
Disclosing the Nutritional Quality Diversity of Portuguese Common Beans—The Missing Link for Their Effective Use in Protein Quality Breeding Programs

 ,
,  , ,
, ,  , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Sample Preparation
2.4. General Nutritional Composition
2.4.1. Total Protein, Fat, Fiber, Moisture and Ash Content
2.4.2. Calculated Total Carbohydrate Content
2.5. Protein Quality
2.5.1. Amino Acids’ Extraction
2.5.2. Amino Acid Content
2.5.3. Trypsin Inhibitor Activity
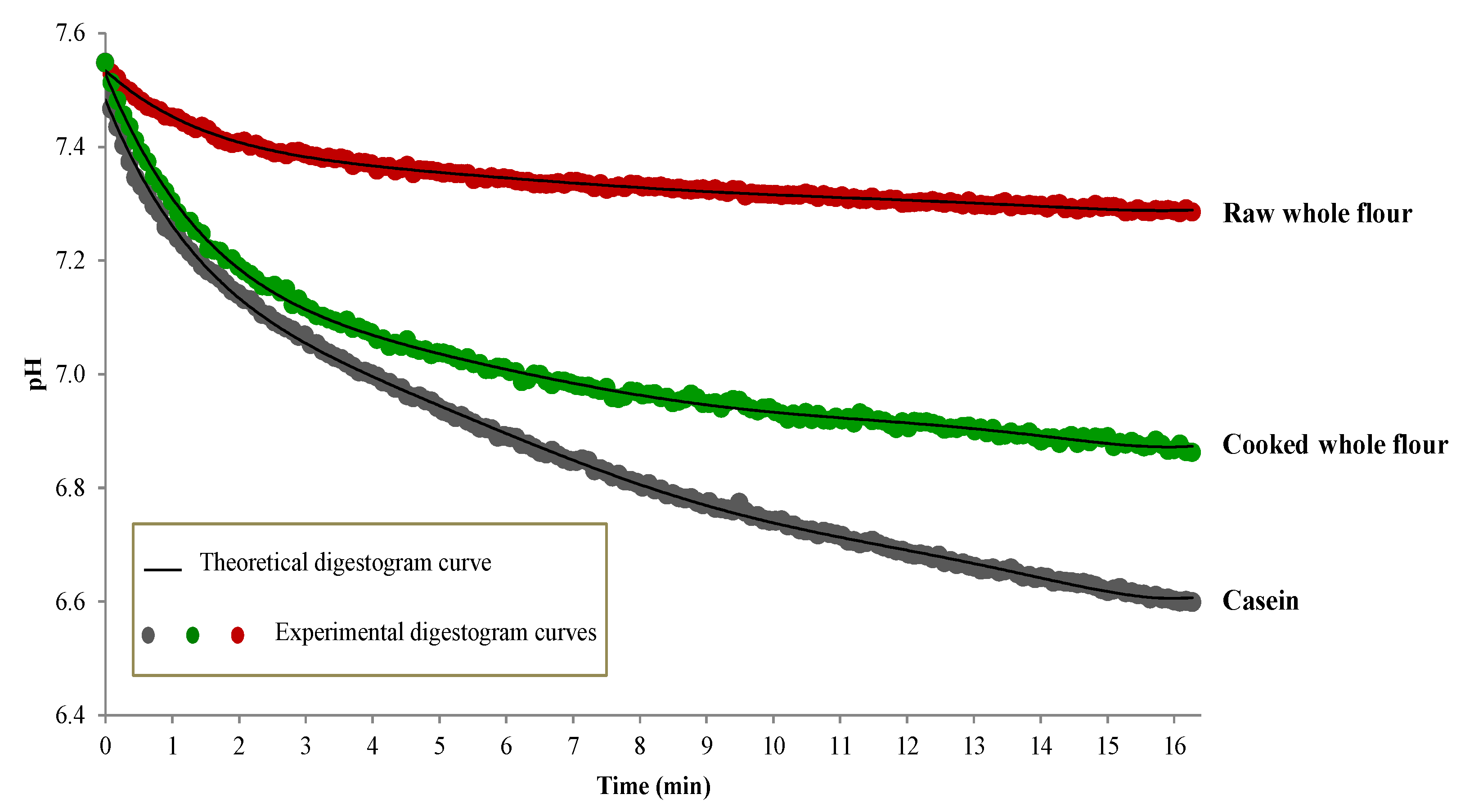
2.5.4. In Vitro Protein Digestibility
2.5.5. Calculated Protein Quality
2.6. Statistical Analyses
3. Results and Discussion
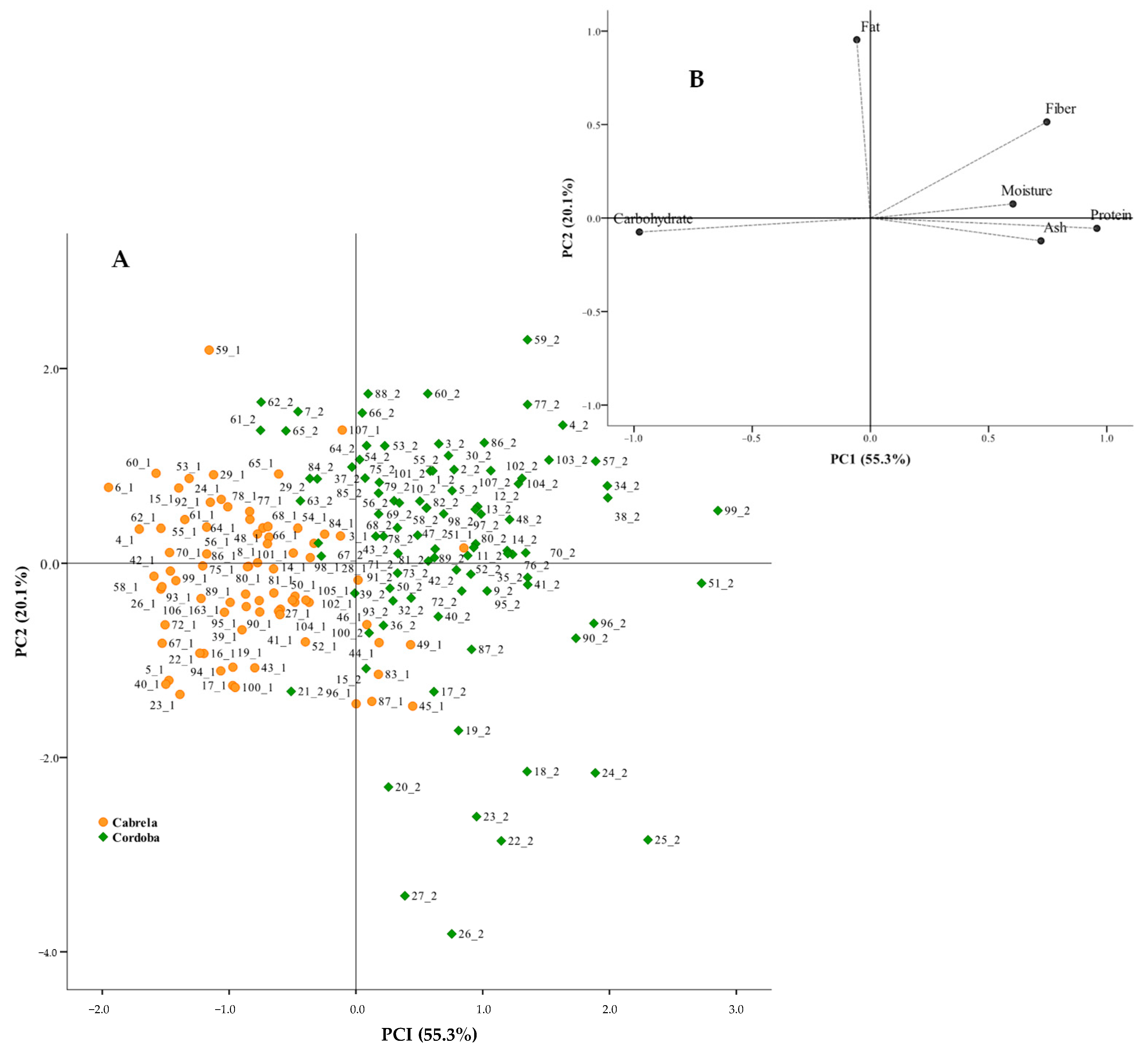
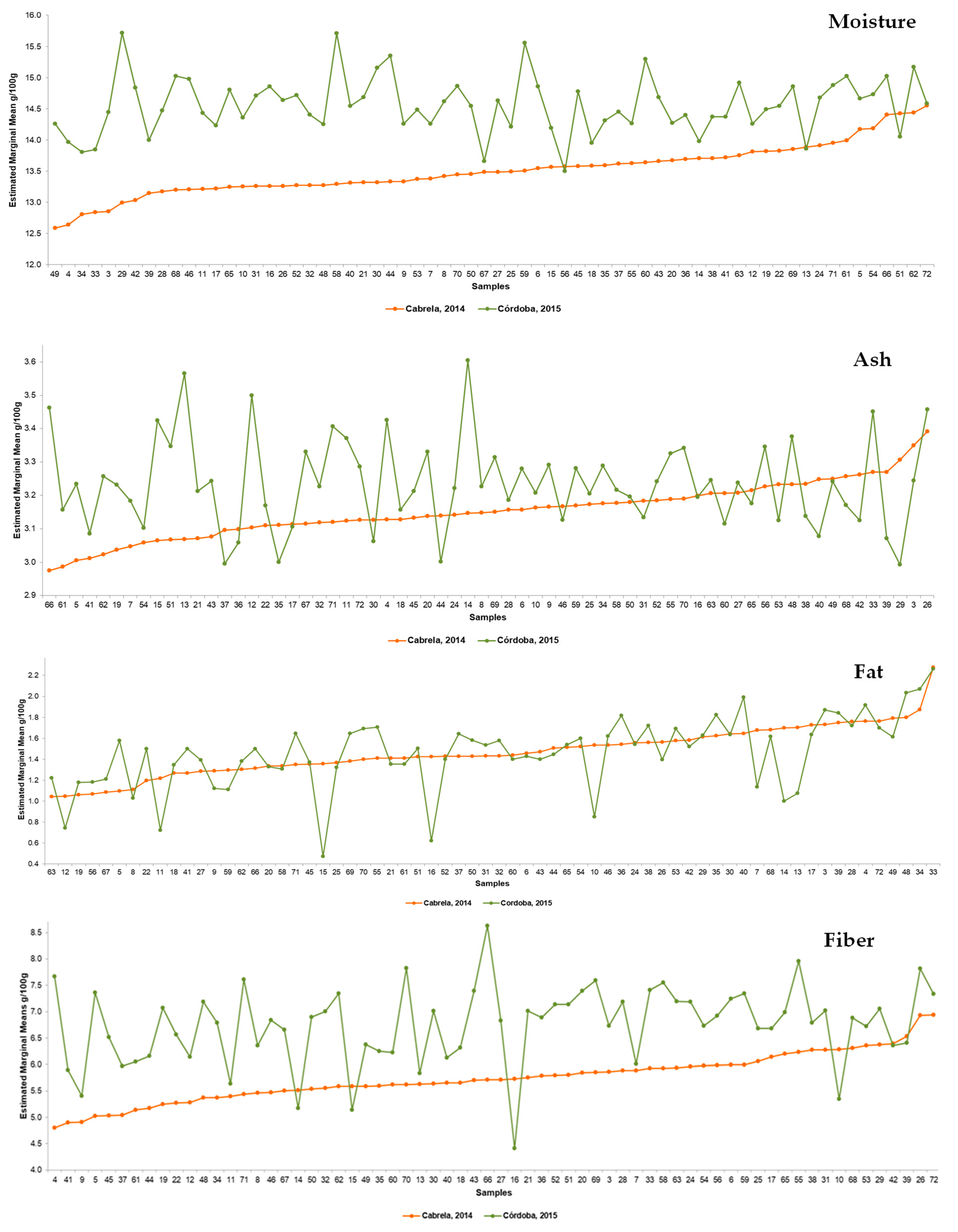
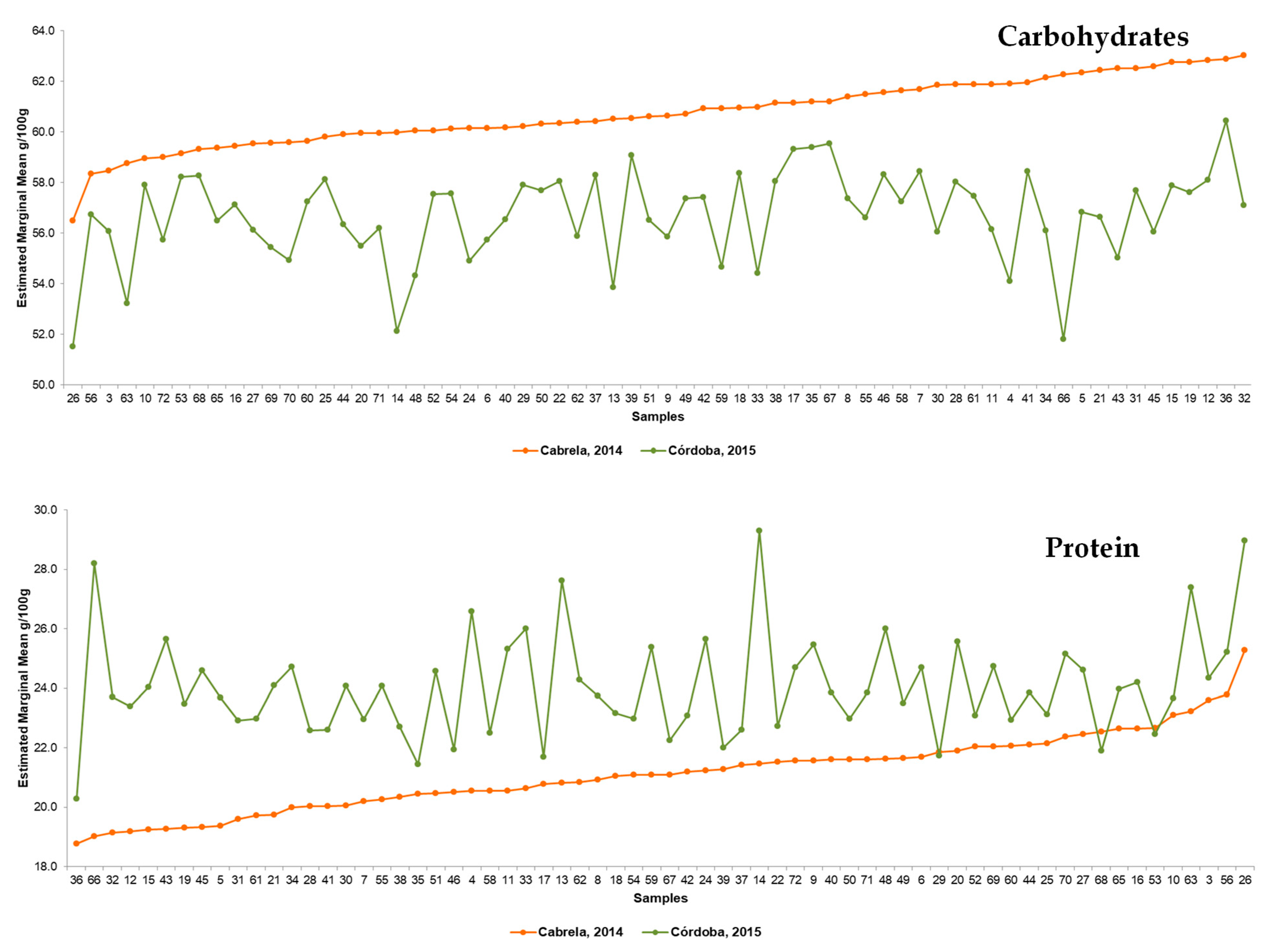
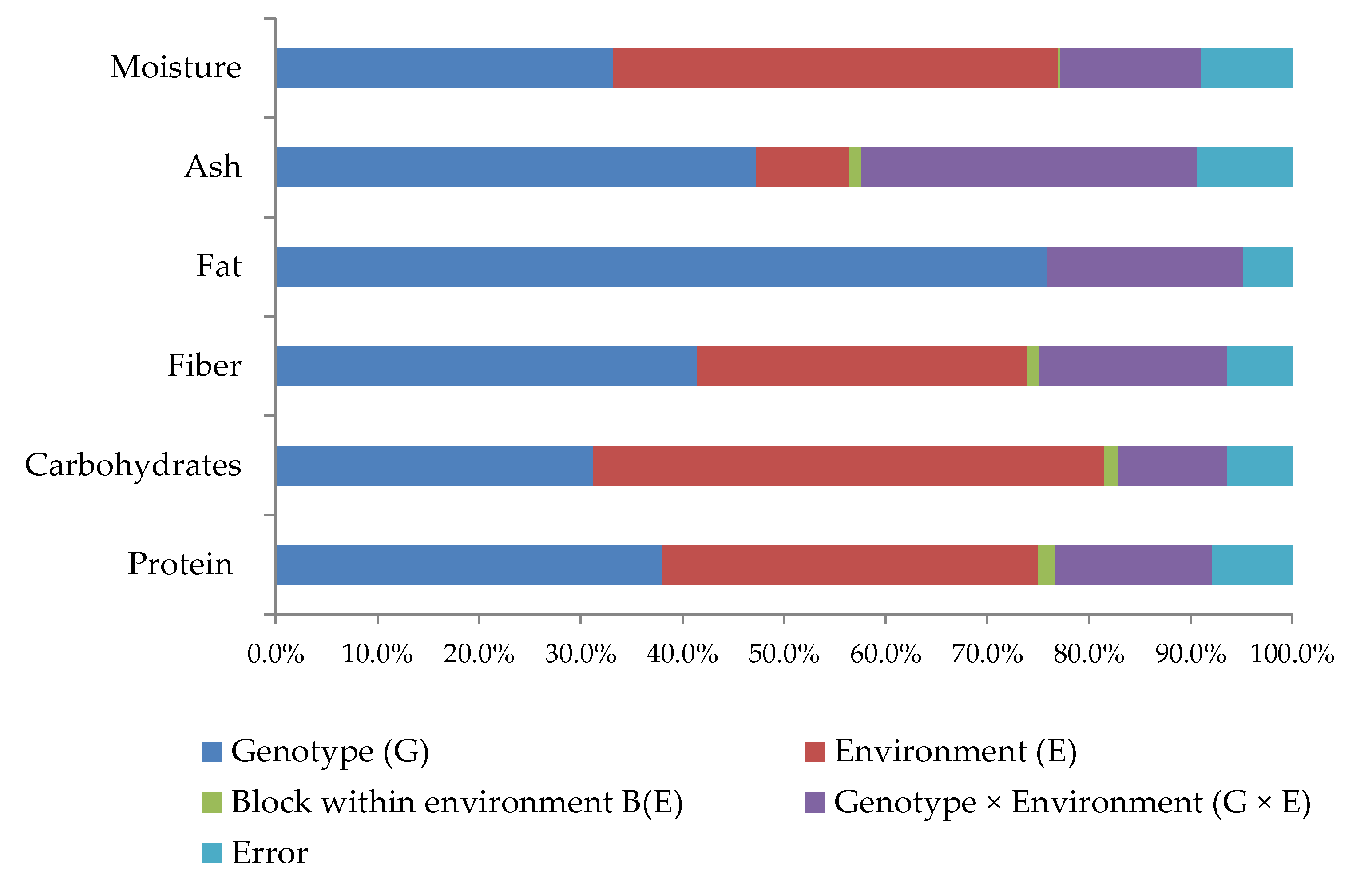
3.1. Diversity in the Nutritional Composition of Portuguese Common Beans
