
Optimization of Agrobacterium Mediated Genetic Transformation in Paspalum scrobiculatum L. (Kodo Millet)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of Explants
2.2. Screening of Potent Varieties
2.3. Antibiotic Sensitivity Test
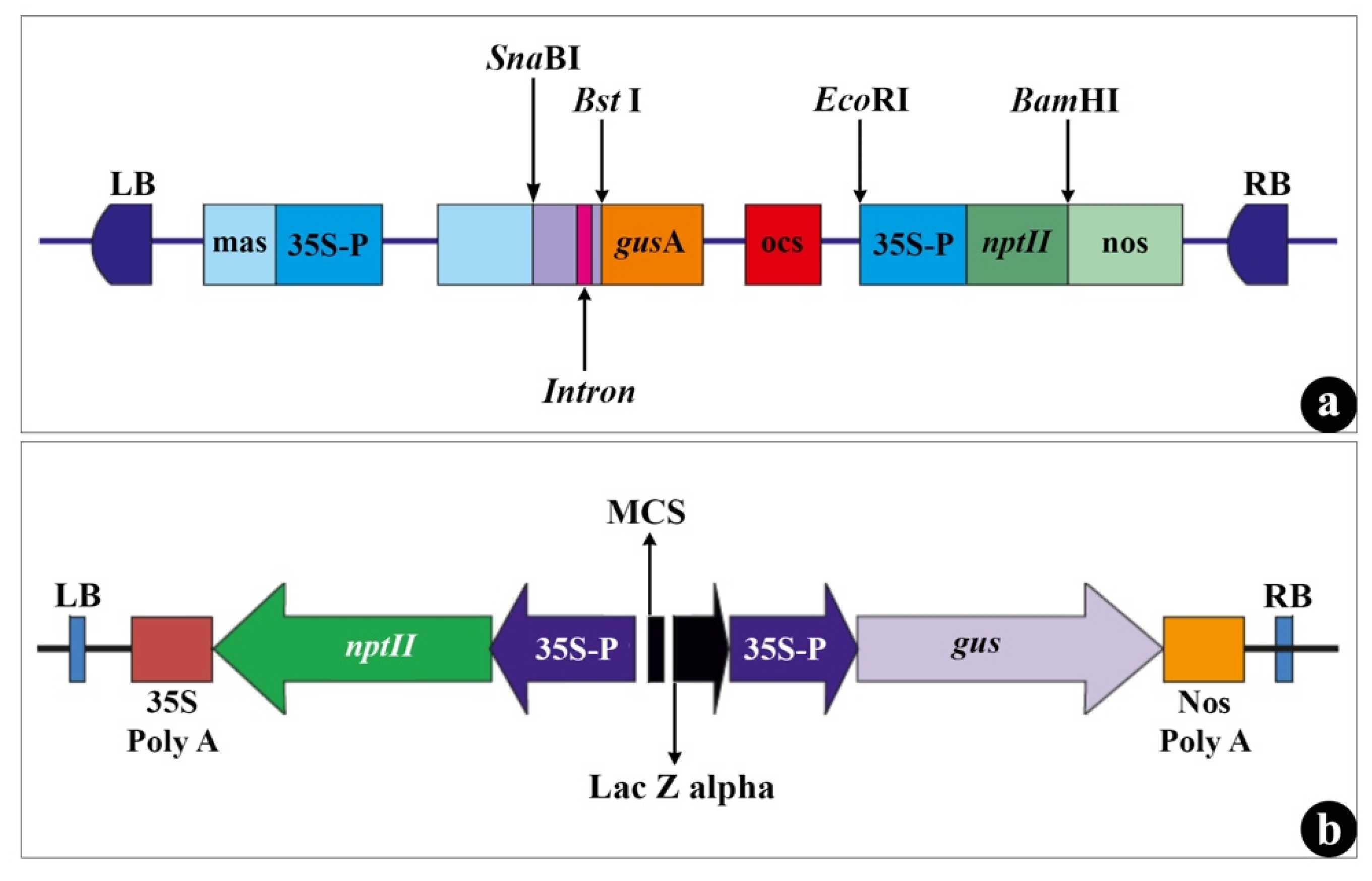
2.4. Bacterial Strain and Inoculum Preparation
2.5. Infection and Co-Cultivation
2.6. Selection and Regeneration of Transformed Plantlets
2.7. GUS Histochemical Assay
3. Result and Discussion
3.1. Selection of Variety
3.2. Antibiotic Sensitivity
3.3. Optimization of Different Parameters for Efficient A. tumefaciens Mediated Transformation
3.3.1. Agrobacterium Strain(s)
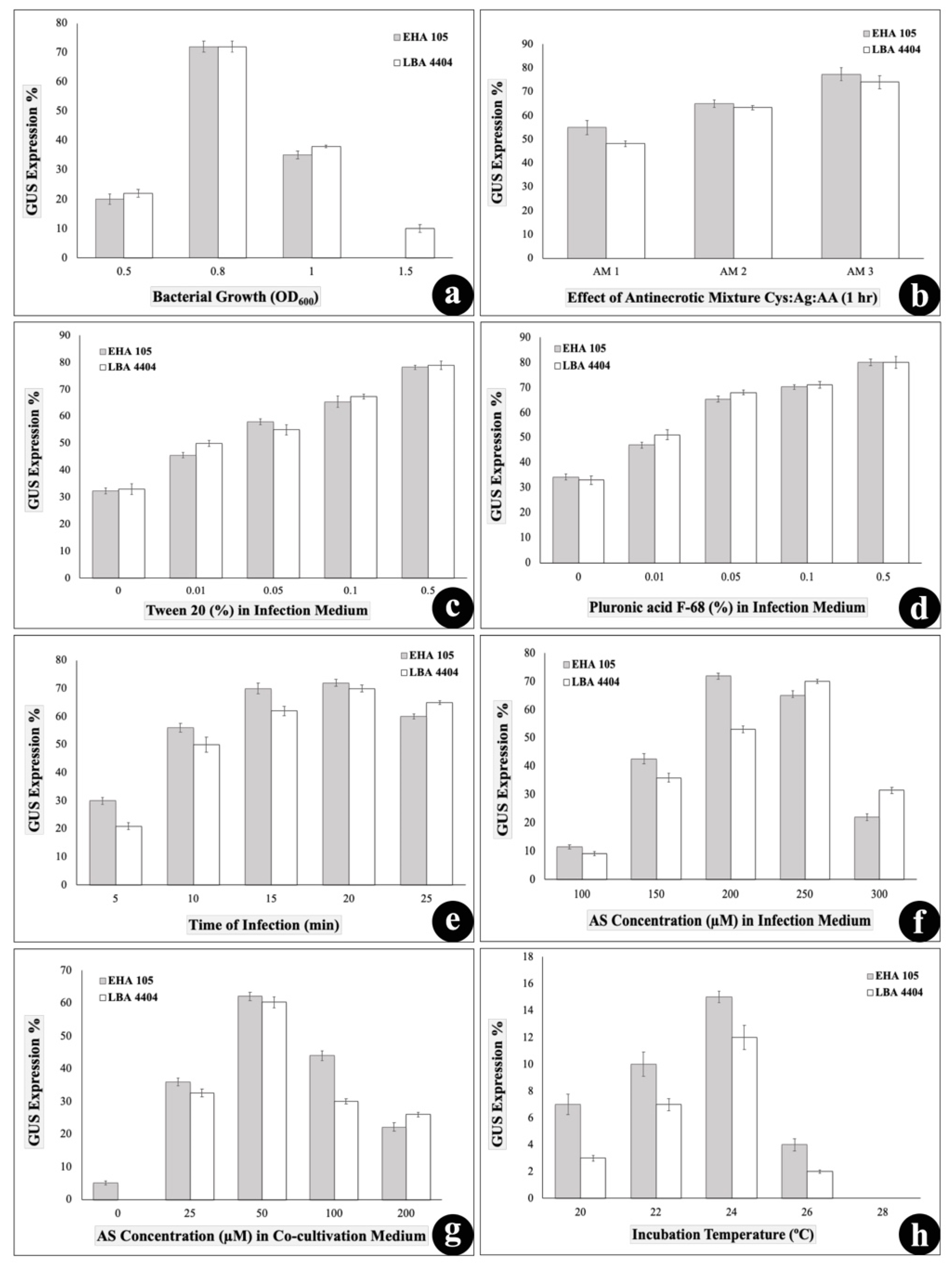
3.3.2. Effect of Cell Density
3.3.3. Effect of Pretreatment with Antinecrotic Mixture
3.3.4. Effect of Surfactants
3.3.5. Effect of Infection Time
3.3.6. Effect of Acetosyringone
3.3.7. Effect of Co-Cultivation Time and Temperature
3.4. Co-Cultivation, Selection and Regeneration
3.5. GUS Histochemical Assay, Transformation Efficiency and Field Transfer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Density (OD600) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| 0.5 | 90 | 90 | 22 | 18 | 5 | 3 | 5.5 | 3.3 |
| 0.8 | 90 | 90 | 35 | 26 | 14 | 10 | 15.5 | 11.1 |
| 1 | 90 | 90 | 20 | 32 | 2 | 13 | 2.2 | 14.4 |
| 1.5 | 90 | 90 | 9 | 5 | 0 | 2 | 0 | 2.22 |
| Antinecrotic Mixture (AM) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| AM 1 | 80 | 80 | 16 | 9 | 6 | 5 | 7.5 | 6.2 |
| AM 2 | 80 | 80 | 22 | 17 | 10 | 9 | 12.5 | 11.2 |
| AM 3 | 80 | 80 | 36 | 32 | 18 | 13 | 22.5 | 16.2 |
| Infection Time (min) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| 5 | 105 | 105 | 2 | 0 | 0 | 0 | 0 | 0 |
| 10 | 105 | 105 | 16 | 12 | 8 | 10 | 7.6 | 9.5 |
| 15 | 105 | 105 | 21 | 18 | 17 | 13 | 16.1 | 12.38 |
| 20 | 105 | 105 | 32 | 28 | 26 | 21 | 24.76 | 20 |
| 25 | 105 | 105 | 6 | 10 | 3 | 2 | 2.8 | 1.9 |
| AS concentration (µM) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| 100 | 90 | 90 | 27 | 19 | 8 | 2 | 8.8 | 2.2 |
| 150 | 90 | 90 | 36 | 25 | 17 | 9 | 18.8 | 10 |
| 200 | 90 | 90 | 42 | 32 | 25 | 18 | 27.7 | 20 |
| 250 | 90 | 90 | 25 | 38 | 14 | 21 | 15.5 | 23.3 |
