
Brassinosteroid Signaling Pathways Interplaying with Diverse Signaling Cues for Crop Enhancement


1
Department of Industrial Plant Science & Technology, College of Agriculture, Life & Environmental Sciences, Chungbuk National University, Cheongju 28644, Korea
2
Department Biology, College of Natural Sciences, Chungbuk National University, Cheongju 28644, Korea
*
Authors to whom correspondence should be addressed.
Agronomy 2021, 11(3), 556; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030556
Submission received: 15 February 2021
/
Revised: 11 March 2021
/
Accepted: 11 March 2021
/
Published: 15 March 2021
(This article belongs to the Special Issue Rice Genetics: Trends and Challenges for the Future Crops Production)
{kind=link}
{kind=link}
Abstract
:The signaling pathways of brassinosteroids (BRs), a unique plant steroid hormone, are critically involved in a diverse range of plant growth and developmental processes as well as many important agronomic traits. Recent advances in the understanding of BR biosynthetic and signaling pathways in model organisms and crops have increased the feasibility of modulating BR responses in crop plants to enhance adaptation to various vulnerable environmental changes. In particular, the identification and functional analysis of BR signaling components in rice (Oryza sativa) present the possibility of their utilization to improve many agricultural traits involved in crop yields. In this review, we summarize recent advances and progress in the understanding of the BR signaling pathway and its interactions with diverse internal and external signaling cues. We also discuss how these physiological modulations of BR and the abundant signaling crosstalk can be applied to enhance rice productivity through the manipulation of plant architecture and fine-tuning of stress responses. Finally, we discuss how the complex regulation of BR signaling pathways could favor application in the molecular design of plant growth and development, precise breeding strategies, and cultivation methods for rice crop improvement.
1. Introduction
Brassinosteroids (BR) regulate a diverse spectrum of processes in plant growth and development as well as internal modulation in response to environmental fluctuation. Since they are essential in plant adaptation and seed plant evolution, the biosynthesis and signaling pathways have been identified in the model plant Arabidopsis, providing a comprehensive understanding of how BR synthesis is controlled and how the signaling pathways are coordinated during the plant’s life cycle [1,2,3,4,5,6]. Moreover, BR controls a number of important agronomic traits including plant height, grain size, tillering, leaf angle, and environmental adaptations, exploiting the potential of the BR response controlling genes as an interesting target by which to enhance crop performance [7,8]. In this review, we will focus on (1) the BR signaling pathway, (2) BR control of growth and development, (3) BR action on environmental adaptation in both Arabidopsis and rice as model systems for dicot and monocot plants, respectively, and (4) the potential precise modulation of BR responses in order to enhance crop breeding.
2. BR Signaling Pathway in Dicot and Monocot Model System
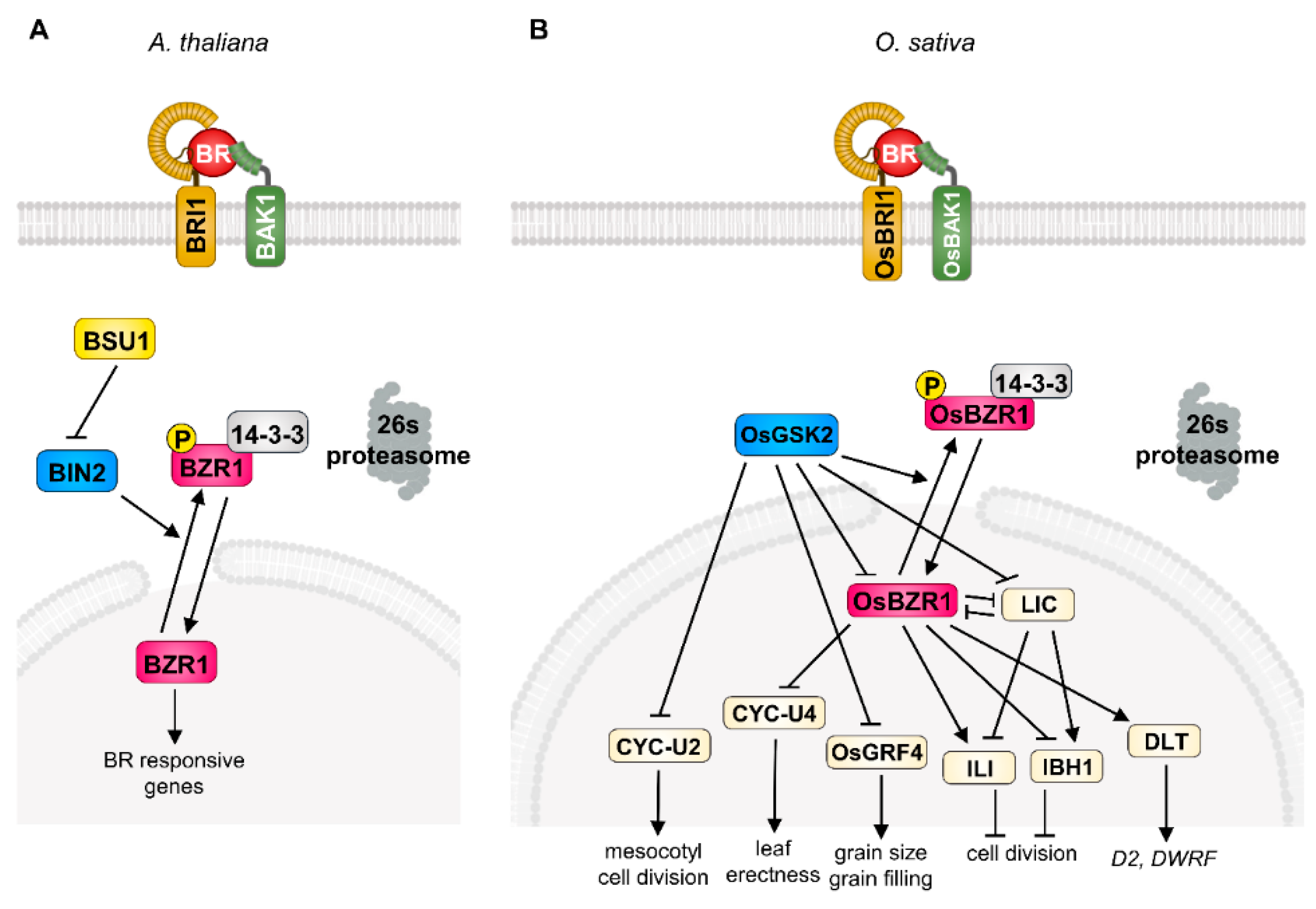
Brassinosteroids were originally isolated from rape (Brassica napus) pollen in 1979 [9]. After identification of BR as a plant-specific steroid hormone, extensive genetic and biochemical studies revealed its signal transduction mechanism in Arabidopsis. In detail, the initiation of BR signaling is tightly mediated by a receptor-like kinase, BRASSINOSTEROID INSENSITIVE 1 (BRI1), and co-receptor kinase, BRI1-ASSOCIATED KINASE 1 (BAK1), at the plasma membrane [1,4]. BR perception through these receptor complexes triggers the dissociation of a negative regulator, BRI1 KINASE INHIBITOR 1 (BKI1), and confers a transphosphorylation of BRI1 and BAK1, leading to the activation of BRI1 SUPPRESSOR 1 (BSU1) and consequent inactivation of BRASSINOSTEROID INSENSITIVE 2 (BIN2) kinase, a representative of the plant GLYCOGEN SYNTHASE KINASE 3 (GSK3) [1,2,3,4,10]. The BIN2 strictly regulates the phosphorylation status of plant-specific transcription factors, BRASSINAZOLE-RESISTANT 1 (BZR1) and BR-INSENSITIVE-EMS-SUPPRESSOR 1/BRASSINAZOLE-RESISTANT 2 (BES1/BZR2), which play critical roles in BR perception downstream events via specific binding to the cis-element in the promoter region of large target genes [5,6,11]. In the absence of BR, BIN2 is activated by auto-phosphorylation and directly phosphorylates BZR1 and BES1, leading to cytosolic accumulation through 14-3-3 binding and degradation by 26S-proteasome [12,13,14].
