
Direct Shoot Organogenesis from Lycium chinense Miller Leaf Explants and Assessment of Genetic Stability Using ISSR Markers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Germination and Explant Preparation
2.2. Culture Conditions and Regeneration of Shoots
2.3. Influence of Carbon Source and Basal Medium on Shoot Regeneration
2.4. Rooting and Plant Acclimatization
2.5. Micromorphological Studies
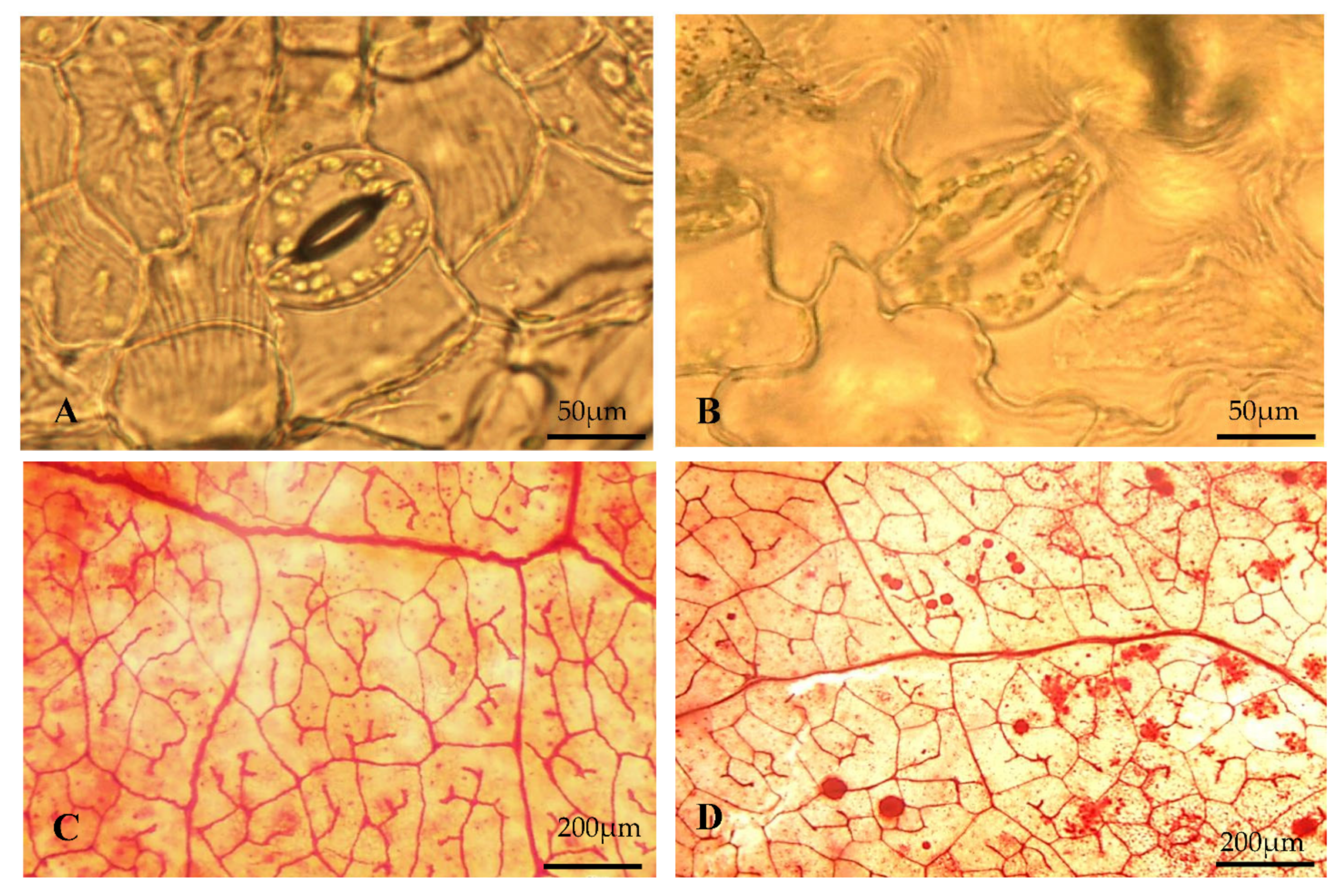
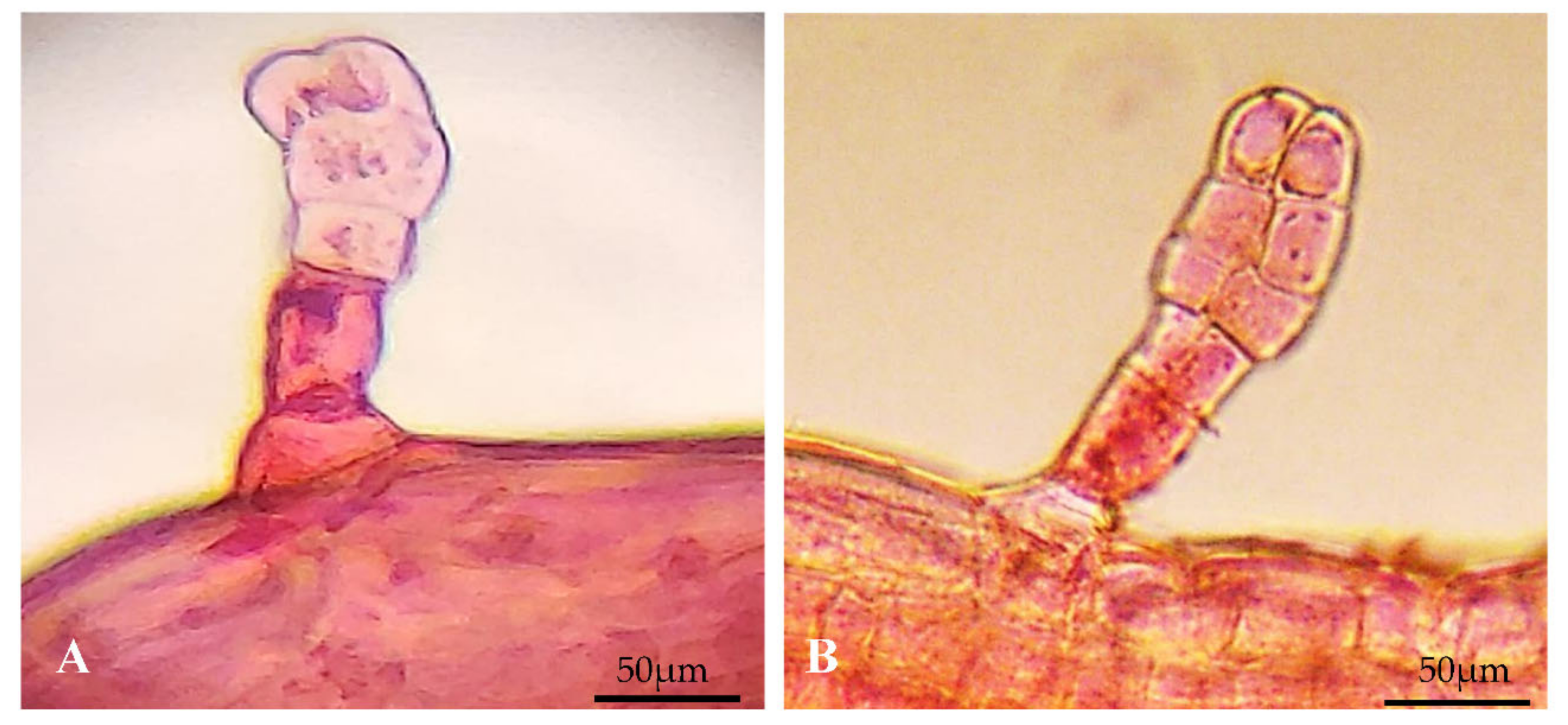
2.5.1. Venation Pattern and Trichome Density
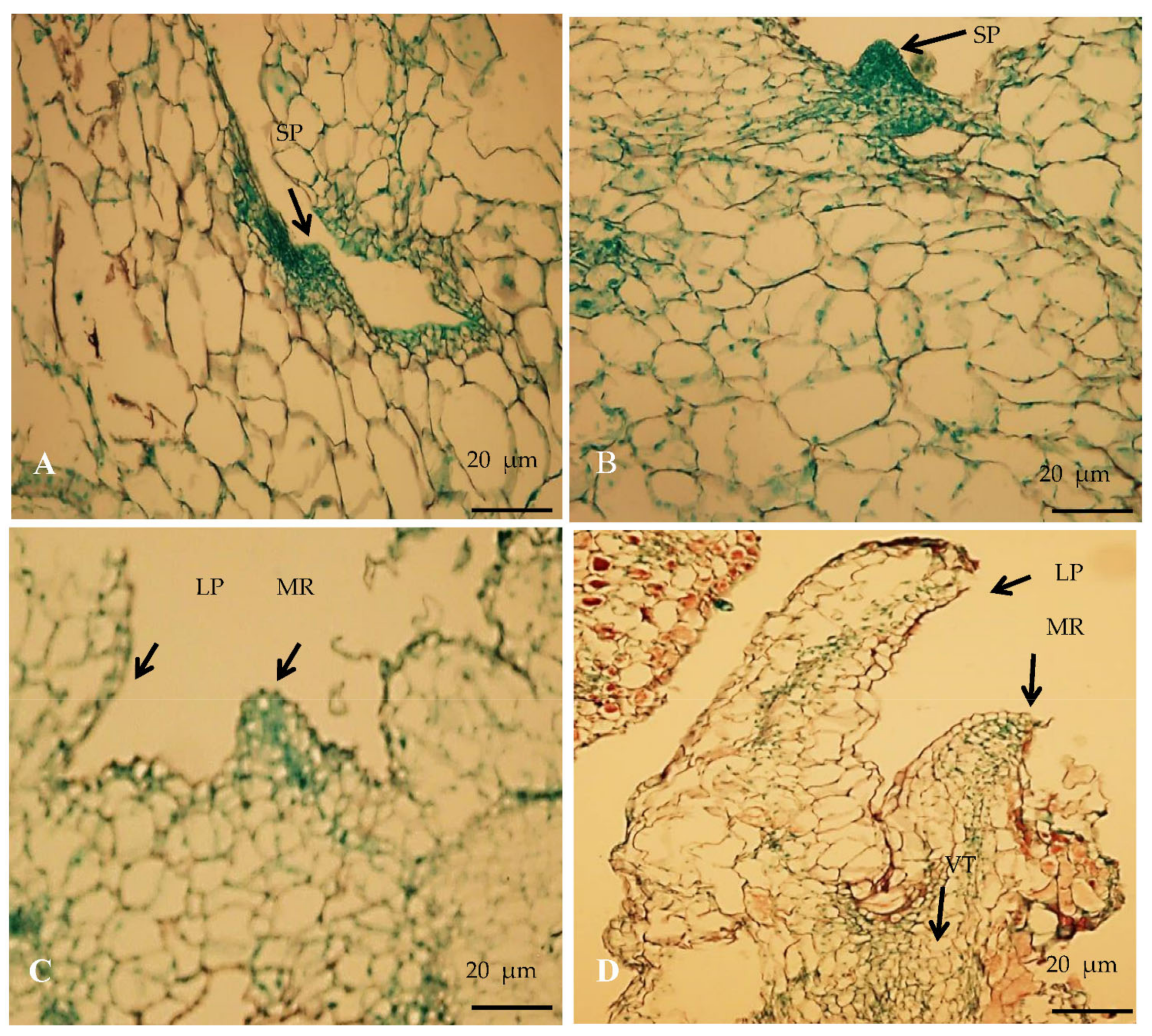
2.5.2. Histological Study of Regenerated Plants
2.5.3. Size and Density of Stomata
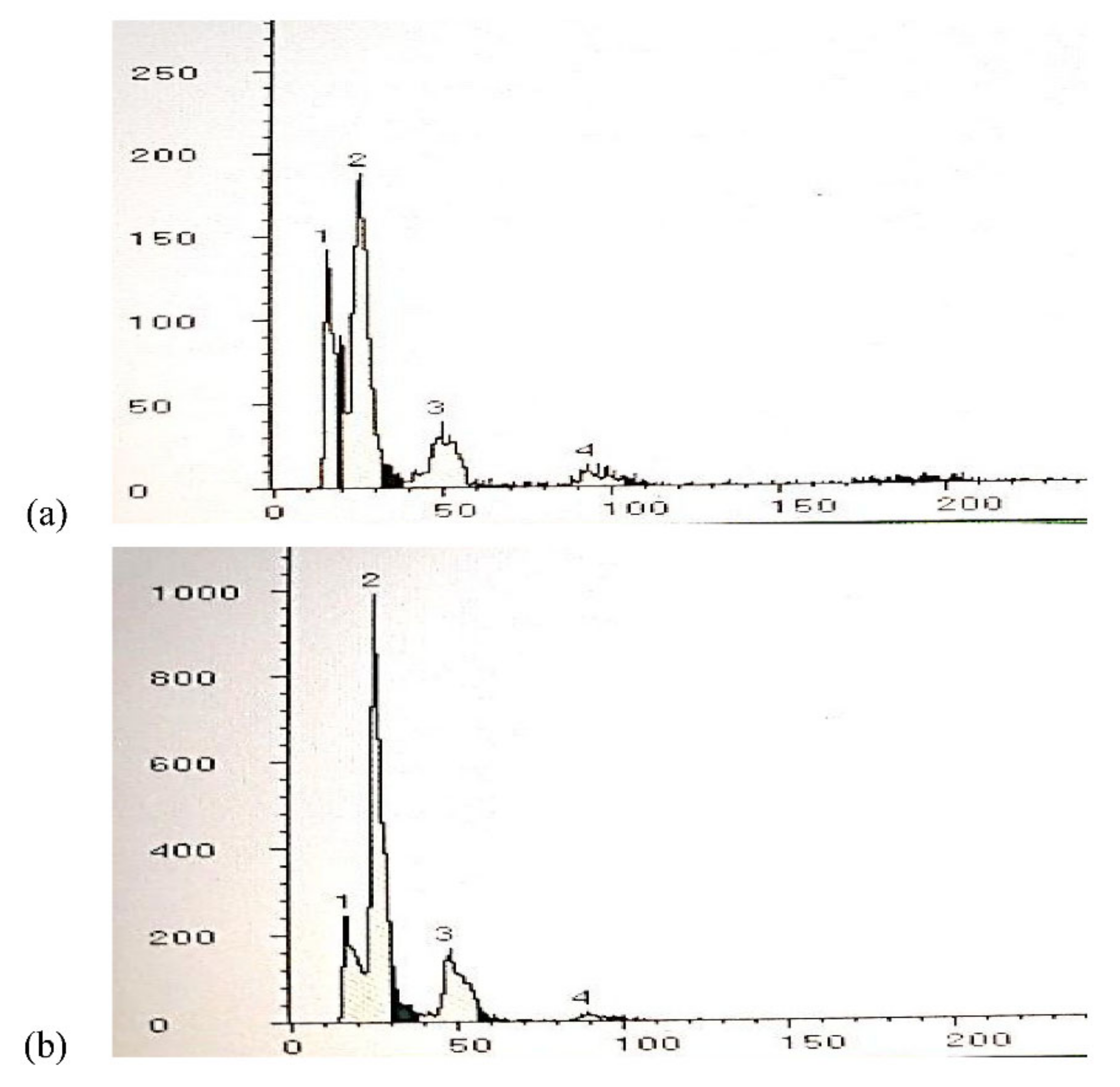
2.6. Assessment of Ploidy Level in Regenerants
2.7. Inter Simple Sequence Repeat (ISSR) Analysis
2.8. Statistical Analysis
3. Results
3.1. Effect of Plant Growth Regulators (PGRs) on Direct Shoot Regeneration
3.2. Influence of Basal Medium and Carbohydrate Source on Organogenesis
3.3. Effect of PGRs and Basal Salt Concentrations on Root Regeneration
3.4. Acclimatization
3.5. Histological Analysis of Regenerated Shoot
3.6. Micromorphological Studies
3.6.1. Stomata and Veinlets
3.6.2. Trichome
3.7. Assessment of Ploidy Levels of Regenerated Plants by Flow Cytometry
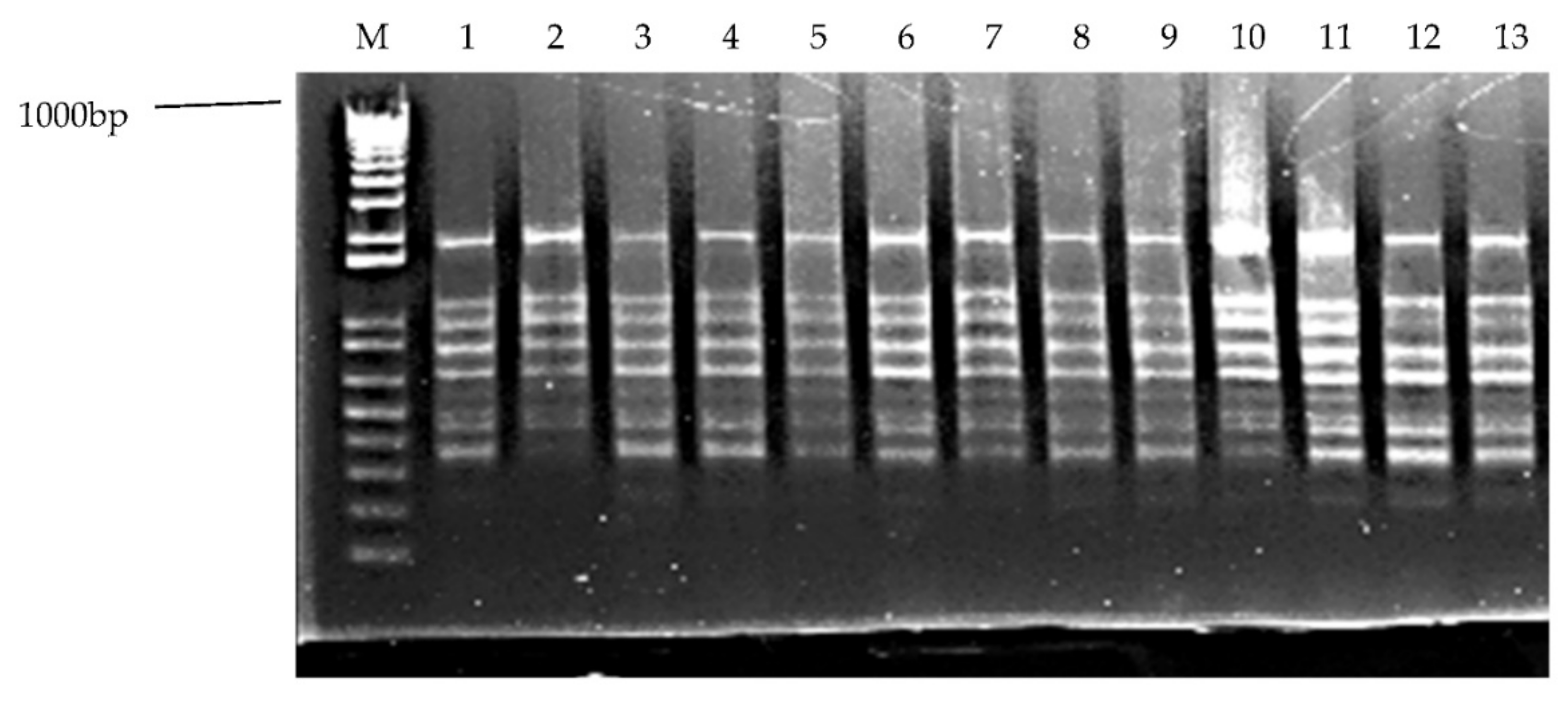
3.8. ISSR Analysis of Regenerated Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hong, D.Y.; Pan, K.Y.; Nicholas, J.T. Flora of China; Science Press: Beijing, China, 2004; pp. 1–9. [Google Scholar]
- Liu, S.C.; Lin, J.T.; Hu, C.C.; Shen, B.Y.; Chen, T.Y.; Chang, Y.L.; Shih, C.H.; Yang, D.J. Phenolic compositions and antioxidant attributes of leaves and stems from three inbred varieties of Lycium chinense Miller harvested at various times. Food Chem. 2017, 215, 284–291. [Google Scholar] [CrossRef]
- Le, K.; Chiu, F.; Ng, K. Identification and Quantification of Antioxidants in Fructus lycii. Food Chem. 2007, 105, 353–363. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, K.H.; Chang, K.S.; Bock, J.Y.; Jung, M.Y. Taste and flavor compounds in box thorn (Lycium chinense Miller) leaves. Food Chem. 1997, 58, 297–303. [Google Scholar] [CrossRef]
- Lee, D.G.; Jung, H.J.; Woo, E.R. Antimicrobial property of (+)-lyoniresinol-3alpha-O-beta-D-glucopyranoside isolated from the root bark of Lycium chinense Miller against human pathogenic microorganisms. Arch. Pharm. Res. 2005, 28, 1031–1036. [Google Scholar] [CrossRef]
- Wang, C.C.; Chang, S.C.; Inbaraj, B.S.; Chen, B.H. Isolation of carotenoids, flavonoids and polysaccharides from Lycium barbarum L. and evaluation of antioxidant activity. Food Chem. 2010, 120, 184–192. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Wu, T.; Liu, J.; Zhang, X.; Yang, X.; Goodheart, M.J.; Engelhardt, J.F. Selective suppression of cervical cancer Hela cells by 2-O-b-D-glucopyranosyl-L-ascorbic acid isolated from the fruit of Lycium barbarum L. Cell Biol. Toxicol. 2011, 27, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Liu, D.; Huang, A. The efficiency of flavonoids in polar extracts of Lycium chinense Mill. fruits as free radical scavenger. Food Chem. 2004, 87, 283–288. [Google Scholar] [CrossRef]
