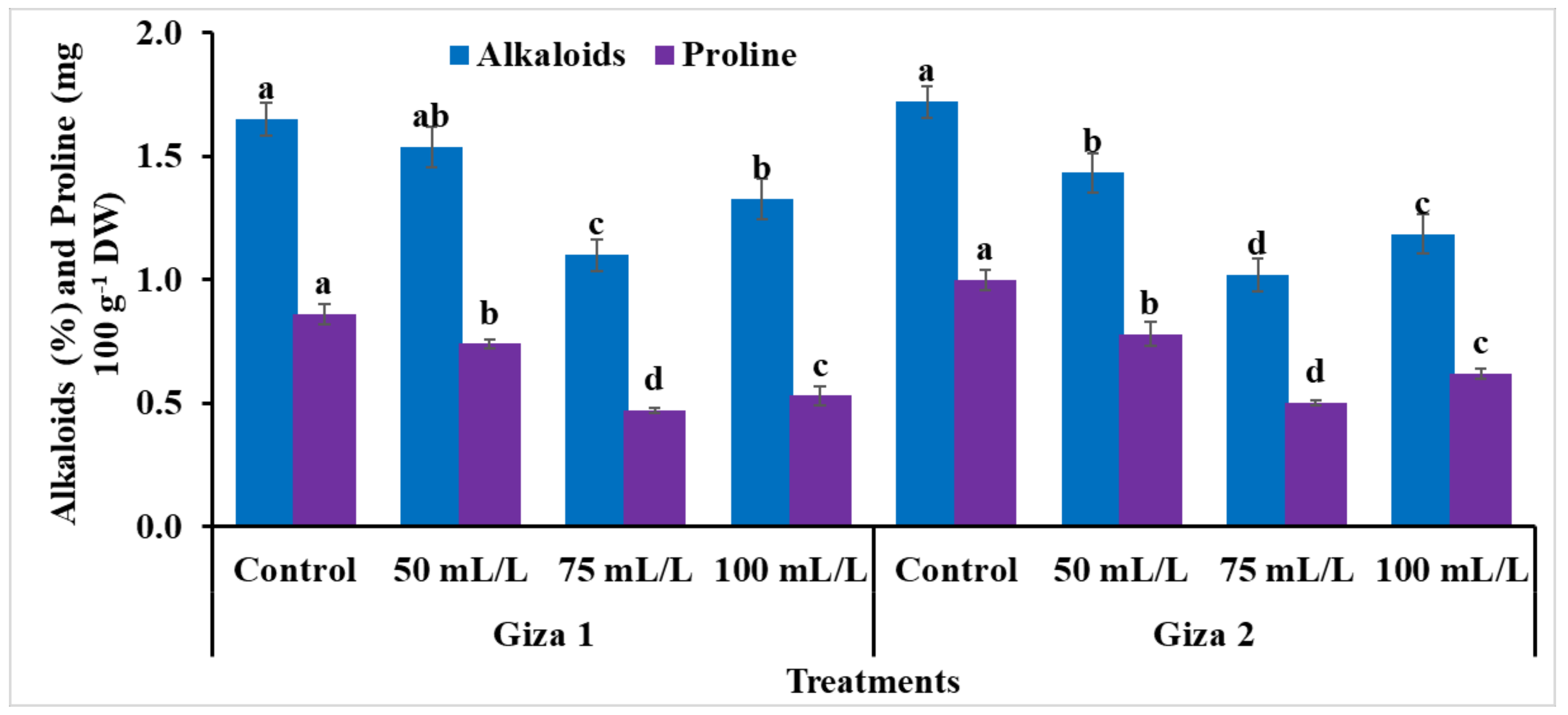
3.4. Total Protein, Total Soluble Sugar (TSS), Alkaloids, and Proline
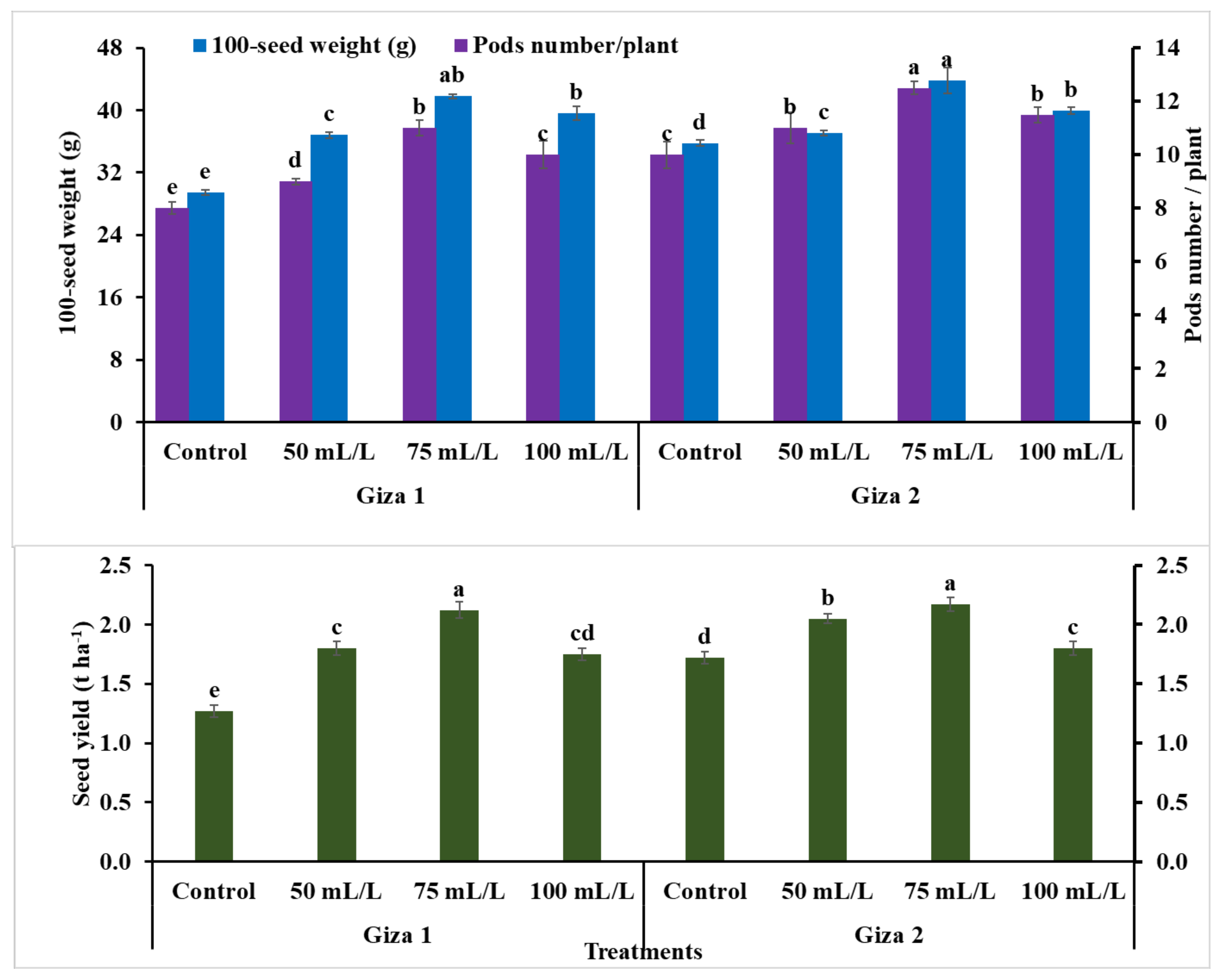
3.5. Yield and Its Components
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seleiman, M.F.; Santanen, A.; Stoddard, F.L.; Mäkelä, P.S.A. Feedstock Quality and Growth of Bioenergy Crops Fertilized with Sewage Sludge. Chemosphere 2012, 89, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Santanen, A.; Kleemola, J.; Stoddard, F.L.; Mäkelä, P.S.A. Improved Sustainability of Feedstock Production with Sludge and Interacting Mychorriza. Chemosphere 2013, 91, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Kheir, A.M.S. Maize Productivity, Heavy Metals Uptake and Their Availability in Contaminated Clay and Sandy Alkaline Soils as Affected by Inorganic and Organic Amendments. Chemosphere 2018, 204, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Kheir, A.M.S. Saline Soil Properties, Quality and Productivity of Wheat Grown with Bagasse Ash and Thiourea in Different Climatic Zones. Chemosphere 2018, 193, 538–546. [Google Scholar] [CrossRef]
- Seleiman, M.F. Use of Plant Nutrients in Improving Abiotic Stress Tolerance in Wheat. In Wheat Production in Changing Environments: Management, Adaptation and Tolerance; Hasanuzzaman, M., Nahar, K., Hossain, A., Eds.; Springer Nature: Singapore, 2019; pp. 481–495. ISBN 978-981-13-6883-7. [Google Scholar]
- Seleiman, M.F.; Kheir, A.M.S.; Al-Dhumri, S.; Alghamdi, A.G.; Omar, E.-S.H.; Aboelsoud, H.M.; Abdella, K.A.; Abou El Hassan, W.H. 2019. Exploring optimal tillage improved soil characteristics and productivity of wheat irrigated with different water qualities. Agronomy 2019, 9, 233. [Google Scholar] [CrossRef] [Green Version]