In the case of the BR signaling pathway in rice, the physiological role of BR and potential BR signaling components have been investigated since the 1980s. Among the diverse primary BR responses (plant height, grain size, lamina bending, grain filling, stress resistance) in rice, the degree of the lamina joint angle serves as a model system by which to determine BR response and sensitivity and contributes to the extensive identification of BR biosynthesis and/or signaling components in rice [15,16]. As a result of intense screening using lamina joint angle and plant height, the primary BR signaling components, the orthologue of Arabidopsis BRI1 in rice (OsBRI1), was first characterized as a functional BR receptor through forward genetic screening as a typical BR-defective phenotype such as erect leaf and dwarfism. Among the various alleles of the OsBRI1 gene, the d61-4 mutant, which carries a null mutation of the BR receptor, shows severely reduced second-node elongation and lamina joint angle but has a mild effect on the plant’s fertility. The second component of BR signaling, OsBAK1, also characterized as an orthologue of Arabidopsis BAK1, functions as a BR co-receptor with BRI1 as evidenced by its loss-of-function mutation, and it displays an erect leaf phenotype and BR insensitivity [17]. In addition, ectopic expression of OsBRI1 or OsBAK1 complements the BR-defective phenotype of d61, supporting the notion that OsBRI1 and OsBAK1 function as BR co-receptors in rice [18]. After the perception of BR in the plasma membrane in rice, OsBRI1/OsBAK1 transduces the phosphorylation cascade and consequently inactivates GLYCOGEN SYNTHASE KINASE (GSK), OsGSK1, and OsGSK2 [19,20]. These Arabidopsis BIN2 counterparts in rice mainly suppress the BR response through direct phosphorylation of rice BR transcription factors. The loss-of-function mutation of OsGSK1/2 results in increased BR sensitivity to lamina inclination and plant height, highlighting the conserved functional role in rice BR signaling [21]. In addition, the rice genome contains four OsBZR1s encoding the rice counterpart of Arabidopsis BZR1, a key transcription factor regulating BR-responsive gene expression. Similar to the phosphorylation-dependent regulation of BZR1 in Arabidopsis, OsBZR1 physically interacts with and is phosphorylated by OsGSKs, which induces the cytoplasmic retention of OsBZR1 via the 14-3-3 protein in rice [22]. OsBZR1 is also controlled by the OsPUB24 ubiquitin ligase through targeted degradation to balance BR signaling homeostasis [23]. Consistent with its positive role in BR signaling, the silencing of OsBZR1 in rice results in semi-dwarfism and decreased lamina joint angle, suggesting that OsBZR1 acts as a downstream BR signaling component in rice.
Interestingly, rice BR signaling involves a diverse range of transcriptional regulators which modulate the primary BR-responsive genes. These include DWARF AND LOW-TILLERING (DLT), LEAF AND TILLER ANGLE INCREASED CONTROLLER (LIC), GRAIN LENGTH 2/GROWTH REGULATING FACTOR 4 (GL2/OsGRF4), OVATE FAMILY PROTEIN (OFP), REDUCED LEAF ANGLE 1/SMALL ORGAN SIZE 1 (RLA1/SMOS1), and U-TYPE CYCLIN (CYC-U), which fine-tune the BR response spatiotemporally via direct and/or indirect interaction with canonical BR signaling components (OsBRI1, OsBAK1, OsGSKs, OsBZR1) in rice (Figure 1). The DLT is a transcriptional regulator that belongs to the GRAS gene family and is phosphorylated by OsGSK2 in a BR-dependent manner. In the presence of BR, the accumulation of the hypo-phosphorylated form of DLT induces several BR responses, such as cell elongation and lamina inclination. Interestingly, BR negatively regulates DLT expression through OsBZR1 activation, but DLT induces OsBZR1 to maintain BR signaling homeostasis [20,24]. The loss-of-function dlt displays the typical BR-deficient phenotype, namely, semi-dwarfism, erect leaves, and reduced tiller number, but overexpression of DLT induces increased BR sensitivity and a physiological BR response. The DLT is also involved in the BR biosynthetic pathway in the context of feedback regulation in controlling the expression of key BR biosynthesis enzymes D2 and DWARF, suggesting that DLT is another positive transcriptional regulator of BR signaling in rice [7,25].
The CCCH-type zinc-finger protein, LIC, is another important transcriptional regulator in rice BR signaling. The silencing and ectopic expression of LIC resulted in elevated and compromised physiological BR responses in rice, respectively. In addition to genetic evidence, the biochemical study of OsGSK2 with LIC also supports its role in the BR signal transduction pathway. The LIC directly interacts with OsGSK2 and is phosphorylated and subsequently accumulated in the cytosol. As a consequence, BR deprivation leads to the loss of the transcriptional activity of LIC and results in enhanced BR sensitivity as a feedback mechanism. Importantly, OsBZR1 directly binds to the promoter of LIC and represses its expression in a BR-dependent manner, indicating the antagonistic relationship between LIC and OsBZR1 in rice BR signaling. Moreover, this relationship seems to be manifested in differential responses to low or high BR concentrations; a low physiological concentration of BR preferentially activates BR signaling via OsBZR1, but a high BR concentration mainly functions to repress the BR response in the LIC-dependent signaling pathway, which potentially aids in rice adaptation via precise BR responses in development and stress resistance [26,27].
The OsGRF4 is another key player controlling BR-responsive gene expression in rice. OsGRF4 was originally characterized as a positive regulator of grain size and filling in rice through grain-size-associated quantitative trait loci (QTL) analysis. Interestingly, OsGSK2 directly interacts with and represses OsGRF4 transcriptional activity under the control of BR perception and miR396 also targets the OsGRF family, indicating that OsGRF4 is specifically involved in grain-related BR responses and is an integrator of diverse hormonal crosstalk in rice [28,29,30,31].
The OFP has been recently identified as a transcription factor that interacts with the DLT protein. In the absence of BR, the OsGSK2 kinase attenuates OFP’s transcriptional activity, whereas BR induces the accumulation of OFP protein in the nucleus and positively regulates the BR response in rice in a DLT-dependent manner [32]. Interestingly, ectopic expression of OFP stimulates gibberellin (GA) inactivation enzyme expression, suggesting that OFP serves as the locus of crosstalk in the context of the BR-induced GA inactivation process in rice [8].
A recent study identified a rice-specific interaction partner of OsBZR1. The APETALA2 DNA-binding transcription factor RLA/SMOS1 forms a complex with OsBZR1 for full activation of BR signal transduction. OsGSK2 also phosphorylates and inactivates, but auxin stimulates RLA/SMOS1 expression, indicating possible crosstalk between BR and auxin signal transduction [33,34].
The CYC-U2 protein, U-type cyclin, is also an important OsGSK2 kinase substrate promoting cell division in the mesocotyl of rice. In the presence of BR, inactivation of OsGSK2 leads to mesocotyl cell division [35]. The RELATED TO ABI3/VP1-LIKE 1 (RAVL1) is another BR signaling component that is required for full activation of BR signaling. The expression level of OsBRI1 is regulated by RAVL1, which directly controls the BR synthesis genes D2, D11, and BRASSINOSTEROID DEFICIENT 1 (BRD1) through binding to the E-box on its target gene’s promoter region [7]. The loss-of-function of RAVL1 is manifested by semi-dwarfism, delayed germination of seedlings, and a dark green phenotype, supporting the notion of its positive effect on rice BR responses [36].