- Noculak-Palczewska, A.; Matkowski, A.; Gasiorowski, K.; Tabaka, H.; Oszmianski, J.; Lamer-Zarawska, E. Chemical characterisation of methanolic-water extracts from the fruit of acclimated Lycium chinense Mill. Herba Pol. 2004, 50, 47–53. [Google Scholar]
- Leung, I.; Tso, M.; Li, W.; Lam, T. Absorption and Tissue Distribution of Zeaxanthin and Lutein in Rhesus Monkeys after Taking Fructus lycii (Gou Qi Zi) Extract. Invest. Ophthalmol. Vis. Sci. 2001, 42, 466–471. [Google Scholar] [PubMed]
- Kim, S.Y.; Choi, Y.H.; Huh, H.; Kim, J.; Kim, Y.C.; Lee, H.S. New antihepatotoxic cerebroside from Lycium chinense fruits. J. Nat. Prod. 1997, 60, 274–276. [Google Scholar] [CrossRef]
- Noguchi, M.; Mochida, K.; Shingu, T.; Kozuka, M.; Fujitani, K. Uber die Bestandteile der chinesischen Droge ‘‘Ti-ku-‘pi’’.I. isolierung und constitution von lyciumamid, einem neuen dipeptid. Chem. Pharm. Bull. 1984, 32, 3584–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, Y.W.; Lim, S.W.; Kim, S.H.; Shin, D.Y.; Suh, Y.G.; Kim, Y.B.; Kim, Y.C.; Kim, J. Hepatoprotective pyrrole derivatives of Lycium chinense fruits. Bioorg. Med. Chem. Lett. 2003, 13, 79–81. [Google Scholar] [CrossRef]
- Kim, J.S.; Chung, H.Y. GC-MS Analysis of the Volatile Components in Dried Boxthorn (Lycium chinenesis) Fruit. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 516–524. [Google Scholar] [CrossRef]
- Potterat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, Pharmacology, and Safety in the Per-spective of Traditional Uses and Recent Popularity. Planta Med. 2010, 76, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Tudor, V.; Asănică, A.; Teodorescu, I.R.; Gîdea, M.; Tănăsescu, C.; Tudor, A.D.; Țiu, V.J. Germination capacity of some Lycium barbarum L., and Lycium chinense Mill. biotypes seed. Rom. Biotechnol. Lett. 2017, 22, 12191–12196. [Google Scholar]
- Kim, D.C.; Chung, H.J. Plant regeneration from explant types and cultivars of boxthorn (Lycium chinense Mill.). Korean J. Plant Biotechnol. 2002, 29, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Jo, M.H.; Ham, I.K.; Lee, B.C.; Kim, J.W.; Lee, W.S.; Kwon, S.Y.; Lee, H.S.; Kwak, S.S. High frequency shoot formation and plant regeneration from Cotyledonary hypocotyl explants of boxthorn (Lycium chinense Mill.) seedlings. Korean J. Plant Biotechnol. 2004, 31, 203–207. [Google Scholar]
- Larkins, P.J.; Seowcroft, W.R. Somaclonal variation—A novel source of variability from cell cultures for plant improvement. Theor. Appl. Genet. 1981, 60, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Nyman, L.P.; Gonzales, C.J.; Arditti, J. In vitro selection for salt tolerance of taro (Colocasia esculenta vat. antiquorum). Ann. Bot. 1983, 51, 229–236. [Google Scholar] [CrossRef]
- Rani, V.; Raina, S.N. Genetic fedility of organized meristem-derived micropropagated plants: A critical reappraisal. Vitr. Cell. Dev. Biol. Plant 1998, 36, 319–330. [Google Scholar] [CrossRef]