- Seleiman, M.F.; Alotaibi, M.A.; Alhammad, B.A.; Alharbi, B.M.; Refay, Y.; Badawy, S.A. Effects of ZnO Nanoparticles and Biochar of Rice Straw and Cow Manure on Characteristics of Contaminated Soil and Sunflower Productivity, Oil Quality, and Heavy Metals Uptake. Agronomy 2020, 10, 790. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Mäkelä, P.S.A. Recycling Sludge on Cropland as Fertilizer—Advantages and Risks. Resour. Conserv. Recycl. 2020, 155, 104647. [Google Scholar] [CrossRef]
- Taha, R.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; Mahdi, A.H.A. Exogenous Potassium Treatments Elevate Salt Tolerance and Performances of Glycine max L. by Boosting Antioxidant Defense System Under Actual Saline Field Conditions. Agronomy 2020, 10, 1741. [Google Scholar] [CrossRef]
- Dawood, M.G.; Taie, H.A.A.; Nassar, R.M.A.; Abdelhamid, M.T.; Schmidhalter, U. The Changes Induced in the Physiological, Biochemical and Anatomical Characteristics of Vicia Faba by The Exogenous Application of Proline Under Seawater Stress. S. Afr. J. Bot. 2014, 93, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; Taha, R.S.; Mahdi, A.H.A. Proline Enhances Growth, Productivity and Anatomy of Two Cultivars of Lupinus termis L. Grown Under Salt Stress. S. Afr. J. Bot. 2016, 102, 221–227. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; El-Hendawy, S.; Al-Suhaibani, N.; El-Kafafi, S.; Seleiman, M.F. Detecting Salt Tolerance in Doubled Haploid Wheat Lines. Agronomy 2019, 9, 211. [Google Scholar] [CrossRef] [Green Version]
- Al-Ashkar, I.; Alderfasi, A.; Ben Romdhane, W.; Seleiman, M.F.; El-Said, R.A.; Al-Doss, A. Morphological and Genetic Diversity within Salt Tolerance Detection in Eighteen Wheat Genotypes. Plants 2020, 9, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Z.; Kheir, A.S.; Ali, O.A.; Hafez, E.; Elshamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.; Ge, Y.; Fahmy, A.E.; et al. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2020, 277, 111388. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.S. Improving Salt Tolerance of Helianthus annuus, (L.) Plants by Moringa oleifera Leaf Extract. Egypt. J. Agron. 2016, 38, 117–140. [Google Scholar]