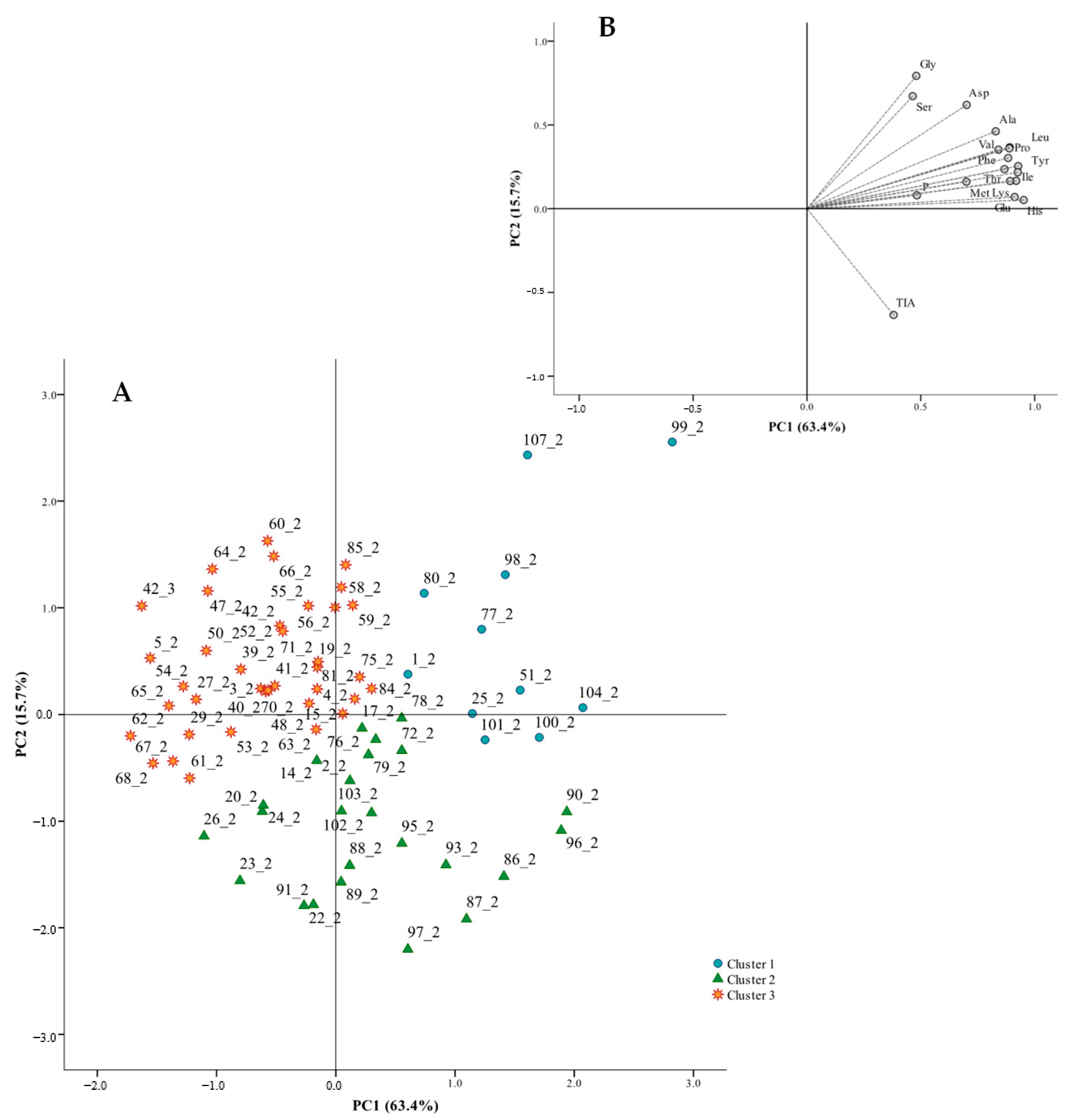
3.2. Diversity in the Protein Quality of Portuguese Common Beans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UNICEF; WHO; World Bank Group. Levels and Trends in Child Malnutrition 2020. Available online: https://www.unicef.org/media/69816/file/Joint-malnutrition-estimates-2020.pdf (accessed on 29 September 2020).
- Miller, B.D.D.; Welch, R.M. Food system strategies for preventing micronutrient malnutrition. Food Policy 2013, 42, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Klag, M.J.; Christian, P. Chapter 1.2 Population growth and malnutrition. In Good Nutrition: Perspectives for the 21st Century, 1st ed.; Eggersdorfer, M., Kraemer, K., Cordaro, J.B., Fanzo, J., Gibney, M., Kennedy, E., Labrique, A., Steffen, J., Eds.; Karger: Basel, Switzerland, 2016; pp. 31–44. [Google Scholar]