- Dey, M.; Bakshi, S.; Galiba, G.; Sahoo, L.; Panda, S.K. Development of a genotype independent and transformation amenable regeneration system from shoot apex in rice (Oryza sativa spp. indica) using TDZ. 3 Biotech. 2012, 2, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Sticklen, M.B.; Oraby, H.F. Shoot apical meristem: A sustainable explant for genetic transformation of cereal crops. Vitr. Cell Dev. Biol. Plant 2005, 41, 187–200. [Google Scholar] [CrossRef]
- Bramhanapalli, M.; Thogatabalija, L.; Gudipalli, P. Efficient in vitro plant regeneration from seedling-derived explants and genetic stability analysis of regenerated plants of Simarouba glauca DC. by RAPD and ISSR markers. Vitr. Cell Dev. Biol. Plant 2016, 53, 50–63. [Google Scholar] [CrossRef]
- Romyanon, K.; Mosaleeyanon, K.; Kirdmanee, C. Direct-shoot organogenesis as an alternative protocol for in vitro regeneration of oil palm (Elaeis guineensis Jacq.). Sci. Hortic. 2015, 195, 1–7. [Google Scholar] [CrossRef]
- Czene, S.; Harms-Ringdahl, M. Detection of single-strand breaks and formamidopyrimidine-DNA glycosylase-sensitive sites in DNA of cultured human fibroblasts. Mutat. Res. DNA Repair 1995, 336, 235–242. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Iwase, A.; Sugimoto, K. Control of plant cell diferentiation by histone modifcation and DNA methylation. Curr. Opin. Plant Biol. 2015, 28, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Seo, P.J. Dynamic epigenetic changes during plant regeneration. Trends Plant Sci. 2018, 23, 235–247. [Google Scholar] [CrossRef]
- Cui, C.; Li, Y.; Liu, Y.; Li, X.; Luo, S.; Zhang, Z.; Wu, R.; Liang, G.; Sun, J.; Peng, J.; et al. Determination of genetic diversity among Saccharina germplasm using ISSR and RAPD markers. C. R. Biol. 2017, 340, 76–86. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genetic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–479. [Google Scholar] [CrossRef]
- Gamborg, L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A.C. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Seong, E.S.; Nguyen, T.X.; Yu, C.Y.; Kim, S.H.; Chung, I.M. In vitro regeneration of Melastoma malabatricum Linn. through organogenesis and assessment of clonal and biochemical fidelity using RAPD and HPLC. Plant Cell Tiss. Org. Cult. 2016, 124, 517–529. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Yu, C.Y.; Chung, I.M. Direct shoot organogenesis and assessment of genetic stability in regenerants of Solanum aculeatissimum Jacq. Plant Cell Tiss. Org. Cult. 2012, 108, 455–464. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13. [Google Scholar]
- Cappelletti, R.; Sabbadini, S.; Mezzetti, B. The use of TDZ for the efficient in vitro regeneration and organogenesis of strawberry and blueberry cultivars. Sci. Hortic. 2016, 207, 117–124. [Google Scholar] [CrossRef]