In general, the identification of canonical BR signaling pathway components in rice has greatly improved the understanding of the BR’s functional mechanism for enhancing crop performance. It is noteworthy that OsBZR1 directly/indirectly interacts with a number of TF complexes, such as DLT, LIC, OsGRF4OFP, RLA1/SMOS1, and CYC-U4;1, and these interaction partners are also largely controlled by OsGSKs in an OsBRI1-dependent manner. In the context of different BR levels in specific tissues and stages, differential assembly combination of OsBZR1-interacting transcriptional regulators will generate a diverse series of BR responses or specific BR responses in different tissues/stages during the rice life cycle.
2.1. BR Control of Plant Growth and Development in Dicot and Monocot
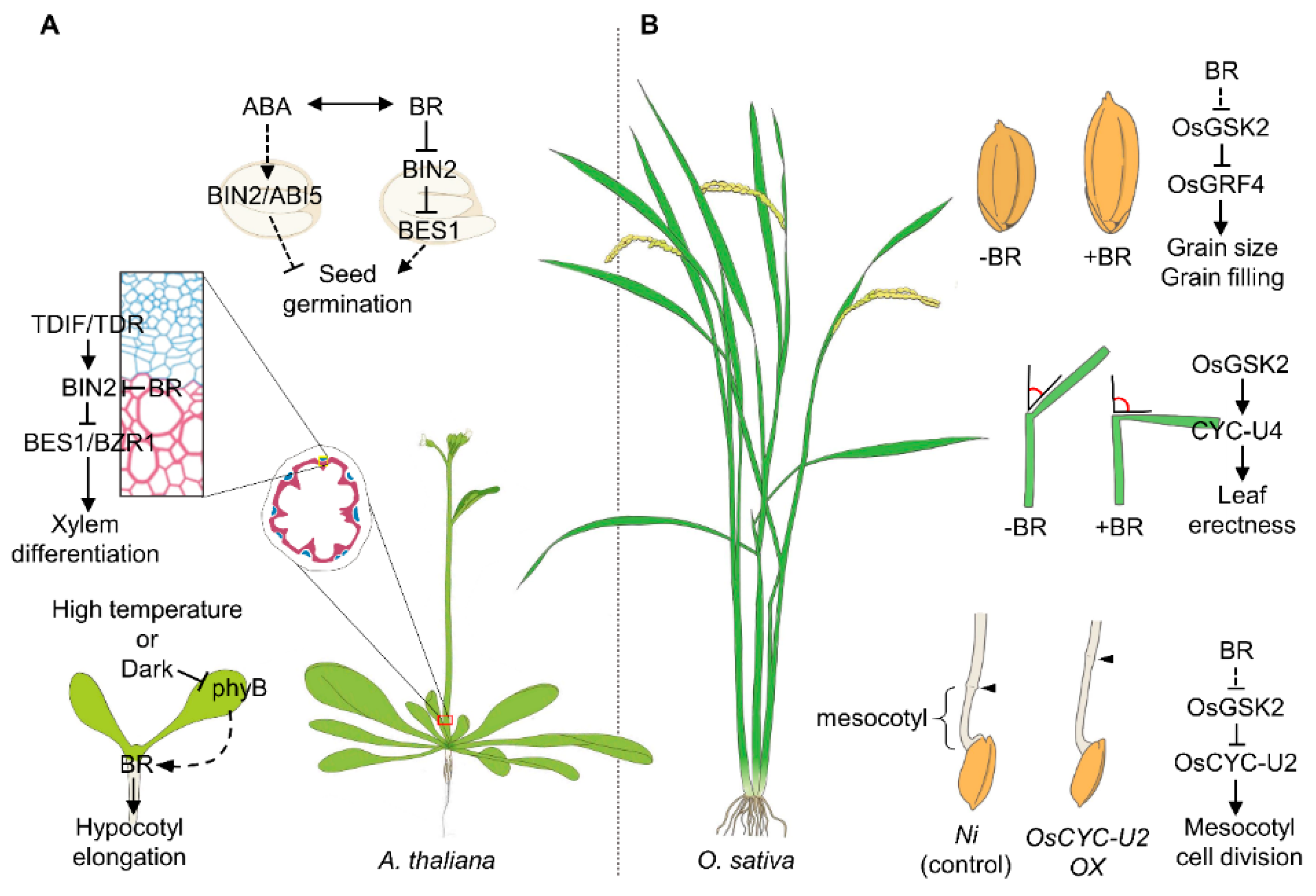
At the beginning of post-embryonic development, germination initiates communication between the environmental conditions and endogenous growth. This process is mainly regulated by a phytohormone, abscisic acid (ABA), that negatively regulates the seed germination in Arabidopsis (Figure 2) [37]. In contrast, BR promotes germination by inhibiting the ABA response. These antagonistic regulatory mechanisms are mediated by BIN2. In the presence of ABA, BIN2 directly phosphorylates and activates the bZIP transcription factor ABSCISIC ACID INSENSITIVE 5 (ABI5) to regulate the expression of ABA-responsive genes. However, BR inactivates BIN2 and leads to the activation of TFs, BZR1/BES1. BR-activated BES1 physically interacts with TOPLESS-HISTONE DEACETYLASE 19 (TPL-HDA19), forming the inhibitory complex. BES1-TPL-HDL19 targets and represses ABI3 and ABI5 by mediating the deacetylation. In conclusion, BR acts as a positive regulator of seed germination [38,39]. BR was found to carry out a key function in the regulation of cambium maintenance and vascular cell differentiation [40]. In the BR signaling cascade, BZR1 and BES1 redundantly promote xylem and phloem cell differentiation. The core signaling pathway, composed of a peptide/receptor (TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR (TDIF)/TDIF RECEPTOR/PHLOEM INTERCALATED WITH XYLEM, (TDR/PXY), and WUSCHEL-RELATED HOMEOBOX 4 (WOX4), mediates self-proliferation and differentiation in close interaction with the BR signaling pathway [41,42,43,44]. The TDR–TDIF signaling directly suppresses BES1 and BZR1 through the activation of the BIN2 kinase in the cambial region, which inhibits and facilitates xylem formation and cambium proliferation, respectively [44].
BR is known to be a key positive hormonal regulator of seed and grain development in both Arabidopsis and rice. In Arabidopsis, the BR biosynthesis-deficient mutant, de-etiolated 2 (det2), shows reduced seed size and fertility [45]. Among a number of genes controlling the seed and embryo/endosperm development, SHORT HYPOCOTYL UNDER BLUE 1 (SHB1), HAIKU 1 (IKU1), HAIKU 2 (IKU2), and MINISEED 3 (MINI3) are known to be the main regulators of seed size and weight through their interactions with canonical BR signaling [46,47,48,49]. These proteins act downstream of BZR1 under the control of the BR-BRI1-BIN2 phosphorylation cascade. On the other hand, BR also inhibits the expression of APETALA 2 (AP2), the floral homeotic gene, and AUXIN RESPONSE FACTOR 2 (ARF2), the key negative regulators of seed size and weight [50,51].