| 300 | 90 | 90 | 6 | 11 | 0 | 0 | 0 | 0 |
| AS concentration (µM) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| 0 | 90 | 90 | 21 | 15 | 12 | 6 | 13.3 | 6.6 |
| 25 | 90 | 90 | 38 | 32 | 21 | 19 | 23.3 | 21.1 |
| 50 | 90 | 90 | 51 | 43 | 27 | 21 | 30 | 23.3 |
| 100 | 90 | 90 | 22 | 36 | 14 | 17 | 15.5 | 18.8 |
| 200 | 90 | 90 | 11 | 6 | 3 | 1 | 3.3 | 1.1 |
| Tween-20 Conc. (%) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| 0 (Control) | 75 | 75 | 12 | 6 | 4 | 2 | 5.3 | 2.6 |
| 0.01 | 75 | 75 | 26 | 15 | 16 | 9 | 21.3 | 12 |
| 0.05 | 75 | 75 | 33 | 27 | 22 | 18 | 29.3 | 24 |
| 0.1 | 75 | 75 | 38 | 31 | 24 | 21 | 32 | 28 |
| 0.5 | 75 | 75 | 42 | 39 | 32 | 29 | 42.6 | 38.6 |
| Pluronic Acid F-68 Conc. (%) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| 0 (Control) | 75 | 75 | 14 | 7 | 5 | 3 | 6.6 | 4 |
| 0.01 | 75 | 75 | 29 | 18 | 17 | 10 | 22.6 | 13.3 |
| 0.05 | 75 | 75 | 35 | 28 | 25 | 19 | 33.3 | 25.3 |
| 0.1 | 75 | 75 | 41 | 33 | 28 | 23 | 37.3 | 30.6 |
| 0.5 | 75 | 75 | 45 | 40 | 33 | 30 | 44 | 40 |
| Incubation Temp. (°C) | Callus Infected | Kan Resistant Calli | Kan Resistant Plants | Transformation Efficiency (%) | ||||
|---|---|---|---|---|---|---|---|---|
| EHA | LBA | EHA | LBA | EHA | LBA | EHA | LBA | |
| 20 | 105 | 105 | 7 | 1 | 2 | 0 | 1.9 | 0 |
| 22 | 105 | 105 | 23 | 16 | 13 | 8 | 12.3 | 7.6 |
| 24 | 105 | 105 | 36 | 27 | 24 | 17 | 22.8 | 16.1 |
| 26 | 105 | 105 | 21 | 10 | 12 | 4 | 11.4 | 3.8 |
| 28 | 105 | 105 | 0 | 0 | 0 | 0 | 0 | 0 |
References
- Kothari, S.; Kumar, S.; Kothari, A.; Watanabe, K. Applications of biotechnology for improvement of millet crops: Review of progress and future prospects. Plant Biotechnol. 2005, 22, 81–88. [Google Scholar] [CrossRef]
- Pokharia, A.; Kharakwal, J.; Srivastava, A. Archaeobotanical evidence of millets in the Indian subcontinent with some observations on their role in the Indus civilization. J. Archaeol. Sci. 2014, 42, 442–455. [Google Scholar] [CrossRef]
- Kothari-Chajer, A.; Sharma, M.; Kachhwaha, S.; Kothari, S. Micronutrient optimization results into highly improved in vitro plant regeneration in kodo (Paspalum scrobiculatum L.) and finger (Eleusine coracana (L.) Gaertn.) millets. Plant Cell Tiss. Org. Cult. 2008, 94, 105–112. [Google Scholar] [CrossRef]