- Gurel, E.; Topal, E.; Gurel, S. The effect of pretreating seedlings with BAP on direct shoot regeneration from periole explants of sugar beet (Beta vulgaris L.). Biotechnol. Biotechnol. Eq. 2003, 17, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Bhagwat, B.; Lane, W.D. In vitro shoot regeneration from leaves of sweet cherry (Prunus avium) ‘Lapins’ and ‘Sweetheart’. Plant Cell Tiss. Org. Cult. 2004, 78, 173–181. [Google Scholar] [CrossRef]
- Kumar, P.P.; Dimps Rao, C.; Goh, C.J. Influence of petiole and lamina on adventitious shoot initiation from leaf explants of Paulownia fortunei. Plant Cell Rep. 1998, 17, 886–890. [Google Scholar] [CrossRef]
- Deore, A.C.; Johnson, T.S. High-frequency plant regeneration from leaf-disc cultures of Jatropha curcas L.: An important biodiesel plant. Plant Biotechnol. Rep. 2008, 2, 7–11. [Google Scholar] [CrossRef]
- Kumar, N.; Vijay Anand, K.G.; Reddy, M.P. Shoot regeneration from cotyledonary leaf explants of Jatropha curcas a biodiesel plant. Acta Physiol. Plant 2010, 32, 917–924. [Google Scholar] [CrossRef]
- Jones, M.P.A.; Cao, J.; O’Brien, R.; Murch, S.J.; Saxena, P.K. The mode of action of thidiazuron: Auxins, indoleamines, and ion channels in the regeneration of Echinacea purpurea L. Plant Cell Rep. 2007, 26, 1481–1490. [Google Scholar] [CrossRef]
- Zhang, C.R.; Huang, X.L.; Wu, J.Y.; Feng, B.H.; Chen, Y.F. Identification of thidiazuron-induced ESTs expressed differentially during callus differentiation of alfalfa (Medicago sativa). Physiol. Plant 2006, 128, 732–739. [Google Scholar] [CrossRef]
- Huetteman, C.A.; Preece, J.E. Thidiazuron: A potent cytokinin for woody plant tissue culture. Plant Cell Tiss. Org. Cult. 1993, 33, 105–119. [Google Scholar] [CrossRef]
- Murthy, B.N.S.; Murch, S.J.; Saxena, P.K. Thidiazuron: A potent regulator of in vitro plant morphogenesis. Vitr. Cell Dev. Biol. Plant 1998, 34, 267–275. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Seong, E.S.; Goh, E.J.; Kim, N.Y.; Kang, W.H.; Kim, E.H.; Yu, C.Y.; Chung, I.M. High-frequency direct shoot regeneration from Drymaria cordata Willd. leaves. Plant Cell Tiss. Org. Cult. 2010, 100, 209–217. [Google Scholar] [CrossRef]
- Karam, N.S.; Al-Majathoub, M. Direct shoot regeneration and microtuberization in wild Cyclamen persicum Mill. using seedling tissue. Sci. Hort. 2000, 86, 235–246. [Google Scholar] [CrossRef]
- Ďurkovič, J.; Alžbeta Lengyelová, A.; Ingrid Čaňová, I.; Daniel Kurjak, D.; Hladka, D. Photosynthetic performance and stomatal characteristics during ex vitro acclimatization of true service tree (Sorbus domestica L.). J. Hort. Sci. Biotechnol. 2009, 84, 223–227. [Google Scholar] [CrossRef]
- Mani, M.; Shekhawat, M.S. Foliar micromorphology of in vitro-cultured shoots and field-grown plants of Passiflora foetida. Hortic. Plant J. 2017, 3, 34–40. [Google Scholar] [CrossRef]
- Carvalho Luisa, C.; Leonor Osório, M.; Manuela Chaves, M.; Amâncio, S. Chlorophyll fluorescence as an indicator of photosynthetic functioning of in vitro grapevine and chestnut plantlets under ex vitro acclimatization. Plant Cell Tiss. Org. Cult. 2001, 67, 271–280. [Google Scholar] [CrossRef]