In rice, extensive reverse genetic screens isolated from a number of mutants identified a significant number of BR-related genes in these mutant pools [28,52]. In addition, several key players in BR signaling derived from QTLs support the notion of BR as an essential determinant of grain size, shape, and filling in rice. Among them, the loss-of-function Osbri1 mutant and OsGSK2 overexpression in rice lead to decreased grain size and weight [53]. The seed-size-associated QTL, GL2, which encodes the OsGRF4, also regulates the seed size by inducing the BR response. Antagonistically, GSK2 directly interacts with and inhibits OsGRF4, which results in a reduction in seed development and growth in rice [28]. In addition, GRAIN WIDTH 5 (GW5), a membrane-localized calmodulin-binding protein, regulates the grain width and weight, acting through a promoter of BR signaling [54]. In detail, the GW5-overexpressing plants in the OsGSK2 overexpression line (Go) background show increased grain length and hypersensitivity to exogenous BL treatment, indicating GW5 as a novel BR signaling component involved in grain development. GW5 directly interacts with and suppresses OsGSK2 activity by compromising the auto-phosphorylation of OsGSK2, which is required for its activation. Subsequently, GW5-mediated inactivation of OsGSK2 induces the BR response through the accumulation of the active form of OsBZR1 and DLT in the nucleus. Besides these developmental processes, grain filling contributes to the yield of cereal crops. The organ-specific enhancement of BR synthesis at the stem, leaf, and root, but not at the embryo or endosperm, increases the grain weight and number through increased carbon partitioning to the sink tissue (grain). This process is controlled by the OsBZR1 target, MYB-domain transcription factor CARBON STARVED ANTHER (CSA) [55,56].
The erect leaf and decreased lamina joint angle of rice are key agronomic traits involved in crop enhancement. BR is responsible for increasing the lamina joint angle through the differential elongation of adaxial/abaxial cells in this region. In detail, BR inhibits the protein accumulation and expression level of CYC-U4;1, consequently suppressing cell proliferation in the abaxial side of the lamina joint region through inactivation of OsGSK2 and activation of OsBZR1, respectively [57].
2.2. BR Crosstalk with the Environmental Signal in Dicot and Monocot
In Arabidopsis, BR appears to play the role of central integrator between internal signal and external stress/signal cues. Dark conditions or high air temperatures cause hypocotyl elongation due to integration with endogenous hormonal cues, BR, GA, and auxin. In detail, de-det2, a BR biosynthesis-deficient mutant, displays a defect in the form of dark-induced hypocotyl elongation, whereas bzr1-1D, a mutant that is hypersensitive to BR, suppresses the dark-induced hypocotyl elongation of det2. At the molecular level, BR-activated BZR1 forms a heterodimer with PHYTOCHROME INTERACTING FACTOR 4 (PIF4) and directly regulates the expression of PACLOBUTRAZOLE RESISTANCE (PRE) family genes which are positive regulators of cell elongation [58]. In conclusion, these results suggest the crucial role of BR signaling in cell elongation and crosstalk with the stress response, especially under high-temperature conditions. Under drought stress, ABA stabilizes and activates the transcription factor RESPONSIVE TO DESICCATION 26 (RD26) through BIN2-induced phosphorylation. RD26 regulates the drought-stress-responsive genes and inactivates BES1 to inhibit growth. In contrast, under normal conditions, BR prevents BIN2 activity and BR-activated BES1 downregulates the expression of RD26 [59,60]. Heat stress, which prevents optimal plant growth, accompanies protein denaturation, increased membrane fluidity, and the production of reactive oxygen species (ROS), which all cause photoinhibition and photo-oxidative stress. Photoinhibition- and photo-oxidative-stress-induced overproduction of ROS results in cellular and molecular damage. However, low levels of ROS rather play a role in the plant stress response by acting with phytohormones, such BR, ABA, salicylic acid (SA), and jasmonic acid (JA). Under heat stress, BR upregulates the expression of RESPIRATORY BURST OXIDASE HOMOLOGS (RBOH) at an early stage, which functions as NADPH oxidase and leads to apoplastic accumulation of H2O2 [61]. BR-induced H2O2 triggers the MITOGEN-ACTIVATED PROTEIN KINASE (MAPK) pathway and ABA biosynthesis. In addition, BR primarily mediates the initial stress resistance response, but the prolonged response is regulated by ABA, which is triggered by BR [62]. Meanwhile, in the context of BR signaling, the loss-of-function mutant, bes1, is more sensitive to heat stress compared with WT [63]. In rice, previous studies elucidated the effect of BR in the pathogen-specific function of BR in rice. A brassinolide (BL) treatment experiment showed the broad positive effect of BR for resistance to a broad range of pathogens [64]. To a significant extent, the BL-treated plant rescues the Magnaporthe grisea (M. grisea)-induced lesions on the leaf blade and Xanthomonas oryzae-mediated damage. Moreover, BR induces hypersensitivity to some specific viruses, including the rice black-streaked dwarf virus (RBSDV), which causes a broad range of severe symptoms in rice [65]. Based on transcriptional profiling after RBSDV infection in rice, expression levels of OsDWRF4, OsCPD, OsBRI1, and OsBAK1 are dramatically decreased, while JA signaling-related genes are quickly upregulated. Consistent with this expression pattern, JA-treated rice displays increased resistance to RBSDV and reduced transcript levels of BR-related genes. Interestingly, the transcript levels of BR signaling components show little fluctuation in coi1-13, a loss-of-function mutation of the JA signaling coreceptor, O. sativa CORONATINE INSENSITIVE 1 (OsCOI1), suggesting that the virus-specific effect of BR signaling is suppressed by JA through OsCOI under stress conditions in rice [65]. The BR also regulates the immune response in the root by antagonistic action with SA and GA against the fungal pathogen Pythium graminicola (P. graminicola) [66].
Salinity is one of the major external stresses that severely affects plant development, growth, and seed production. Salt stress mostly impedes the yield of crops by limiting their optimal growth, affecting traits such as source organ size, plant height, grain size, grain weight, and grain quality. Interestingly, among the key salt stress-related hormones in rice, namely, ABA, auxin, and BR, the growth-promoting hormone BR enhances the salt tolerance through the action of OsSERK2, which behaves as a BR signaling co-receptor with OsBRI1 [31]. Interestingly, both the somatic embryogenesis receptor-like kinase 2 (serk2) knockout mutant and SERK2-overexpressing rice show increased grain size, but the SERK2-overexpressing plant displays resistance to salt stress, suggesting a complex interaction network between BR’s promotion of growth and stress tolerance. Moreover, in an analysis of rice T-DNA insertion lines that are associated with abiotic stress, the OsGSK1 was isolated as a negative regulator of BR signaling and a key component conveying potential BR-mediated stress resistance [19].
2.3. BR Application for Increasing Crop Yield in Rice
Recent studies have reported effects on the positive correlation between BR and crop yield increase [67]. Consistently, the physiological effects of BR on the regulation of many important agronomic traits, including the number and weight of grains and planting density, have been widely evaluated, especially in rice. It is well known that enhancing the endogenous BR content by overexpressing its biosynthetic-related genes or exogenous BR application is able to increase grain number and weight [55]. Rice plants overexpressing BR signaling components and Arabidopsis C-22 hydroxylase in the embryos or endosperms displayed BR responses such as enhanced bending of the lamina joint and increased grain size [55,68]. Conversely, many BR biosynthetic- or signaling-related rice mutants showed a dwarf shoot architecture, harboring small seed size and bearing seeds reduced in both length and width. The successful breeding of high-yielding semi-dwarf wheat and rice varieties has contributed significantly to the green revolution of these crops. The semi-dwarf varieties resulted in higher crop yields by enhancing seed biomass production and tolerance to the lodging of rice stems [69]. Many studies reported the critical role of BR in the elongation of the second internode. A BR-insensitive Osbri1 mutant (d61) and BR biosynthetic d2 and d11 mutants displayed dwarf phenotypes [18,25,53,70]. As mentioned above, planting density is also one of the most important agronomic traits for crop yield. The rice varieties with erect leaves enable more dense planting and show higher photosynthetic activity. BR and auxin are well-known hormones that determine the leaf angle of rice [71]. BR-defective rice mutants generally display erect leaf angles with dwarf shoots. RAVL1-mediated regulation of BR homeostasis through the downstream action of BRI1, D2, D11, and BRD1 in rice is critically involved in enlarged leaf angle [25,36]. Upstream BR signaling pathways are integrated into OsBZR1 to directly or indirectly regulate LIC, IBH1, and a rice U-type cyclin, CYC-U4;1, to increase lamella jointing cell elongation [27,72]. Similarly, GA-related d1 mutant (mutated in OsRGA1) also showed a stunted second internode phenotype with erect leaves, suggesting that a specific synergistic interplay between BR and GA takes place in the regulation of rice architecture [73,74,75].