- Shahidi, F.; Chandrasekara, A. Millet grain phenolics and their role in disease risk reduction and health promotion: A review. J. Funct. Foods 2013, 5, 570–581. [Google Scholar] [CrossRef]
- Sood, P.; Singh, R.; Prasad, M. Millets genetic engineering: The progress made and prospects for the future. Plant Cell Tiss. Org. Cult. 2019, 137, 421–439. [Google Scholar] [CrossRef]
- Geervani, P.; Eggum, B. Nutrient composition and protein quality of minor millets. Plant Foods Hum. Nutr. 1989, 39, 201–208. [Google Scholar] [CrossRef]
- Chaudhary, N.; Vyas, S. Effect of germination on proximate composition and anti nutritional factor of millet (ragi) based premixes. Int. J. Food Nutr. Sci. 2014, 3, 72–77. [Google Scholar]
- Taylor, J.; Schober, T.; Bean, S. Novel food and non-food uses for sorghum and millets. J. Cereal Sci. 2006, 44, 252–271. [Google Scholar] [CrossRef]
- Michaelraj, P.; Shanmugam, A. A study on millets based cultivation and consumption in India. Int. J. Mark. Financ. Serv. Manag. Res. 2013, 2, 49–58. [Google Scholar]
- de Wet, J.; Brink, D.; Rao, K.; Mengesha, M. Diversity in kodo millet, Paspalum scrobiculatum. Econ. Bot. 1983, 37, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Hegde, P.; Rajasekaran, N.; Chandra, T. Effects of the antioxidant properties of millet species on oxidative stress and glycemic status in alloxan-induced rats. Nutr. Res. 2005, 25, 1109–1120. [Google Scholar] [CrossRef]
- Devakumar, C.; Chhonkar, P. Integration of Millets in Fortified Foods, 54th ed.; National Academy of Sciences: New Delhi, India, 2012. [Google Scholar]
- Dwivedi, S.; Upadhyaya, H.; Senthilvel, S.; Hash, C.; Fukunaga, K.; Diao, X.; Santra, D.; Baltensperge, D.; Prasad, M. Millets: Genetic and genomic resources. Plant Breed. Rev. 2012, 35, 247–375. [Google Scholar]
- Nadolska-Orczyk, A.; Orczyk, W.; Przetakiewicz, A. Agrobacterium-mediated transformation of cereals—From technique development to its application. Acta Physiol. Plant. 2000, 22, 77–88. [Google Scholar] [CrossRef]