- Revathi, J.; Manokari, M.; Shekhawat, M.S. Optimization of factors affecting in vitro regeneration, flowering, ex vitro rooting and foliar micromorphological studies of Oldenlandia corymbosa L.: A multipotent herb. Plant Cell Tiss. Org. Cult. 2018, 134, 1–13. [Google Scholar] [CrossRef]
- Kozai, T.; Iwabuchi, K.; Watanabe, K.; Watanabe, I. Photoautotrophic and photomixotrophic growth of strawberry plantlets in vitro and changes in nutrient composition of the medium. Plant Cell Tiss. Org. Cult. 1991, 25, 107–115. [Google Scholar]
- Gaspar, T.; Franck, T.; Bisbis, B.; Kevers, C.; Jouve, L.; Hausman, J.F.; Dommes, J. Concepts in plant stress physiology. Application to plant tissue cultures. Plant Growth Regul. 2002, 37, 263–285. [Google Scholar] [CrossRef]
- Manokari, M.; Shekhawat, M.S. Comprehensive analysis of in vitro to feld transition of micromorphology and leaf architecture in Passiflora edulis Sims. f. flavicarpa Deg. Ind. J. Plant Physiol. 2017, 22, 240–246. [Google Scholar] [CrossRef]
- Wang, F.; Xin, X.; Wei, H.; Qiu, X.; Liu, B. In vitro regeneration, ex vitro rooting and foliar stoma studies of Pseudostellaria heterophylla (Miq.) Pax. Agronomy 2020, 10, 949. [Google Scholar] [CrossRef]
- Shekhawat Mahipal, S.; Manokari, M.; Kannan, N. Micromorphological response towards altered environmental conditions in subsequent stages of in vitro propagation of Morinda coreia. Environ. Exp. Biol. 2017, 15, 37–46. [Google Scholar]
- Saez, P.L.; Bravo, L.A.; Latsagne, M.I.; Rios, D.G. Increased light intensity during in vitro culture improves water loss control and photosynthetic performance of Castanea sativa grown in ventilated vessels. Sci. Hortic. 2012, 138, 7–16. [Google Scholar] [CrossRef]
- Machado, M.P.; Silva, A.L.L.; Biasi, L.A.; Deschamps, C.; Filho, J.C.B.; Zanette, F. Influence of calcium content of tissue on hyperhydricity and shoot tip necrosis of in vitro regenerated shoots of Lavandula angustifolia Mill. Braz. Arch. Biol. Technol. 2014, 57, 636–643. [Google Scholar] [CrossRef] [Green Version]
- Spring, O. Chemotoxonomy based on metabolites from glandular trichomes. Adv. Bot. Res. 2000, 31, 153–174. [Google Scholar]
- Naidoo, G.; Kaliamoorthy, S.; Naidoo, Y. The secretory apparatus of Xerophyta viscosa (Velloziaceaea): Epidermis anatomy and chemical composition of the secretory product. Flora 2009, 204, 561–568. [Google Scholar] [CrossRef]
- Manetas, Y. The importance of being hairy: The adverse effects of hair removal on stem photosynthesis of Verbascum speciosum are due to solar UV-B radiation. New Phytol. 2003, 158, 503–508. [Google Scholar] [CrossRef]
- Yang, F.; Miao, L.F. Adaptive responses to progressive drought stress in two poplar species originating from different altitudes. Silva Fenn. 2010, 44, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Osório, M.L.; Gonçalves, S.; Coelho, N.; Osório, J.; Romano, A. Morphological, physiological and oxidative stress markers during acclimatization and field transfer of micropropagated Tuberaria major plants. Plant Cell Tiss. Org. Cult. 2013, 115, 85–97. [Google Scholar] [CrossRef]