- Alemu, E.A. Malnutrition and its implications on food security. In Zero Hunger, 1st ed.; Leal Filho, W., Azul, A.M., Brandli, L., Özuyar, P.G., Wall, T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–10. [Google Scholar]
- Courty, P.-E.; Smith, P.; Koegel, S.; Redecker, D.; Wipf, D. Inorganic nitrogen uptake and transport in beneficial plant root-microbe interactions. CRC Crit. Rev. Plant. Sci. 2015, 34. [Google Scholar] [CrossRef]
- Peix, A.; Ramírez-Bahena, M.H.; Velázquez, E.; Bedmar, E.J. Bacterial associations with legumes. CRC Crit. Rev. Plant Sci. 2015, 34, 17–42. [Google Scholar] [CrossRef]
- FAO. FAOSTAT. 2017. Available online: http://www.fao.org/faostat/en/#compare (accessed on 20 May 2020).
- Rubiales, D.; Mikic, A. Introduction: Legumes in sustainable agriculture. CRC Crit. Rev. Plant. Sci. 2015, 34, 2–3. [Google Scholar] [CrossRef] [Green Version]
- Pelzer, E.; Bourlet, C.; Carlsson, G.; Lopez-Bellido, R.J.; Jensen, E.S.; Jeuffroy, M.-H. Design, assessment and feasibility of legume-based cropping systems in three European regions. Crop Pasture Sci. 2017, 68, 902–914. [Google Scholar] [CrossRef] [Green Version]
- Vadez, V.; Berger, J.D.; Warkentin, T.; Asseng, S.; Ratnakumar, P.; Rao, K.P.C.; Gaur, P.M.; Munier-Jolain, N.; Larmure, A.; Voisin, A.-S.; et al. Adaptation of grain legumes to climate change: A review. Agron. Sustain. Dev. 2012, 32, 31–44. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; HanumanthaRao, B.; Nair, R.M.; Vara Prasad, P.V.; Kumar, S.; Gaur, P.M.; Farooq, M.; Siddique, K.H.M.; Varshney, R.K.; et al. Food legumes and rising temperatures: Effects, adaptive functional mechanisms specific to reproductive growth stage and strategies to improve heat tolerance. Front. Plant Sci. 2017, 1658, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Izaguirre-Mayoral, M.L.; Lazarovits, G.; Baral, B. Ureide metabolism in plant-associated bacteria: Purine plant-bacteria interactive scenarios under nitrogen deficiency. Plant. Soil 2018, 428, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Leal, J.L.; Gálvez-Valdivieso, G.; Fernández, J.; Pineda, M.; Alamillo, J.M. Developmental effects on ureide levels are mediated by tissue-specific regulation of allantoinase in Phaseolus vulgaris L. J. Exp. Bot. 2012, 63, 4095–4106. [Google Scholar] [CrossRef]
- Schumacher, H.; Paulsen, H.M.; Gau, A.E.; Link, W.; Jürgens, H.U.; Sass, O.; Dieterich, R. Seed protein amino acid composition of important local grain legumes Lupinus angustifolius L., Lupinus luteus L., Pisum sativum L. and Vicia faba L. Plant Breed. 2011, 130, 156–164. [Google Scholar] [CrossRef]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition. Available online: http://www.fao.org/ag/humannutrition/35978-02317b979a686a57aa4593304ffc17f06.pdf (accessed on 13 May 2020).