- Rady, M.M.; Taha, R.S.; Semida, W.M.; Alharby, H.F. Modulation of Salt Stress Effects on Vicia faba L. Plants Grown on a Reclaimed-Saline Soil by Salicylic Acid Application. Rom. Agric. Res. 2017, 34, 175–185. [Google Scholar]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; Ali, H.M.; Elshikh, M.S.; Abdel-Salam, E.M.; El-Esawi, M.; El-Ansary, D.O. Bioactivities of Traditional Medicinal Plants in Alexandria. Evid. Based Complement. Altern. Med. 2018, 2018, 1463579. [Google Scholar] [CrossRef] [Green Version]
- Al Mahmud, J.; Bhuyan, M.B.; Anee, T.I.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Reactive Oxygen Species Metabolism and Antioxidant Defense in Plants Under Metal/Metalloid Stress. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, Switzerland, 2019; pp. 221–257. [Google Scholar]
- Abdelaal, K.A.; El-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Subbarao, G.V.; Johansen, C.J. Potential for genetic improvement in salinity tolerance in legumes. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA, 1993; pp. 581–591. [Google Scholar]
- Seleiman, M.F.; Santanen, A.; Stoddard, F.L.; Mäkelä, P. Maize and white lupin as bioenergy crops grown in soil amended with sewage sludge. In Proceedings of the ‘Agro2010, the XIth ESA Congress’, Montpellier, France, 29 August–3 September 2010; Wery, J., Shili-Touzi, I., Perrin, A., Eds.; Agropolis International Editions: Montpellier, France; pp. 207–208. [Google Scholar]
- FAOSTAT—Food and Agriculture Organization of the United Nations Statistics Division. 2020. Available online: http://faostat.fao.org/site/567/DesktopDefault.aspx (accessed on 20 November 2020).