BR signaling outputs are also positively connected with the improvement of grain size and filling. Tissue-specific overexpression of a BR biosynthetic Arabidopsis C-22 hydroxylase gene in grain tissues increased grain weight and grain number in rice. Similarly, the ectopic expression of BR positive regulators O. sativa INCREASE LEAF INCLINATION 1 (OsILI1) and OsILI4 served to enhance both grain size and bending of the lamina joint [68]. Consistently, BR-defective mutants including d61, d11, d2, and brd1 bear small seeds, suggesting the positive role of BR signaling in rice grain filling. Although the BR-defective erect rice mutants showed greater biomass production with higher planting density, the final grain yield did not increase significantly due to the small grain size [53]. However, recent studies have reported alternative approaches to overcome the small-grain phenotype of BR-defective mutants through co-suppression in the shoot by reducing the expression of endogenous OsBRI1. This mutant showed erect leaves and normal seed size, and thus it could be planted at high density. The estimated grain yield of this transformant was almost 30% higher than that of the wild type. A recessive dwarf4-1 mutant reduces bioactive BR content in shoots, resulting in erect leaves but normal reproductive development. Under high-density planting conditions, the grain yield of the dwarf4-1 rice mutant resulted in a nearly 40% enhancement [76]. These findings demonstrate that the yield of rice crop can be increased by generating erect-leaf plants with no defects in grain formation by specifically reducing the BR signaling activity in shoots rather than targeting the reproductive organs. These results collectively indicate that specific BR-related components could offer the possibility to modulate BR responses in order to obtain desirable agronomic traits to improve crop yield and environmental adaptation in rice.
3. Perspective
Currently, BR signaling components in monocots and dicots are undergoing characterization with extensive forward-/reverse genetic screening. Here, we highlighted the current understanding of the signaling network which controls BR signaling in dicot and monocot model systems. The functional characterization of novel BR signaling components in the monocot boosts the breeding program with desirable and customized phenotypes in modern agriculture, such as semi-dwarfism, stress resistance, high yield, and erect leaves. These traits are all strongly associated with BR in cereals and serve as promising targets for enhanced crop yield. Therefore, functional characterization of new components in BR signaling, as well as investigation of their signaling network with endogenous/exogenous signaling cues, are essential fields of research for future crop breeding programs.
Author Contributions
Conceptualization, H.C. and H.R.; writing—original draft preparation, H.C. and H.H.; writing—review and editing, H.C. and H.H.; visualization, H.H.; supervision, H.C.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by The Research Grant of Chungbuk National University in 2019.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Date sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BR | Brassinosteroid |
| BRI1 | BRASSINOSTEROIDS INSENSITIVE 1 |
| BAK1 | BRI1-ASSOCIATED RECEPTOR KINASE 1 |
| BKI1 | BRI1 KINASE INHIBITOR 1 |
| BSU1 | BRI1 SUPPRESSOR 1 |
| BIN2 | BRASSINOSTEROIDS INSENSITIVE 2 |
| GSK3 | GLYCOGEN SYNTHASE KINASE 3 |
| BZR1 | BRASSINAZOLE RESISTANT 1 |
| BES1/BZR2 | BRI1-EMS-SUPPRESSOR 1/BRASSINAZOLE RESISTANT 2 |
| DLT | DWARF AND LOW-TILLERING |
| LIC | LEAF AND TILLER ANGLE INCREASED CONTROLLER |
| GL2/OsGRF4 | GRAIN LENGTH 2/GROWTH REGULATING FACTOR 4 |
| QTL | Quantitative trait loci |
| OFP | OVATE FAMILY PROTEIN |
| GA | Gibberellin |
| RLA/SMOS1 | REDUCED LEAF ANGLE 1/SMALL ORGAN SIZE 1 |
| RAVL1 | RELATED TO ABI 3/VP1-LIKE 1 |
| BRD1 | BRASSINOSTEROID DEFICIENT 1 |
| CYC-U | U-type CYCLIN |
| ABA | Abscisic acid |
| ABI5 | ABSCISIC ACID INSENSITIVE 5 |
| TPL | TOPLESS |
| HDA19 | HISTONE DEACETYLASE 19 |
| TDIF | TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR |
| TDR/PXY | TDIF RECEPTOR/PHLOEM INTERCALATED WITH XYLEM |
| WOX4 | WUSCHEL-RELATED HOMEOBOX 4 |
| DET2 | DE-ETIOLATED 2 |
| SHB1 | SHORT HYPOCOTYL UNDER BLUE 1 |
References
- Li, J.; Chory, J. A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction. Cell 1997, 90, 929–938. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Nam, K.H.J.S. Regulation of brassinosteroid signaling by a GSK3/SHAGGY-like kinase. Science 2002, 295, 1299–1301. [Google Scholar]
- Mora-García, S.; Vert, G.; Yin, Y.; Caño-Delgado, A.; Cheong, H.; Chory, J. Nuclear protein phosphatases with Kelch-repeat domains modulate the response to brassinosteroids in Arabidopsis. Genes Dev. 2004, 18, 448–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, K.H.; Li, J. BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell 2002, 110, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Nakano, T.; Gendron, J.; He, J.; Chen, M.; Vafeados, D.; Yang, Y.; Fujioka, S.; Yoshida, S.; Asami, T. Nuclear-localized BZR1 mediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis. Dev. Cell 2002, 2, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wang, Z.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.; Nomura, T.; Ooka, H.; Ishizaka, M.; Yokota, T.; Sugimoto, K.; Okabe, K.; Kajiwara, H.; Satoh, K.; Yamamoto, K. Isolation and characterization of a rice dwarf mutant with a defect in brassinosteroid biosynthesis. Plant Physiol. 2002, 130, 1152–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Liu, D.; Zhang, G.; Tong, H.; Chu, C. Brassinosteroids regulate OFP1, a DLT interacting protein, to modulate plant architecture and grain morphology in rice. Fornt. Plant Sci. 2017, 8, 1698. [Google Scholar] [CrossRef] [Green Version]
- Grove, M.; Spencer, G.; Rohwedder, W.; Mandava, N.; Worley, J.; Warthen, J.; Steffens, G.; Flippen-Anderson, J.; Cook, J. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature 1979, 281, 216–217. [Google Scholar] [CrossRef]
- Wang, X.; Chory, J. Brassinosteroids regulate dissociation of BKI1, a negative regulator of BRI1 signaling, from the plasma membrane. Science 2006, 313, 1118–1122. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Zola, J.; Aluru, M.; Ye, H.; Foudree, A.; Guo, H.; Anderson, S.; Aluru, S.; Liu, P. A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Arabidopsis thaliana. Plant J. 2011, 65, 634–646. [Google Scholar] [CrossRef]