- Lodha, D.; Patel, A.K.; Shekhawat, N.S. A high frequency in vitro multiplication, morphological studies and ex vitro rooting of Cadaba fruticose (L.) Druce (Bahuguni): A multipurpose endangered medicinal shrub. Physiol. Mol. Biol. Plants 2015, 21, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Traw, M.B.; Bergelson, J. Interactive effects of jasmonic acid, salicylic acid, and gibberellin on induction of trichomes in Arabidopsis. Plant Physiol. 2003, 133, 1367–1375. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.D.; Lee, B.S.; Kim, T.S.; Choi, Y.E. Developmental plasticity of glandular trichomes into somatic embryogenesis in Tilia Amurensis. Ann. Bot. 2007, 100, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Shekhawat, M.S.; Manokari, M. vitro propagation, micromorphological studies and ex vitro rooting of cannon ball tree (Couroupita guianensis Aubl.): A multipurpose threatened species. Physiol. Mol. Biol. Plants 2016, 22, 131–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kull, U.; Herbig, A. Das Blattadersystem der Angiospermen: Form und Evolution. Naturwissenschaften 1995, 82, 441–451. [Google Scholar] [CrossRef]
- Roth-Nebelsick, A.; Uhl, D.; Mosbrugger, V.; Kerp, H. Evolution and function of leaf venation architecture: A review. Ann. Bot. 2001, 87, 553–566. [Google Scholar] [CrossRef]
- Rathore, M.S.; Rathore, M.S.; Shekhawat, N.S. Ex vivo implications of phyto-hormones on various in vitro responses in Leptadenia reticulata (Retz.) Wight & Arn—An endangered plant. Environ. Exp. Bot. 2013, 86, 86–93. [Google Scholar]
- Fukaki, H.; Tasaka, M. Hormone interactions during lateral root formation. Plant Mol. Biol. 2009, 69, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Rufai, S.; Hanafi, M.M.; Rafii, M.Y.; Mohidin, H.; Omar, S.R.S. Growth and development of Moringa (Moringa oleifera L.) stem cuttings as affected by diameter magnitude, growth media and indole-3-butyric acid. Ann. Res. 2016, 59, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Sharma, U.; Kataria, V.; Shekhawat, N.S. vitro propagation, ex vitro rooting and leaf micromorphology of Bauhinia racemosa lam.: A leguminous tree with medicinal values. Physiol. Mol. Biol. Plants 2017, 23, 969–977. [Google Scholar] [CrossRef]
- Purohit, S.D.; Singhvi, A. Micropropagation of Achras sapota through enhanced axillary branching. Sci. Hort. 1998, 76, 219–229. [Google Scholar] [CrossRef]
- Ochatt, S.J.; Mousset-Déclas, C.; Rancillac, M. Fertile pea plants regenerate from protoplasts when calluses have not undergone endoreduplication. Plant Sci. 2000, 156, 177–183. [Google Scholar] [CrossRef]
- Nassour, M.; Chassériaux, G.; Dorion, N. Optimization of protoplast-to-plant system for Pelargonium × hortorum ‘Alain’ and genetic stability of the regenerated plants. Plant Sci. 2003, 165, 121–128. [Google Scholar] [CrossRef]