- Vnučec, D.; Kutnar, A.; Goršek, A. Soy-based adhesives for wood-bonding–A review. J. Adhes. Sci. Technol. 2017, 31, 910–931. [Google Scholar] [CrossRef]
- Hou, Y.; Yin, Y.; Wu, G. Dietary essentiality of “nutritionally non-essential amino acids” for animals and humans. Exp. Biol. Med. 2015, 240, 997–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbano, G.; López-Jurado, M.; Frejnagel, S.; Gómez-Villalva, E.; Porres, J.M.; Frías, J.; Vidal-Valverde, C.; Aranda, P. Nutritional assessment of raw and germinated pea (Pisum sativum L.) protein and carbohydrate by in vitro and in vivo techniques. Nutrition 2005, 21, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Nosworthy, M.G.; House, J.D. Factors influencing the quality of dietary proteins: Implications for pulses. Cereal Chem. 2017, 94, 49–57. [Google Scholar] [CrossRef]
- Vaz Patto, M.C. Grain legume protein quality: A hot subject. Arbor 2016, 192, a314. [Google Scholar] [CrossRef] [Green Version]
- Leitão, S.T.; Dinis, M.; Veloso, M.M.; Šatović, Z.; Vaz Patto, M.C. Establishing the bases for introducing the unexplored Portuguese common bean germplasm into the breeding world. Front. Plant. Sci. 2017, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Time and Date AS. Available online: https://www.timeanddate.com/weather/ (accessed on 13 March 2020).
- FAO. Key to the FAO Soil Units (1974). Available online: http://www.fao.org/soils-portal/soil-survey/soil-classification/fao-legend/key-to-the-fao-soil-units/en/ (accessed on 3 August 2020).
- Serrano, C.; Carbas, B.; Castanho, A.; Soares, A.; Vaz Patto, M.C.; Brites, C. Characterisation of nutritional quality traits of a chickpea (Cicer arietinum) germplasm collection exploited in chickpea breeding in Europe. Crop Pasture Sci. 2017, 68, 1031–1040. [Google Scholar] [CrossRef]
- USDA. Food Data Central. 2019. Available online: https://fdc.nal.usda.gov/ (accessed on 20 May 2020).
- Igor, J.; Krstović, S.; Glamocic, D.; Jakšić, S.; Abramović, B. Validation of an HPLC method for the determination of amino acids in feed. J. Serb. Chem. Soc. 2013, 78, 839–850. [Google Scholar]
- Shrivastava, A.; Gupta, V. Methods for the determination of limit of detection and limit of quantitation of the analytical methods. Chron. Young Sci. 2011, 2, 21–25. [Google Scholar] [CrossRef]
- ISO. ISO 14902:2001(en). Animal Feeding Stuffs—Determination of Trypsin Inhibitor Activity of Soya Products; ISO: Geneva, Switzerland, 2001. [Google Scholar]
- Tinus, T.; Damour, M.; van Riel, V.; Sopade, P.A. Particle size-starch–protein digestibility relationships in cowpea (Vigna unguiculata). J. Food Eng. 2012, 113, 254–264. [Google Scholar] [CrossRef]
- Drulyte, D.; Orlien, V. The Effect of processing on digestion of legume proteins. Foods 2019, 8, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Institute of Agriculture and Natural Resources-University Nebraska-Lincoln Food. How to Cook Dry Beans from Scratch. Available online: https://food.unl.edu/article/how-cook-dry-beans-scratch (accessed on 4 November 2020).
- Young, V.R.; Pellett, P.L. Plant proteins in relation to human protein and amino acid nutrition. Am. J. Clin. Nutr. 1994, 59, 1203S–12l2S. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Cavada, E.; Juan, R.; Pastor, J.E.; Alaiz, M.; Vioque, J. Protein and amino acid composition of select wild legume species of tribe Fabeae. Food Chem. 2014, 163, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimelis, E.A.; Rakshit, S.K. Proximate composition and physico-chemical properties of improved dry bean (Phaseolus vulgaris L.) varieties grown in Ethiopia. LWT 2005, 38, 331–338. [Google Scholar] [CrossRef]
- Wilcox, J.R.; Guodong, Z. Relationships between seed yield and seed protein in determinate and indeterminate soybean populations. Crop Sci. 1997, 37. [Google Scholar] [CrossRef]
- Solberg, S.Ø.; Yndgaard, F.; Poulsen, G.; von Bothmer, R. Seed yield and protein content in the Weibullsholm Pisum collection. Genet. Resour. Crop Evol. 2017, 64, 2035–2047. [Google Scholar] [CrossRef]