- Gampala, S.; Kim, T.; He, J.; Tang, W.; Deng, Z.; Bai, M.; Guan, S.; Lalonde, S.; Sun, Y.; Gendron, J. An essential role for 14-3-3 proteins in brassinosteroid signal transduction in Arabidopsis. Dev. Cell 2007, 13, 177–189. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Gendron, J.; Yang, Y.; Li, J.; Wang, Z. The GSK3-like kinase BIN2 phosphorylates and destabilizes BZR1, a positive regulator of the brassinosteroid signaling pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2002, 99, 10185–10190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, H.; Kim, K.; Cho, H.; Park, J.; Choe, S.; Hwang, I. Nucleocytoplasmic shuttling of BZR1 mediated by phosphorylation is essential in Arabidopsis brassinosteroid signaling. Plant Cell 2007, 19, 2749–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, K.; Marumo, S.; Abe, H.; Morishita, T.; Nakamura, K.; Uchiyama, M.; Mori, K.; Chemistry, B. A rice lamina inclination test-a micro-quantitative bioassay for brassinosteroids. Agric. Biol. Chem. 1984, 48, 719–726. [Google Scholar]
- Wada, K.; Marumo, S.; Ikekawa, N.; Morisaki, M.; Mori, K. Brassinolide and homobrassinolide promotion of lamina inclination of rice seedlings. Plant Cell Physiol. 1981, 22, 323–325. [Google Scholar]
- Li, D.; Wang, L.; Wang, M.; Xu, Y.Y.; Luo, W.; Liu, Y.J.; Xu, Z.H.; Li, J.; Chong, K. Engineering OsBAK1 gene as a molecular tool to improve rice architecture for high yield. Plant Biotechnol. J. 2009, 7, 791–806. [Google Scholar] [CrossRef]
- Yamamuro, C.; Ihara, Y.; Wu, X.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. Plant Cell 2000, 12, 1591–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, S.; Lee, S.; Kim, M.; Koh, J.; Lee, S.; An, G.; Choe, S.; Kim, S. T-DNA tagged knockout mutation of rice OsGSK1, an orthologue of Arabidopsis BIN2, with enhanced tolerance to various abiotic stresses. Plant Mol. Biol. 2007, 65, 453–466. [Google Scholar] [CrossRef]
- Tong, H.; Liu, L.; Jin, Y.; Du, L.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. Dwarf and Low-Tillering acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice. Plant Cell 2012, 24, 2562–2577. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, J.; Zhang, X.; Zhou, J.; Jiang, Z.; Huang, P.; Tang, Z.; Bao, Y.; Cheng, J.; Tang, H. Rice qGL3/OsPPKL1 functions with the GSK3/SHAGGY-like kinase OsGSK3 to modulate brassinosteroid signaling. Plant Cell. 2019, 31, 1077–1093. [Google Scholar] [CrossRef]
- Bai, M.; Zhang, L.; Gampala, S.; Zhu, S.; Song, W.; Chong, K.; Wang, Z. Functions of OsBZR1 and 14-3-3 proteins in brassinosteroid signaling in rice. Proc. Natl. Acad. Sci. USA 2007, 104, 13839–13844. [Google Scholar] [CrossRef] [Green Version]
- Min, H.; Cui, L.; Oh, T.; Kim, J.; Kim, T.; Kim, W. Os BZR 1 turnover mediated by Os SK 22-regulated U-box E3 ligase Os PUB 24 in rice BR response. Plant J. 2019, 99, 426–438. [Google Scholar] [CrossRef]
- Tong, H.; Jin, Y.; Liu, W.; Li, F.; Fang, J.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. DWARF AND LOW-TILLERING, a new member of the GRAS family, plays positive roles in brassinosteroid signaling in rice. Plant J. 2009, 58, 803–816. [Google Scholar] [CrossRef]
- Hong, Z.; Ueguchi-Tanaka, M.; Shimizu-Sato, S.; Inukai, Y.; Fujioka, S.; Shimada, Y.; Takatsuto, S.; Agetsuma, M.; Yoshida, S.; Watanabe, Y. Loss-of-function of a rice brassinosteroid biosynthetic enzyme, C-6 oxidase, prevents the organized arrangement and polar elongation of cells in the leaves and stem. Plant J. 2002, 32, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, Y.; Zhang, C.; Ma, Q.; Joo, S.; Kim, S.; Xu, Z.; Chong, K. OsLIC, a novel CCCH-type zinc finger protein with transcription activation, mediates rice architecture via brassinosteroids signaling. PLoS ONE 2008, 3, e3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Xu, Y.; Guo, S.; Zhu, J.; Huan, Q.; Liu, H.; Wang, L.; Luo, G.; Wang, X.; Chong, K. Dynamics of brassinosteroid response modulated by negative regulator LIC in rice. PLoS Genet. 2012, 8, e1002686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, R.; Tong, H.; Shi, B.; Liu, Y.; Fang, S.; Liu, D.; Xiao, Y.; Hu, B.; Liu, L.; Wang, H. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nat. Plants 2015, 2, 1–8. [Google Scholar]
- Duan, P.; Ni, S.; Wang, J.; Zhang, B.; Xu, R.; Wang, Y.; Chen, H.; Zhu, X.; Li, Y. Regulation of OsGRF4 by OsmiR396 controls grain size and yield in rice. Nat. Plants 2015, 2, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Y.; Fang, Y.; Zeng, L.; Xu, J.; Yu, H.; Shi, Z.; Pan, J.; Zhang, D.; Kang, S. A rare allele of GS2 enhances grain size and grain yield in rice. Mol. Plant 2015, 8, 1455–1465. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Liu, H.; Guo, S.; Wang, B.; Li, Z.; Chong, K.; Xu, Y. OsmiR396d affects gibberellin and brassinosteroid signaling to regulate plant architecture in rice. Plant Physiol. 2018, 176, 946–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, N.; Yin, W.; Liu, D.; Zhang, X.; Yu, Z.; Huang, W.; Liu, J.; Yang, Y.; Meng, W.; Niu, M. Regulation of Brassinosteroid Signaling and Salt Resistance by SERK2 and Potential Utilization for Crop Improvement in Rice. Front. Plant Sci. 2020, 11, 621859. [Google Scholar] [CrossRef]
- Aya, K.; Hobo, T.; Sato-Izawa, K.; Ueguchi-Tanaka, M.; Kitano, H.; Matsuoka, M. A novel AP2-type transcription factor, SMALL ORGAN SIZE1, controls organ size downstream of an auxin signaling pathway. Plant Cell Physiol. 2014, 55, 897–912. [Google Scholar] [CrossRef] [Green Version]
- Hirano, K.; Yoshida, H.; Aya, K.; Kawamura, M.; Hayashi, M.; Hobo, T.; Sato-Izawa, K.; Kitano, H.; Ueguchi-Tanaka, M.; Matsuoka, M. Small organ size 1 and small organ size 2/dwarf and low-tillering form a complex to integrate auxin and brassinosteroid signaling in rice. Mol. Plant 2017, 10, 590–604. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Wang, T.; Wang, L.; Li, X.; Jia, Y.; Liu, C.; Huang, X.; Xie, W.; Wang, X. Natural selection of a GSK3 determines rice mesocotyl domestication by coordinating strigolactone and brassinosteroid signaling. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Je, B.; Piao, H.; Park, S.; Park, S.; Kim, C.; Xuan, Y.; Park, S.; Huang, J.; Choi, Y.; An, G. RAV-Like1 Maintains Brassinosteroid Homeostasis via the Coordinated Activation of BRI1 and Biosynthetic Genes in Rice. Plant Cell 2010, 22, 1777–1791. [Google Scholar] [CrossRef] [Green Version]