- LoSchiavo, F.; Pitto, L.; Giuliano, G.G.; Torti, V.; Nuti-Ronchi, D.; Marazziti, R.; Vergara, S.; Orselli, M.T. DNA methylation of embryogenic carrot cell cultures and its variations as caused by mutation, differentiation, hormones and hypomethylating drugs. Theor. Appl. Genet. 1989, 77, 325–331. [Google Scholar] [CrossRef]
- Leljak-Levanić, D.; Bauer, N.; Mihaljević, S.; Jelaska, S. Changes in DNA methylation during somatic embryogenesis in Cucurbita pepo L. Plant Cell Rep. 2004, 23, 120–127. [Google Scholar] [CrossRef]
- Fraga, H.P.F.; Vieira, L.N.; Caprestano, C.A.; Steinmacher, D.A.; Micke, G.A.; Spudeit, D.A.; Pescador, R.; Guerra, M.P. 5-Azacytidine combined with 2,4-D improves somatic embryogenesis of Acca sellowiana (O. Berg) Burret by means of changes in global DNA methylation levels. Plant Cell Rep. 2012, 31, 2165–2176. [Google Scholar] [CrossRef]
- Fehér, A.; Pasternak, T.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tiss. Org. Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Martin, K.P.; Pachathundikandi, S.; Zhang, C.L.; Slater, A.; Madassery, J. RAPD analysis of a variant of banana (Musa sp.) cv. grandenaine and its propagation via shoot tip culture. Vitr. Cell Dev. Biol. Plant 2006, 42, 188–192. [Google Scholar] [CrossRef]
- Venkatachalam, L.; Sreedhar, R.V.; Bhagyalakshmi, N. Molecular analysis of genetic stability in long-term micropropagated shoots of banana using RAPD and ISSR markers. Elect. J. Biotech. 2007, 15, 106–113. [Google Scholar]
- Jin, W.; Wang, Y.; Wang, H. Adventitious shoot regeneration from leaves of apple rootstock ‘Pingyitiancha’ (Malus hupehensis var. pinyiensis) and genetic fidelity of regenerated plantlets using SSR markers. Can. J. Plant Sci. 2014, 94, 1345–1354. [Google Scholar] [CrossRef] [Green Version]
- Osena, G.; Amugune, N.O.; Nyaboga, E.N. Genetic stability of Cassava plants regenerated through organogenesis using microsatellite markers. J. Plant Sci. 2017, 5, 19–28. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Growth Regulator (mg/L) | No. of Shoots Per Explant | Shoot Height (mm) | No. of Roots Per Explant | Shoot Regeneration (%) | Root Length (mm) | ||
|---|---|---|---|---|---|---|---|
| NAA | BA | TDZ | |||||
| 0.0 | 0.0 | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | |
| 0.0 | 0.5 | 7.50 ± 0.90 e | 12.30 ± 1.00 e | 0.00 a | 70.10 ± 1.10 f | 0.00 a | |
| 0.1 | 0.1 | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | |
| 0.1 | 1.0 | 3.01 ± 0.52 b,c | 1.60 ± 0.20 b | 0.00 a | 36.71 ± 2.50 e | 0.00 a | |
| 0.1 | 2.0 | 3.80 ± 0.10 c,d | 3.77 ± 0.10 d | 0.00 a | 20.10 ± 1.00 b | 0.00 a | |
| 1.0 | 0.1 | 0.00 a | 0.00 a | 2.01 ± 0.50 c | 0.00 a | 3.43 ± 0.20 c | |
| 1.0 | 1.0 | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | |
| 1.0 | 2.0 | 0.00 a | 0.00 a | 4.77 ± 0.60 d | 0.00 a | 5.70 ± 0.05 d | |
| 2.0 | 0.1 | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | |
| 2.0 | 1.0 | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | |
| 2.0 | 2.0 | 3.10 ± 0.50 b–d | 1.10 ± 0.10 b | 3.34 ± 0.20 c | 20.31 ± 1.00 d | 1.77 ± 0.10 b | |
| 0.0 | 0.5 | 8.50 ± 0.90 e | 13.30 ± 1.00 f | 0.00 a | 80.10 ± 1.50 g | 0.00 a | |
| 0.1 | 0.1 | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | |
| 0.1 | 1.0 | 4.38 ± 0.30 d | 2.67 ± 0.10 c | 0.00 a | 30.10 ± 2.50 d | 0.00 a | |
| 0.1 | 2.0 | 2.10 ± 0.10 b | 2.60 ± 0.40 c | 0.00 a | 18.10 ± 0.60 b | 0.00 a | |
| Medium | No. of Shoot Per Explant | Average Shoot Length (mm) |
|---|---|---|
| MS | 8.67 ± 0.94 b | 23.33 ± 1.05 b |
| B5 | 0 a | 0 a |
| SH | 0 a | 0 a |
| Carbohydrate Source | No. of Shoot Per Explant | Average Shoot Length (mm) |
|---|---|---|
| Sucrose (3%) | 8.33 ± 1.10 c | 22.67 ± 1.69 c |
| Fructose (3%) | 1.33 ± 0.47 a | 1.67 ± 0.40 a |
| Maltose (3%) | 5.00 ± 0.82 b | 16.67 ± 2.49 b |
| Medium | Average Root no. Per Explant | Average Root Length (mm) | Average Shoot Length (mm) | Callus |
|---|---|---|---|---|
| MS | 4.11 ± 0.50 a,b | 8.89 ± 1.10 b,c | 8.00 ± 0.80 b,c | No |
| 1/2MS | 7.22 ± 1.10 cd | 59.67 ± 2.10 f | 51.00 ± 3.10 e | No |
| 1/3MS | 2.78 ± 1.05 a | 14.11 ± 1.60 e | 7.33 ± 1.05 b,c | No |
| 1/4MS | 5.72± 1.00 b,c | 6.68 ± 1.10 a | 5.00 ± 1.60 a | No |
| MS + 1 mg/L IBA | 8.44 ± 0.50 d | 10.67 ± 1.60 c | 6.67 ± 1.00 b | Yes |
| MS + 1 mg/L IAA | 13.33 ± 0.15 e | 8.67 ± 2.00 b | 13.50 ± 1.05 d | No |
| MS + 1 mg/L NAA | 2.33 ± 0.40 a | 14.00 ± 1.00 e | 8.83 ± 1.00 c | Yes |
| Plants | Length (μm) | Breadth (μm) | Total Number * |
|---|---|---|---|
| Field grown plants | 15.00 ± 0.82 b | 10.00 ± 0.83 b | 35.00 ± 1.56 b |
| Regenerated plant | 14.00 ± 1.05 a | 9.00 ± 0.47 a | 33.33 ± 2.49 a |
| Plants | Density of Trichome | Density of Islet Termination * |
|---|---|---|
| Field grown plants | 24.00 ± 1.56 a | 45.00 ± 2.90 b |
| Regenerated plant | 25.00 ± 1.74 b | 43.33 ± 2.40 a |
| ISSR Primer | Base Sequence (5′-3′) |
|---|---|
| ISSR1 | (CTG)7G |
| ISSR2 | (GAG)6C |
| ISSR3 | (GAC)6T |
| ISSR4 | (GACA)5 |
| ISSR5 | (GTC)6A |
| ISSR6 | (GTG)6C |
| ISSR7 | (CTC)7 |
| ISSR8 | (GTG)6A |
| ISSR9 | A(CACA)3CACTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, W.-S.; Chung, I.-M.; Kim, S.-H.; Chi, H.-Y.; Yu, C.Y.; Ghimire, B.K. Direct Shoot Organogenesis from Lycium chinense Miller Leaf Explants and Assessment of Genetic Stability Using ISSR Markers. Agronomy 2021, 11, 503. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030503
Jung W-S, Chung I-M, Kim S-H, Chi H-Y, Yu CY, Ghimire BK. Direct Shoot Organogenesis from Lycium chinense Miller Leaf Explants and Assessment of Genetic Stability Using ISSR Markers. Agronomy. 2021; 11(3):503. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030503
Chicago/Turabian StyleJung, Woo-Suk, Ill-Min Chung, Seung-Hyun Kim, Hee-Yeon Chi, Chang Yeon Yu, and Bimal Kumar Ghimire. 2021. "Direct Shoot Organogenesis from Lycium chinense Miller Leaf Explants and Assessment of Genetic Stability Using ISSR Markers" Agronomy 11, no. 3: 503. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030503