- Akladious, S.A.; Hanafy, R.S. lleviation of oxidative effects of salt stress in white lupine, (Lupinus termis L.) plants by foliar treatment with L-arginine. J. Anim. Plant. Sci. 2018, 28, 165–176. [Google Scholar]
- Barnett, J.A.; Payne, R.W.; Yarrow, D. Yeasts: Characteristics and Identification, 3rd ed.; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Abdelaal, K.A.; Hafez, Y.M.; El Sabagh, A.; Saneoka, H. Ameliorative Effects of Abscisic Acid and Yeast on Morpho-Physiological and Yield Characteristics of Maize Plant, (Zea mays L.) Under Water Deficit Conditions. Fresen. Environ. Bull. 2017, 26, 7372–7383. [Google Scholar]
- Xi, Q.; Lai, W.; Cui, Y.; Wu, H.; Zhao, T. Effect of Yeast Extract on Seedling Growth Promotion and Soil Improvement in Afforestation in a Semiarid Chestnut Soil Area. Forests 2019, 10, 76. [Google Scholar] [CrossRef] [Green Version]
- Kasim, W.A.E.A.; AboKassem, E.M.; Ragab, G.A.A. Ameliorative Effect of Yeast Extract, IAA and Green-Synthesized Nano Zinc Oxide on the Growth of Cu-Stressed Vicia faba Seedlings. Egypt. J. Bot. 2017, 57, 1–10. [Google Scholar] [CrossRef]
- El-Desouky, S.A.; Wanas, A.L.A.; Khedr, Z.M.A. Utilization of Some Natural Plant Extracts, (Garlic and Yeast) as Seed-Soaked Materials to Squash, (Cucurbita pepo, L.). Effect on Growth, Sex Expression and Fruit Yield and Quality. Ann. Agric. Sci. Moshtohor 1998, 36, 839–854. [Google Scholar]
- Wanas, A.L. Resonance of Faba Bean, (Vicia faba L.) Plants to Seed Soaking Application with Natural Yeast and Carrot Extracts. Ann. Agric. Sci. Moshtohor 2002, 40, 259–278. [Google Scholar]
- Wanas, A.L. Trails for Improving Growth and Productivity of Tomato Plants Grown in Winter. Ann. Agric. Sci. Moshtohor 2006, 44, 466–471. [Google Scholar]
- Klute, A. Methods of Soil Analysis: Part. 1—Physical and Mineralogical Methods, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1986. [Google Scholar]
- Page, A.I.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis: Part. 2—Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Nassar, M.A.; El-Sahhar, K.F. Botanical Preparations and Microscopy, (Microtechnique); Academic Bookshop: Dokki, Giza, Egypt, 1998; p. 219. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Irigoyen, J.J.; Emerich, D.W.; Sanchez-Diaz, M. Water Stress Induced Changes in the Concentrations of Proline and Total Soluble Sugars in Nodulated Alfalfa, (Medicago sativa) Plants. Plant. Physiol. 1992, 8, 455–460. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant. Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-Total. Methods of Soil Analysis: Part 2—Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1992; pp. 595–624. [Google Scholar]
- Misra, N.; Gupta, A.K. Effect of Salinity and Different Nitrogen Sources on the Activity of Antioxidant Enzymes and Indole Alkaloid Content in Catharanthus roseus Seedlings. J. Plant. Physiol. 2006, 163, 11–18. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: Singapore, 1984; p. 680. [Google Scholar]
- Farsiani, A.; Ghobadi, M.E. Effects of PEG and NaCl Stress on Two Cultivars of Corn, (Zea mays L.) at Germination and Early Seedling Stages. World Acad. Sci. Eng. Technol. 2009, 57, 382–385. [Google Scholar]
- Wu, D.; Qiu, L.; Xu, L.; Ye, L.; Chen, M.; Sun, D.; Chen, Z.; Zhang, H.; Jin, X.; Dai, F.; et al. Genetic Variation of HvCBF Genes and Their Association with Salinity Tolerance in Tibetan Annual Wild Barley. PLoS ONE 2011, 6, e22938. [Google Scholar] [CrossRef] [Green Version]