- Steber, C.M.; McCourt, P. A Role for Brassinosteroids in Germination in Arabidopsis. Plant Physiol. 2001, 125, 763–769. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Yu, D. BRASSINOSTEROID INSENSITIVE2 Interacts with ABSCISIC ACID INSENSITIVE5 to Mediate the Antagonism of Brassinosteroids to Abscisic Acid During Seed Germination in Arabidopsis. Plant Cell 2014, 26, 4394–4408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, H.; Cho, H.; Bae, W.; Hwang, I. Control of Early Seedling Development by BES1/TPL/HDA19-mediated Epigenetic Regulation of ABI3. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Saito, M.; Kondo, Y.; Fukuda, H. BES1 and BZR1 Redundantly Promote Phloem and Xylem Differentiation. Plant Cell Physiol. 2018, 59, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Fisher, K.; Turner, S. PXY, a Receptor-like Kinase Essential for Maintaining Polarity During Plant Vascular-tissue Development. Curr. Biol. 2007, 17, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, Y.; Shinohara, H.; Kondo, Y.; Inoue, A.; Nakanomyo, I.; Ogawa, M.; Sawa, S.; Ohashi-Ito, K.; Matsubayashi, Y.; Fukuda, H. Non-cell-autonomous Control of Vascular Stem Cell Fate by a CLE Peptide/Receptor System. Proc. Natl. Acad. Sci. USA 2008, 105, 15208–15213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Strable, J.; Shimizu, R.; Koenig, D.; Sinha, N.; Scanlon, M.J. WOX4 Promotes Procambial Development. Plant Physiol. 2010, 152, 1346–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, Y.; Ito, T.; Nakagami, H.; Hirakawa, Y.; Saito, M.; Tamaki, T.; Shirasu, K.; Fukuda, H. Plant GSK3 Proteins Regulate Xylem Cell Differentiation Downstream of TDIF–TDR Signalling. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Huang, H.; Hu, Y.; Zhu, S.; Wang, Z.; Lin, W. Brassinosteroid Regulates Seed Size and Shape in Arabidopsis. Plant Physiol. 2013, 162, 1965–1977. [Google Scholar] [CrossRef]
- Garcia, D.; Saingery, V.; Chambrier, P.; Mayer, U.; Jürgens, G.; Berger, F. Arabidopsis Haiku Mutants Reveal New Controls of Seed Size by Endosperm. Plant Physiol. 2003, 131, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Dennis, E.; Berger, F.; Peacock, W.; Chaudhury, A. MINISEED3 (MINI3), a WRKY Family Gene, and HAIKU2 (IKU2), a Leucine-rich Repeat (LRR) KINASE Gene, are Regulators of Seed Size in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 17531–17536. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Garcia, D.; Zhang, H.; Feng, K.; Chaudhury, A.; Berger, F.; Peacock, W.; Dennis, E.; Luo, M. The VQ Motif Protein IKU1 Regulates Endosperm Growth and Seed Size in Arabidopsis. Plant J. 2010, 63, 670–679. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, X.; Kang, X.; Zhao, X.; Zhang, X.; Ni, M. SHORT HYPOCOTYL UNDER BLUE1 Associates with MINISEED3 and HAIKU2 Promoters in vivo to Regulate Arabidopsis Seed Development. Plant Cell 2009, 21, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Ohto, M.; Floyd, S.; Fischer, R.; Goldberg, R.; Harada, J. Effects of APETALA2 on Embryo, Endosperm, and Seed Coat Development Determine Seed Size in Arabidopsis. Sex Plant Reprod. 2009, 22, 277–289. [Google Scholar] [CrossRef] [Green Version]
- Schruff, M.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R. The AUXIN RESPONSE FACTOR 2 Gene of Arabidopsis links Auxin Signalling, Cell Division, and the Size of Seeds and Other Organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, J.; Gu, S.; Wan, X.; Gao, H.; Guo, T.; Su, N.; Lei, C.; Zhang, X.; Cheng, Z.; Guo, X. Isolation and Initial Characterization of GW5, a Major QTL Associated with Rice Grain Width and Weight. Cell Res. 2008, 18, 1199–1209. [Google Scholar] [CrossRef]
- Morinaka, Y.; Sakamoto, T.; Inukai, Y.; Agetsuma, M.; Kitano, H.; Ashikari, M.; Matsuoka, M. Morphological Alteration Caused by Brassinosteroid Insensitivity Increases the Biomass and Grain Production of Rice. Plant Physiol. 2006, 141, 924–931. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, J.; Zheng, X.; Wu, F.; Lin, Q.; Heng, Y.; Tian, P.; Cheng, Z.; Yu, X.; Zhou, K. GW5 Acts in the Brassinosteroid Signalling Pathway to Regulate Grain Width and Weight in Rice. Nat. Plants 2017, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Trieu, A.; Radhakrishnan, P.; Kwok, S.; Harris, S.; Zhang, K.; Wang, J.; Wan, J.; Zhai, H.; Takatsuto, S. Brassinosteroids Regulate Grain Filling in Rice. Plant Cell 2008, 20, 2130–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Liang, W.; Cui, X.; Chen, M.; Yin, C.; Luo, Z.; Zhu, J.; Lucas, W.; Wang, Z.; Zhang, D. Brassinosteroids Promote Development of Rice Pollen Grains and Seeds by Triggering Expression of Carbon Starved Anther, a MYB Domain Protein. Plant J. 2015, 82, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Chen, D.; Li, X.; Qiao, S.; Shi, C.; Li, C.; Shen, H.; Wang, X. Brassinosteroid Signaling Regulates Leaf Erectness in Oryza sativa via the Control of a Specific U-type Cyclin and Cell Proliferation. Dev. Cell. 2015, 34, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Zhu, J.; Bai, M.; Arenhart, R.; Sun, Y.; Wang, Z. Cell Elongation is Regulated Through a Central Circuit of Interacting Transcription Factors in the Arabidopsis Hypocotyl. Elife 2014, 3, e03031. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Tang, B.; Xie, Z.; Nolan, T.; Ye, H.; Song, G.; Walley, J.; Yin, Y. GSK 3-like Kinase BIN 2 Phosphorylates RD 26 to Potentiate Drought Signaling in Arabidopsis. Plant J. 2019, 100, 923–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Liu, S.; Tang, B.; Chen, J.; Xie, Z.; Nolan, T.; Jiang, H.; Guo, H.; Lin, H.; Li, L. RD26 Mediates Crosstalk Between Drought and Brassinosteroid Signalling Pathways. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X.; Wang, Y.; Zhou, Y.; Tao, Y.; Mao, W.; Shi, K.; Asami, T.; Chen, Z.; Yu, J. Reactive Oxygen Species are Involved in Brassinosteroid-induced Stress Tolerance in Cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wang, J.; Li, X.; Xia, X.; Zhou, Y.; Shi, K.; Chen, Z.; Yu, J. H2O2 Mediates the Crosstalk of Brassinosteroid and Abscisic Acid in Tomato Responses to Heat and Oxidative Stresses. J. Exp. Bot. 2014, 65, 4371–4383. [Google Scholar] [CrossRef] [PubMed]
- Setsungnern, A.; Muñoz, P.; Pérez-Llorca, M.; Müller, M.; Thiravetyan, P.; Munné-Bosch, S. A Defect in BRI1-EMS-SUPPRESSOR 1 (BES1)-Mediated Brassinosteroid Signaling Increases Photoinhibition and Photo-oxidative Stress during Heat Stress Acclimation in Arabidopsis. Plant Sci. 2020, 296, 110470. [Google Scholar] [CrossRef] [PubMed]
- Nakashita, H.; Yasuda, M.; Nitta, T.; Asami, T.; Fujioka, S.; Arai, Y.; Sekimata, K.; Takatsuto, S.; Yamaguchi, I.; Yoshida, S. Brassinosteroid Functions in a Broad Range of Disease Resistance in Tobacco and Rice. Plant J. 2003, 33, 887–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Zhang, H.; Sun, Z.; Li, J.; Hong, G.; Zhu, Q.; Zhou, X.; MacFarlane, S.; Yan, F.; Chen, J. Jasmonic Acid-Mediated Defense Suppresses Brassinosteroid-mediated Susceptibility to Rice Black Streaked Dwarf Virus Infection in Rice. New Phytol. 2017, 214, 388–399. [Google Scholar] [CrossRef] [Green Version]