- Tzfira, T.; Citovsky, V. Agrobacterium-mediated genetic transformation of plants: Biology and biotechnology. Curr. Opin. Biotechnol. 2006, 17, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Dosad, S.; Chawla, H. In vitro plant regeneration and transformation studies in millets: Current status and future prospects. Indian J. Plant Physiol. 2016, 21, 239–254. [Google Scholar] [CrossRef]
- Sood, P.; Bhattacharya, A.; Joshi, R.; Gulati, A.; Chanda, S.; Sood, A. A method to overcome the waxy surface, cell wall thickening and polyphenol induced necrosis at wound sites-the major deterrents to Agrobacterium mediated transformation of bamboo, a woody monocot. J. Plant Biochem. Biot. 2014, 23, 69–80. [Google Scholar] [CrossRef]
- Plaza-Wüthrich, S.; Tadele, Z. Millet improvement through regeneration and transformation. Biotechnol. Mol. Biol. Rev. 2012, 7, 48–61. [Google Scholar] [CrossRef]
- Smith, R.; Grando, M.; Li, Y.; Seib, J.; Shatters, R. Transformation of bahiagrass (Paspalum notatum Flugge). Plant Cell Rep. 2002, 20, 1017–1021. [Google Scholar] [CrossRef]
- Gondo, T.; Tsuruta, S.; Akashi, R.; Kawamura, O.; Hoffmann, F. Green, herbicide-resistant plants by particle inflow gun-mediated gene transfer to diploid bahiagrass (Paspalum notatum). J. Plant Physiol. 2005, 162, 1367–1375. [Google Scholar] [CrossRef]
- Wu, X.; Shi, H.; Chen, X.; Liu, Y.; Guo, Z. Establishment of Agrobacterium-mediated transformation of seashore paspalum (Paspalum vaginatum O. Swartz). Vitr. Cell. Dev. Biol. Plant 2018, 54, 545–552. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with Tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Yadav, T.; Kothari, S.; Kachhwaha, S. Optimization of Agrobacterium-Mediated Genetic Transformation and Regeneration of Transgenic Plants in Indian Cultivar of Barley (Hordeum vulgare L. cv. BL 2). Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2013, 83, 255–264. [Google Scholar] [CrossRef]
- Jefferson, R.; Kavanagh, T.; Bevan, M. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Kothari-Chajer, A.; Jagga-Chugh, S.; Kothari, S. Factors influencing Agrobacterium tumefaciens-mediated genetic transformation of Eleusine coracana (L.) Gaertn. Plant Cell Tiss. Org. Cult. 2011, 105, 93–104. [Google Scholar] [CrossRef]