- Mansour, E.; Moustafa, E.S.A.; Desoky, E.-S.M.; Ali, M.M.A.; Yasin, M.A.T.; Attia, A.; Alsuhaibani, N.; Tahir, M.U.; El-Hendawy, S. Multidimensional Evaluation for Detecting Salt Tolerance of Bread Wheat Genotypes Under Actual Saline Field Growing Conditions. Plants 2020, 9, 1324. [Google Scholar] [CrossRef]
- Medani, R.A.; Taha, R.S. Improving Growth and Yield of Caraway, (Carum carvi L.). Plants by Decapitation and/or Active Dry Yeast Application. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 47–60. [Google Scholar]
- Sarhan, T.; Abdullah, O.K. Effect of Azotobacter Inoculation, Dry Bread Yeast Suspension and Varying Levels of Urea on Growth of Potato Cv. Desiree. In Proceedings of the World Food System—A Contribution from Europe, Tropentag, Zurich, Germany, 14–16 September 2010. [Google Scholar]
- Semida, W.M.; Taha, R.S.; Abdelhamid, M.T.; Rady, M.M. Foliar-Applied ɑ-Tocopherol Enhances Salt-Tolerance in Vicia faba L. Plants Grown Under Saline Conditions. S. Afr. J. Bot. 2014, 95, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Jaleel, C.A.; Sankar, B.; Sriaharan, R.; Panneerselvam, R. Soil Salinity Alters Growth, Chlorophyll Content, and Secondary Metabolite Accumulation in Catharanthus roseus. Turk. J. Biol. 2008, 32, 79–83. [Google Scholar]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of Drought Stress on Yield, Proline and Chlorophyll Contents in Three Chickpea Cultivars. Aust. J. Crop. Sci. 2009, 48, 580–585. [Google Scholar]
- Amprayn, K.O.; Rose, M.T.; Kecskés, M.; Pereg, L.; Nguyen, H.T.; Kennedy, I.R. Plant Growth Promoting Characteristics of Soil Yeast, (Candida tropicalis HY) and Its Effectiveness for Promoting Rice Growth. Appl. Soil Ecol. 2012, 61, 295–299. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Alías-Villegas, C.; Bellogín, R.A.; Del Cerro, P.; Espuny, M.R.; Jiménez-Guerrero, I.; López-Baena, F.J.; Ollero, F.J.; Cubo, T. Plant Growth Promotion in Cereal and Leguminous Agricultural Important Plants: From Microorganism Capacities to Crop Production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Foyer, C.H.; Harbinson, J.; Mullineaux, P. Oxygen Metabolism and the Regulation of Photosynthetic Electron Transport. In Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants; CRC Press: New York, NY, USA, 1994; pp. 1–42. [Google Scholar]
- Le Mire, G.; Nguyen, M.L.; Fassotte, B.; Du Jardin, P.; Verheggen, F.; Delaplace, P.; Jijakli, M.H. Implementing Plant Biostimulants and Biocontrol Strategies in the Agroecological Management of Cultivated Ecosystems. A Review. Biotechnol. Agron. Soc. Environ. 2016, 20, 299–313. [Google Scholar]
- Rajendrakumar, C.S.V.; Suryanarayana, T.; Reddy, A.R. DNA Helix Destabilization by Proline and Betaine: Possible Role in the Salinity Tolerance Process. FEBS Lett. 1997, 410, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N.C.; Cumbes, Q.J. Hydroxyl Radical Scavenging Activity of Compatible Solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops Under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Lei, P.; Xu, Z.; Liang, J.; Luo, X.; Zhang, Y.; Feng, X.; Xu, H. Poly, (g-glutamic acid) enhanced tolerance to salt stress by promoting proline accumulation in Brassica napus L. Plant. Growth Regul. 2016, 78, 233–241. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Evaluation of proline functions in saline conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, P.N.; Amrutha, R.N.; Sangam, S.; Verma, D.P.; Kishor, P.B. Biochemical characterization, homology modeling and docking studies of ornithine deltaaminotransferase—An important enzyme in proline biosynthesis of plants. J. Mol. Graph. Model. 2007, 26, 709–719. [Google Scholar] [CrossRef]