- De Vleesschauwer, D.; Van Buyten, E.; Satoh, K.; Balidion, J.; Mauleon, R.; Choi, I.; Vera-Cruz, C.; Kikuchi, S.; Höfte, M. Brassinosteroids Antagonize Gibberellin-and Salicylate-mediated Root Immunity in Rice. Plant Physiol. 2012, 158, 1833–1846. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Chu, C. Functional Specificities of Brassinosteroid and Potential Utilization for Crop Improvement. Trends Plant Sci. 2018, 23, 1016–1028. [Google Scholar] [CrossRef]
- Tanaka, A.; Nakagawa, H.; Tomita, C.; Shimatani, Z.; Ohtake, M.; Nomura, T.; Jiang, C.; Dubouzet, J.; Kikuchi, S.; Sekimoto, H. BRASSINOSTEROID UPREGULATED1, Encoding a Helix-loop-Helix Protein, is a Novel Gene Involved in Brassinosteroid Signaling and Controls Bending of the Lamina Joint in Rice. Plant Physiol. 2009, 151, 669–680. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Matsuoka, M. Generating High-yielding Varieties by Genetic Manipulation of Plant Architecture. Curr. Opin. Biotechnol. 2004, 15, 144–147. [Google Scholar] [CrossRef]
- Tanabe, S.; Ashikari, M.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Yano, M.; Yoshimura, A.; Kitano, H.; Matsuoka, M.; Fujisawa, Y.; et al. A Novel Cytochrome P450 Is Implicated in Brassinosteroid Biosynthesis via the Characterization of a Rice Dwarf Mutant, dwarf11, with Reduced Seed Length. Plant Cell 2005, 17, 776–790. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Fujioka, S.; Takatsuto, S.; Tsujimoto, M.; Kitano, H.; Yoshida, S.; Asami, T.; Nakano, T. Involvement of C-22-hydroxylated Brassinosteroids in Auxin-induced Lamina Joint Bending in Rice. Plant Cell Physiol. 2009, 50, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Bai, M.; Wu, J.; Zhu, J.; Wang, H.; Zhang, Z.; Wang, W.; Sun, Y.; Zhao, J.; Sun, X. Antagonistic HLH/bHLH Transcription Factors Mediate Brassinosteroid Regulation of Cell Elongation and Plant Development in Rice and Arabidopsis. Plant Cell 2009, 21, 3767–3780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashikari, M.; Wu, J.; Yano, M.; Sasaki, T.; Yoshimura, A. Rice Gibberellin-insensitive Dwarf Mutant Gene Dwarf 1 Encodes the α-subunit of GTP-binding Protein. Proc. Natl. Acad. Sci. USA 1999, 96, 10284–10289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, Y.; Kato, T.; Ohki, S.; Ishikawa, A.; Kitano, H.; Sasaki, T.; Asahi, T.; Iwasaki, Y. Suppression of the Heterotrimeric G Protein Causes Abnormal Morphology, Including Dwarfism, in Rice. Proc. Natl. Acad. Sci. USA 1999, 96, 7575–7580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urano, D.; Chen, J.-G.; Botella, J.; Jones, A. Heterotrimeric G Protein Signalling in the Plant Kingdom. Open Biol. 2013, 3, 120186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, T.; Matsuoka, M. Characterization of CONSTITUTIVE PHOTOMORPHOGENESIS AND DWARFISM Homologs in Rice (Oryza sativa L.). J. Plant Growth Regul. 2006, 25, 245–251. [Google Scholar] [CrossRef]
Figure 1.
Brassinosteroid (BR) signaling pathways in Arabidopsis and rice. The simplified BR signaling cascade in Arabidopsis (A) and rice (B). BRI1 directly interacts with BR and forms the receptor protein complex with BAK1 which promotes the phosphorylation of BSU1 indirectly. Phosphorylated BSU1 inactivates the BIN2 and leads to nuclear accumulation of dephosphorylated BZR1 in Arabidopsis (A). The BR-OsBRI1-OsBAK1 complex inactivates the OsGSK2 which inactivates the BR signaling output regulators including OsBZR1, LIC, OsGRF4, and CYC-U2 in rice. De-phosphorylated OsBZR1 regulates the target components (CYC-U4;1, LIC, ILI, IBH1, and DLT) involved in primary BR response in rice (B).
Figure 1.
Brassinosteroid (BR) signaling pathways in Arabidopsis and rice. The simplified BR signaling cascade in Arabidopsis (A) and rice (B). BRI1 directly interacts with BR and forms the receptor protein complex with BAK1 which promotes the phosphorylation of BSU1 indirectly. Phosphorylated BSU1 inactivates the BIN2 and leads to nuclear accumulation of dephosphorylated BZR1 in Arabidopsis (A). The BR-OsBRI1-OsBAK1 complex inactivates the OsGSK2 which inactivates the BR signaling output regulators including OsBZR1, LIC, OsGRF4, and CYC-U2 in rice. De-phosphorylated OsBZR1 regulates the target components (CYC-U4;1, LIC, ILI, IBH1, and DLT) involved in primary BR response in rice (B).

Figure 2.
A schematic diagram of BR-mediated development and growth regulation in plants. BR regulates the development and growth of dicot (A) and monocot (B). BR promotes seed germination through the canonical signaling pathways by inhibiting the abscisic acid (ABA)-mediated seed dormancy. BR signaling enhances the xylem formation during secondary growth. Under the high-temperature condition, thermosensing at cotyledon initiates the BR-induced hypocotyl elongation through intracellular interaction (A). BR signaling enhances the grain size and mesocotyl elongation in rice. The cell division regulation by BR signaling determines the leaf inclination (B).
Figure 2.
A schematic diagram of BR-mediated development and growth regulation in plants. BR regulates the development and growth of dicot (A) and monocot (B). BR promotes seed germination through the canonical signaling pathways by inhibiting the abscisic acid (ABA)-mediated seed dormancy. BR signaling enhances the xylem formation during secondary growth. Under the high-temperature condition, thermosensing at cotyledon initiates the BR-induced hypocotyl elongation through intracellular interaction (A). BR signaling enhances the grain size and mesocotyl elongation in rice. The cell division regulation by BR signaling determines the leaf inclination (B).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hwang, H.; Ryu, H.; Cho, H. Brassinosteroid Signaling Pathways Interplaying with Diverse Signaling Cues for Crop Enhancement. Agronomy 2021, 11, 556. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030556
AMA Style
Hwang H, Ryu H, Cho H. Brassinosteroid Signaling Pathways Interplaying with Diverse Signaling Cues for Crop Enhancement. Agronomy. 2021; 11(3):556. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030556
Chicago/Turabian StyleHwang, Hyeona, Hojin Ryu, and Hyunwoo Cho. 2021. "Brassinosteroid Signaling Pathways Interplaying with Diverse Signaling Cues for Crop Enhancement" Agronomy 11, no. 3: 556. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030556
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.