- Botti, C.; Vasil, I. Plant regeneration by somatic embryogenesis from parts of cultured mature embryos of Pennisetum americanum (L.) K. Schum. Z. Pflanzenphysiol. 1983, 111, 319–325. [Google Scholar] [CrossRef]
- Kachhwaha, S.; Varshney, A.; Kothari, S. Somatic embryogenesis and long term high plant regeneration from barley (Hordeum vulgare L.) using picloram. Cereal Res. Commun. 1997, 25, 117–126. [Google Scholar] [CrossRef]
- Chin, D.; Mishiba, K.; Mii, M. Agrobacterium-mediated transformation of protocorm-like bodies in Cymbidium. Plant Cell Rep. 2007, 26, 735–743. [Google Scholar] [CrossRef]
- Wang, M.; Li, Z.; Matthews, P.; Upadhyaya, N.; Waterhouse, P. Improved vectors for Agrobacterium tumefaciens-mediated transformation of monocot plants. Int. Symp. Biotechnol. Trop. Subtrop. Species 1997, 461, 401–408. [Google Scholar] [CrossRef]
- Wang, M.; Li, P.; Li, C.; Pan, Y.; Jiang, X.; Zhu, D.; Zhao, Q.; Yu, J. SiLEA14, a novel atypical LEA protein, confers abiotic stress resistance in foxtail millet. BMC Plant Biol. 2014, 14, 290. [Google Scholar] [CrossRef]
- Sood, P.; Singh, R.K.; Prasad, M. An efficient Agrobacterium-mediated genetic transformation method for foxtail millet (Setaria italica L.). Plant Cell Rep. 2020, 39, 511–525. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enríquez-Obregón, G.; Prieto-Samsónov, D.; Gustavo, A.; Pérez, M.; Selman-Housein, G.; Vázquez-Padrón, R. Agrobacterium-mediated Japonica rice transformation: A procedure assisted by an antinecrotic treatment. Plant Cell Tiss. Org. Cult. 1999, 59, 159–168. [Google Scholar] [CrossRef]
- Hensel, G.; Kastner, C.; Oleszczuk, S.; Riechen, J.; Kumlehn, J. Agrobacterium-mediated gene transfer to cereal crop plants: Current protocols for barley, wheat, triticale, and maize. Int. J. Plant Genom. 2009, 2009, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Reddy, B.; Ramaiah, B.; Sharma, R. Heterosis in white-grained grain mold resistant sorghum hybrids. J. SAT Agric. Res. 2011, 9, 1–6. [Google Scholar]