- Emam, M.M. Efficiency of Yeast in Enhancement of the Oxidative Defense System in Salt-Stressed Flax Seedlings. Acta Biol. Hung. 2013, 64, 118–130. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Bassuony, F.M.; Baraka, D.M.; Khalil, R.R. Physiological Effects of Nicotinamide and Ascorbic Acid on Zea mays Plant Grown Under Salinity Stress. 1-Changes in Growth, Some Relevant Metabolic Activities and Oxidative Defense Systems. Res. J. Agric. Biol. Sci. 2009, 5, 72–81. [Google Scholar]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioe Ectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 41, 5. [Google Scholar] [CrossRef] [Green Version]
- Sarhan, T.Z.; Ali, S.T.; Rasheed, S.M.S. Effect of Bread Yeast Application and Seaweed Extract on Cucumber, (Cucumis sativus L.) Plant Growth, Yield and Fruit Quality. Mesopotamia J. Agric. 2011, 39, 26–34. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macro-Elements (%) | Value | Micro-Elements (%) | Value |
|---|---|---|---|
| N | 1.00 | Fe | 0.11 |
| P | 0.11 | Mn | 0.06 |
| K | 0.3 | Zn | 0.05 |
| Mg | 0.011 | Cu | 0.03 |
| Ca | 0.02 | B | 0.014 |
| Na | 0.009 | Mo | 0.0002 |
| Total protein (%) | 5.00 | Phyto-hormones (mg mL−1) | |
| Total carbohydrate (%) | 4.5 | Indole-3-acetic acid | 0.6 |
| Gibberellins | 0.3 | ||
| 2017/2018 | 2018/2019 | |
|---|---|---|
| Physical properties | ||
| Clay (%) | 13.2 | 12.1 |
| Silt (%) | 17.0 | 16.5 |
| Sand (%) | 69.8 | 71.4 |
| Soil texture | Sandy loam | Sandy loam |
| Chemical properties | ||
| pH (1:2.5) | 7.40 | 7.70 |
| EC (dS m−1) | 7.55 | 7.75 |
| Organic matter (%) | 0.90 | 0.95 |
| CaCO3 (%) | 4.78 | 4.95 |
| Total N (%) | 0.062 | 0.069 |
| Available P (mg kg−1 soil) | 7.56 | 8.00 |
| Available K (mg kg−1 soil) | 176 | 179 |
| Available Fe (mg kg−1 soil) | 6.22 | 6.28 |
| Treatment | Section Diameter (μ) | Cortex Thickness (μ) | No. of Cortical Layers | Xylem Vessels Diameter (μ) | Pith Diameter (μ) | No. of Pith Layers | |
|---|---|---|---|---|---|---|---|
| Variety | AYE | ||||||
| (mL L−1) | |||||||
| Giza 1 | 0 | 3775.0 e | 300.0 c | 6.0 d | 22.5 c | 2525.0 d | 32.5 cd |
| 50 | 4562.5 c | 350.0 b | 7.0 c | 30.0 b | 2662.5 d | 31.5 d | |
| 75 | 5225.0 a | 400.0 a | 9.0 a | 40.0 a | 3687.5 b | 38.0 a | |
| 100 | 5000.0 b | 400.0 a | 8.0 b | 30.0 b | 3375.0 c | 37.5 a | |
| Means | 4640.63 | 337.50 | 7.5 | 30.63 | 3062.50 | 34.88 | |
| Giza 2 | 0 | 4062.5 d | 240.0 d | 7.0 c | 17.5 d | 2337.5 a | 33.5 c |
| 50 | 5062.5 b | 400.0 a | 8.0 b | 30.0 b | 3312.5 c | 36.0 b | |
| 75 | 5250.0 a | 400.0 a | 9.0 a | 40.0 a | 4062.5 a | 38.5 a | |
| 100 | 5187.5 a | 400.0 a | 8.0 b | 40.0 a | 3437.5 c | 35.0 b | |
| Means | 4890.63 | 360.00 | 8.00 | 31.88 | 3287.50 | 35.75 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
S. Taha, R.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H.A. Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy 2021, 11, 74. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010074
S. Taha R, Seleiman MF, Alhammad BA, Alkahtani J, Alwahibi MS, Mahdi AHA. Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy. 2021; 11(1):74. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010074
Chicago/Turabian StyleS. Taha, Ragab, Mahmoud F. Seleiman, Bushra Ahmed Alhammad, Jawaher Alkahtani, Mona S. Alwahibi, and Ayman H. A. Mahdi. 2021. "Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions" Agronomy 11, no. 1: 74. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010074