- Florez, A.; Pujolà, M.; Valero, J.; Centelles, E.; Almirall, A.; Casañas, F. Genetic and environmental effects on chemical composition related to sensory traits in common beans (Phaseolus vulgaris L.). Food Chem. 2009, 113, 950–956. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 1705, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Ceccarelli, S. Specific adaptation and breeding for marginal conditions. Euphytica 1994, 77, 205–219. [Google Scholar] [CrossRef]
- Gardner, C.D.; Hartle, J.C.; Garrett, R.D.; Offringa, L.C.; Wasserman, A.S. Maximizing the intersection of human health and the health of the environment with regard to the amount and type of protein produced and consumed in the United States. Nutr. Rev. 2019, 77, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Aremu, M.O.; Olaofe, O.; Akintayo, T.E. A comparative study on the chemical and amino acid composition of some Nigerian under-utilized legume flours. Pak. J. Nutr. 2006, 5, 34–38. [Google Scholar]
- Baptista, A.; Pinho, O.; Pinto, E.; Casal, S.; Mota, C.; Ferreira, I. Characterization of protein and fat composition of seeds from common beans (Phaseolus vulgaris L.), cowpea (Vigna unguiculata L. Walp) and bambara groundnuts (Vigna subterranea L. Verdc) from Mozambique. Food Meas. 2017, 11, 442–450. [Google Scholar] [CrossRef]
- Khattab, R.Y.; Arntfield, S.D.; Nyachoti, C.M. Nutritional quality of legume seeds as affected by some physical treatments, Part 1: Protein quality evaluation. LWT 2009, 42, 1107–1112. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. The impact of dietary protein intake on longevity and metabolic health. EBioMedicine 2019, 43, 632–640. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, J.K.; Voutilainen, S.; Rissanen, T.H.; Happonen, P.; Mursu, J.; Laukkanen, J.A.; Poulsen, H.; Lakka, T.A.; Salonen, J.T. High dietary methionine intake increases the risk of acute coronary events in middle-aged men. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 113–120. [Google Scholar] [CrossRef]
- Nosworthy, M.G.; Franczyk, A.; Zimoch-Korzycka, A.; Appah, P.; Utioh, A.; Neufeld, J.; House, J.D. Impact of processing on the protein quality of pinto bean (Phaseolus vulgaris) and buckwheat (Fagopyrum esculentum Moench) flours and blends, as determined by in vitro and in vivo methodologies. J. Agric. Food Chem. 2017, 65, 3919–3925. [Google Scholar] [CrossRef]
- Gabriella, P.; Csapó, J.; Varga-Visi, E. Determination of the enantiomers of methionine and cyst(e)ine in the form of methionine-sulphon and cysteic acid after performic acid oxidation by reversed phase high performance liquid chromatography. Agric. Conspec. Sci. 2003, 68, 269–273. [Google Scholar]
- Caire-Juvera, G.; Vázquez-Ortiz, F.A.; Grijalva-Haro, M. Amino acid composition, score and in vitro protein digestibility of foods commonly consumed in Norhwest Mexico. Nutr. Hosp. 2013, 28, 365–371. [Google Scholar] [PubMed]
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; Serna Saldívar, S.O. Inactivation methods of trypsin inhibitor in legumes: A review. J. Food Sci. 2018, 83, 17–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilani, G.; Cockell, K.; Sepehr, E. Effects of antinutritional factors on protein digestibility and amino acid availability in foods. J. Aoac. Int. 2005, 88, 967–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, H. Prevention of cancer and inflammation by soybean protease inhibitors. Front. Biosci. 2013, 5, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Rezende, A.A.; Pacheco, M.T.B.; da Silva, V.S.N.; de Castro Ferreira, T.A.P. Nutritional and protein quality of dry Brazilian beans (Phaseolus vulgaris L.). Food Sci. Technol. 2018, 38, 421–427. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Cabrela, 2014 (n = 79) | Córdoba, 2015 (n = 93) |
|---|---|---|
| Protein | 21.26 ± 1.42 a | 24.10 ± 1.66 b |
| Carbohydrates | 60.58 ± 1.50 b | 56.68 ± 1.75 a |
| Fat | 1.44 ± 0.23 a | 1.49 ±0.47 a |
| Fiber | 5.75 ± 0.44 a | 6.77 ± 0.72 b |
| Ash | 3.15 ± 0.08 a | 3.25 ± 0.13 b |
| Moisture | 13.56 ± 0.45 a | 14.49 ± 0.51 b |
| Parameters | Cluster 1 | Cluster 2 | Cluster 3 |
|---|---|---|---|
| Protein content, g/100 g | 25.33 ± 2.53 b | 24.35 ± 1.32 a,b | 23.48 ± 1.40 a |