- Bayer, G.; Yemets, A.; Blume, Y. Obtaining the transgenic lines of finger millet Eleusine coracana (L.). with dinitroaniline resistance. Cytol. Genet. 2014, 48, 139–144. [Google Scholar] [CrossRef]
- Ramadevi, R.; Rao, K.; Reddy, V. Agrobacterium tumefaciens-mediated genetic transformation and production of stable transgenic pearl millet (Pennisetum glaucum [L.] R. Br.). Vitr. Cell. Dev. Biol. Plant 2014, 50, 392–400. [Google Scholar] [CrossRef]
- Ziemienowicz, A. Agrobacterium-mediated plant transformation: Factors, applications and recent advances. Biocatal. Agric. Biotechnol. 2014, 3, 95–102. [Google Scholar] [CrossRef]
- Yadav, T.; Kachhwaha, S.; Kothari, S. Efficient in vitro plant regeneration and generation of transgenic plants in barley (Hordeum vulgare L.) using particle bombardment. J. Plant Biochem. Biot. 2013, 22, 202–213. [Google Scholar] [CrossRef]
- Ceasar, S.; Ignacimuthu, S. Agrobacterium-mediated transformation of finger millet (Eleusine coracana (L.) Gaertn.) using shoot apex explants. Plant Cell Rep. 2011, 30, 1759–1770. [Google Scholar] [CrossRef]
- Tyagi, H.; Rajasubramaniam, S.; Dasgupta, I. Regeneration and Agrobacterium-mediated transformation of a popular indica rice variety, ADT39. Curr. Sci. 2007, 93, 678–683. [Google Scholar]
- Cheng, M.; Lowe, B.; Spencer, T.; Ye, X.; Armstrong, C. Factors influencing Agrobacterium-mediated transformation of monocotyledonous species. Vitr. Cell. Dev. Biol. Plant 2004, 40, 31–45. [Google Scholar] [CrossRef]
- Fullner, K.; Nester, E. Temperature affects the T-DNA transfer machinery of Agrobacterium tumefaciens. J. Bacteriol. 1996, 178, 1498–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillen, W.; De Clereq, J.; Kapila, J.; Zambre, M. The effect of temperature on Agrobacterium tumefaciens-mediated gene transfer to plants. Plant J. 1997, 12, 1459–1463. [Google Scholar] [CrossRef]
- Hiei, Y.; Ishida, Y.; Komari, T. Progress of cereal transformation technology mediated by Agrobacterium tumefaciens. Front. Plant Sci. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, A.; Shilpha, J.; Pandian, S.; Ramesh, M. Agrobacterium-mediated transformation of indica rice cv. ADT 43. Plant Cell Tiss. Org. Cult. 2012, 109, 153–165. [Google Scholar] [CrossRef]
- Mundada, P.; Umdale, S.; Ahire, M.; Kumar, S.; Nikam, T. Transgenic Finger Millet [Eleusine coracana (L.) Gaertn.] for Crop Improvement. In Genetically Modified Crops; Springer: Singapore, 2021; pp. 67–77. [Google Scholar]





| Strain | Culture Medium | Antibiotics |
|---|---|---|
| EHA 105 (pCNL 56) | Luria-Bertini (LB) broth | 50 mg/L Kan |
| LBA 4404 (pCAMBIA 2300) | Yeast extract broth (YEB) | 50 mg/L Kan + 20 mg/L (Rifampicin) Rif + 50 mg/L Streptomycin (Strep) |
| Antinecrotic Mixture (AM) | L-Cysteine (mg/L) | Silver Nitrate (mg/L) | Ascorbic Acid (mg/L) |
|---|---|---|---|
| AM 1 | 10 | 5 | 1 |
| AM 2 | 10 | 5 | 2.5 |
| AM 3 | 10 | 3 | 2.5 |
| Variety | 2,4-D Conc. (mg/L) | Callus Response (%) | Avg. No. of Regenerated Shoots/Explant (Mean ± SE) |
|---|---|---|---|
| TNAU 86 | 1 | 28 | 1.2 ± 0.2 |
| 2 | 45 | 3.0 ± 0 | |
| 3 | 62 | 4.4 ± 1.1 | |
| 4 | 20 | 2.3 ± 0.8 | |
| RK-390-25 | 1 | 13 | 0 |
| 2 | 17 | 0 | |
| 3 | 22 | 1.0 ± 0.2 | |
| 4 | 20 | 0 | |
| RK 15 | 1 | 20 | 0 |
| 2 | 34 | 1.1 ± 0.8 | |
| 3 | 35 | 2.3 ± 0.5 | |
| 4 | 30 | 0 | |
| GPUK-3 | 1 | 34 | 2.1 ± 0.3 |
| 2 | 57 | 3.2 ± 1.1 | |
| 3 | 78 | 6.8 ± 1.5 | |
| 4 | 32 | 2.5 ± 0 | |
| RBK-155 | 1 | 18 | 0 |
| 2 | 33 | 1.5 ± 0.7 | |
| 3 | 28 | 1.3 ± 0.5 | |
| 4 | 25 | 1.2 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatt, R.; Asopa, P.P.; Jain, R.; Kothari-Chajer, A.; Kothari, S.L.; Kachhwaha, S. Optimization of Agrobacterium Mediated Genetic Transformation in Paspalum scrobiculatum L. (Kodo Millet). Agronomy 2021, 11, 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061104
Bhatt R, Asopa PP, Jain R, Kothari-Chajer A, Kothari SL, Kachhwaha S. Optimization of Agrobacterium Mediated Genetic Transformation in Paspalum scrobiculatum L. (Kodo Millet). Agronomy. 2021; 11(6):1104. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061104
Chicago/Turabian StyleBhatt, Ritika, Prem Prakash Asopa, Rohit Jain, Aditi Kothari-Chajer, Shanker Lal Kothari, and Sumita Kachhwaha. 2021. "Optimization of Agrobacterium Mediated Genetic Transformation in Paspalum scrobiculatum L. (Kodo Millet)" Agronomy 11, no. 6: 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061104