| Amino acids composition, g/100 g (g/16 g N) | |||
| Gly | 1.27 ± 0.11 c (5.06 ± 0.60) | 1.00 ± 0.09 a (4.12 ± 0.39) | 1.09 ± 0.09 b (4.67 ± 0.38) |
| Ala | 1.32 ± 0.12 b (5.24 ± 0.63) | 1.07 ± 0.08 a (4.41 ± 0.41) | 1.05 ± 0.09 a (4.46 ± 0.40) |
| Ser | 1.52 ± 0.15 c (6.05 ± 0.80) | 1.19 ± 0.16 a (4.89 ± 0.71) | 1.31 ± 0.13 b (5.59 ± 0.62) |
| Pro | 1.12 ± 0.10 b (4.45 ± 0.51) | 0.94 ± 0.09 a (3.86 ± 0.41) | 0.90 ± 0.07 a (3.86 ± 0.31) |
| Val | 1.44 ± 0.13 b (5.71 ± 0.55) | 1.22 ± 0.12 a (5.01 ± 0.54) | 1.17 ± 0.11 a (4.99 ± 0.42) |
| Thr | 1.08 ± 0.14 b (4.30 ± 0.62) | 0.87 ± 0.12 a (3.52 ± 0.47) | 0.81 ± 0.10 a (3.40 ± 0.35) |
| Ile | 1.37 ± 0.12 b (5.45 ± 0.69) | 1.13 ± 0.13 a (4.64 ± 0.50) | 1.06 ± 0.10 a (4.53 ± 0.37) |
| Leu | 2.40 ± 0.22 b (9.56 ± 1.16) | 1.96 ± 0.22 a (8.06 ± 0.84) | 1.89 ± 0.19 a (8.06 ± 0.69) |
| Asp | 2.98 ± 0.21 b (11.85 ± 1.25) | 2.29 ± 0.23 a (9.42 ± 0.93) | 2.39 ± 0.23 a (10.17 ± 0.91) |
| Glu | 5.87 ± 0.50 c (23.38 ± 3.06) | 5.01 ± 0.50 b (20.61 ± 1.92) | 4.56 ± 0.39 a (19.46 ± 1.77) |
| Met | 0.15 ± 0.03 b (0.61 ± 0.10) | 0.13 ± 0.03 a (0.53 ± 0.10) | 0.12 ± 0.02 a (0.50 ± 0.07) |
| His | 0.83 ± 0.07 c (3.30 ± 0.49) | 0.71 ± 0.07 b (2.92 ± 0.31) | 0.64 ± 0.06 a (2.74 ± 0.25) |
| Phe | 1.67 ± 0.11 b (6.63 ± 0.61) | 1.42 ± 0.15 a (5.82 ± 0.54) | 1.36 ± 0.12 a (5.81 ± 0.47) |
| Tyr | 1.00 ± 0.09 b (4.00 ± 0.53) | 0.84 ± 0.09 a (3.46 ± 0.40) | 0.78 ± 0.08 a (3.34 ± 0.33) |
| Lys | 1.76 ± 0.10 b (7.01 ± 0.60) | 1.57 ± 0.17 a (6.45 ± 0.75) | 1.45 ± 0.13 a (6.19 ± 0.59) |
| Arg | 1.36 ± 0.16 b (5.39 ± 0.54) | 1.09 ± 0.14 a (4.47 ± 0.52) | 0.99 ± 0.13 a (4.22 ± 0.43) |
| PER | |||
| PER1 | 3.46 ± 0.50 b | 2.81 ± 0.37 a | 2.81 ± 0.30 a |
| PER2 | 3.45 ± 0.47 b | 2.83 ± 0.35 a | 2.84 ± 0.29 a |
| PER3 | 2.82 ± 0.54 b | 2.05 ± 0.45 a | 2.12 ± 0.38 a |
| AAS, % | |||
| AAS_Val | 163.07 ± 15.74 b | 143.13 ± 15.30 a | 142.50 ± 12.01 a |
| AAS_Thr | 126.38 ± 18.30 b | 105.11 ± 14.03 a | 101.44 ± 11.56 a |
| AAS_Ile | 194.60 ± 24.69 b | 165.70 ± 17.97 a | 161.84 ± 13.11 a |
| AAS_Leu | 144.78 ± 17.52 b | 122.12 ± 12.76 a | 122.14 ± 10.50 a |
| AAS_Met | 24.39 ± 4.07 b | 21.37 ± 4.01 a | 19.77 ± 2.96 a |
| AAS_His | 173.78 ± 25.85 c | 153.49 ± 16.11 b | 144.07 ± 13.29 a |
| AAS_Phe+Tyr | 168.76 ± 17.92 b | 147.25 ± 14.39 a | 145.20 ± 12.04 a |
| AAS_Lys | 120.92 ± 10.29 b | 111.15 ± 12.85 a | 106.79 ± 10.21 a |
| Protease inhibitors | |||
| TIA, mg/g | 9.68 ± 2.75 b | 10.95 ± 2.06 b | 7.47 ± 2.00 a |
| # | Accessions | IVPD (Raw) % | IVPDCAAS (Raw) % | IVPD (Cooked) % |
|---|---|---|---|---|
| 19 | 698 | 69.66 ± 0.64 | 13.0 | - |
| 20 | 706 | 69.47 ± 0.81 | 11.8 | 75.55 ± 1.63 |
| 25 | 1644 | 69.22 ± 0.01 | 10.3 | 73.52 ± 1.75 |
| 29 | 1663 | 69.86 ± 0.18 | 13.0 | 76.78 ± 0.21 |
| 43 | 1984 | 69.06 ± 0.38 | 12.1 | - |
| 48 | 2179 | 68.78 ± 0.30 | 11.4 | 75.07 ± 0.41 |
| 64 | 4119 | 69.61 ± 0.52 | 11.0 | - |
| 65 | 4120 | 69.70 ± 0.13 | 10.3 | - |
| Casein | 88.96 ± 1.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mecha, E.; Natalello, S.; Carbas, B.; da Silva, A.B.; Leitão, S.T.; Brites, C.; Veloso, M.M.; Rubiales, D.; Costa, J.; Cabral, M.d.F.; et al. Disclosing the Nutritional Quality Diversity of Portuguese Common Beans—The Missing Link for Their Effective Use in Protein Quality Breeding Programs. Agronomy 2021, 11, 221. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11020221
Mecha E, Natalello S, Carbas B, da Silva AB, Leitão ST, Brites C, Veloso MM, Rubiales D, Costa J, Cabral MdF, et al. Disclosing the Nutritional Quality Diversity of Portuguese Common Beans—The Missing Link for Their Effective Use in Protein Quality Breeding Programs. Agronomy. 2021; 11(2):221. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11020221
Chicago/Turabian StyleMecha, Elsa, Sofia Natalello, Bruna Carbas, Andreia Bento da Silva, Susana T. Leitão, Carla Brites, Maria Manuela Veloso, Diego Rubiales, Judite Costa, Maria de Fátima Cabral, and et al. 2021. "Disclosing the Nutritional Quality Diversity of Portuguese Common Beans—The Missing Link for Their Effective Use in Protein Quality Breeding Programs" Agronomy 11, no. 2: 221. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11020221