
Quantitative Trait Locus Analysis of Microscopic Phenotypic Characteristic Data Obtained Using Optical Coherence Tomography Imaging of Rice Bacterial Leaf Blight Infection in the Field


 , , ,
, , , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Rice Materials
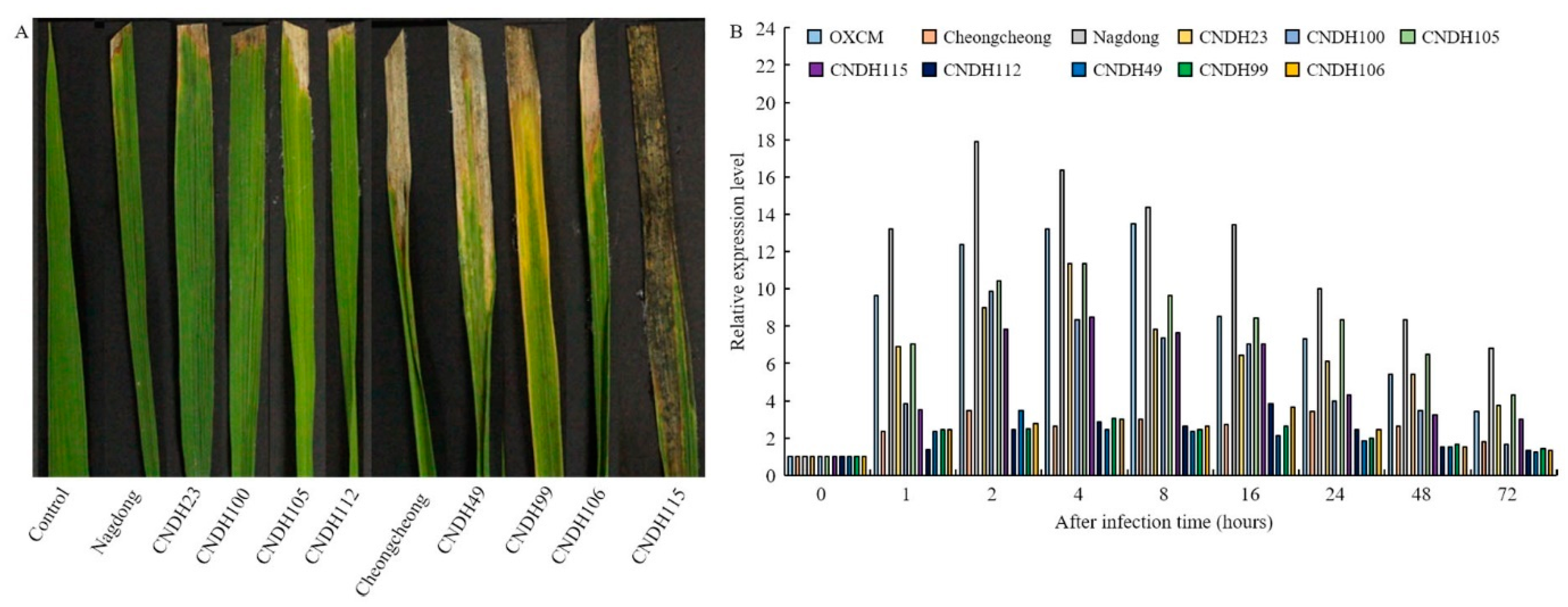
2.2. K3 Strain Culture Media, Inoculation, and Infection Assessment
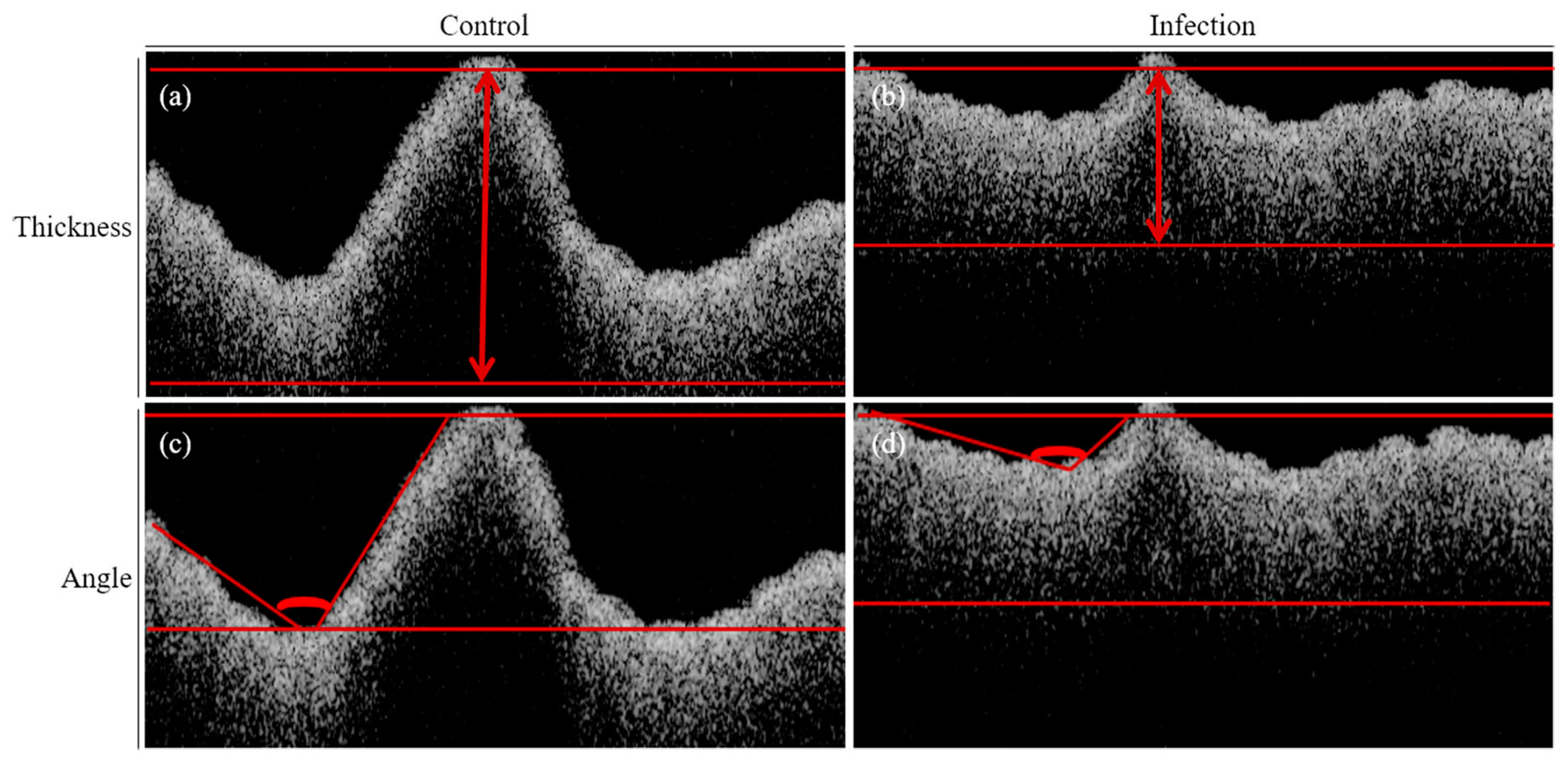
2.3. Optical Coherence Tomography (OCT) Analysis of Infected Rice Leaves
2.4. Quantitative Trait Loci (QTL) Analysis
2.5. Target Gene Selection and Physical Mapping
2.6. RNA Extraction
2.7. Quantitative RT-PCR Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Rice Research Institute World Rice Statistics 2013; IRRI: Los Banos, Philippines, 2013.
- Shankar, K.R.; Guddanti, N. Handbook of Climate Change Adaptation; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Dixit, S.; Singh, U.M.; Singh, A.K.; Alam, S.; Venkateshwarlu, C.; Nachimuthu, V.V.; Yadav, S.; Abbai, R.; Selvaraj, R.; Devi, M.N.; et al. Marker Assisted Forward Breeding to Combine Multiple Biotic-Abiotic Stress Resistance/Tolerance in Rice. Rice 2020. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Yan, J.; Liang, Y.; Shi, Y.; He, Z.; Wu, Y.; Zeng, Q.; Liu, X.; Peng, J. Resistance Genes and their Interactions with Bacterial Blight/Leaf Streak Pathogens (Xanthomonas oryzae) in Rice (Oryza sativa L.)—An Updated Review. Rice 2020. [Google Scholar] [CrossRef] [Green Version]
- Zou, G.; Zhai, G.; Feng, Q.; Yan, S.; Wang, A.; Zhao, Q.; Shao, J.; Zhang, Z.; Zou, J.; Han, B.; et al. Identification of QTLs for eight agronomically important traits using an ultra-high-density map based on SNP. J. Exp. Bot. 2012, 63, 5451–5462. [Google Scholar] [CrossRef] [Green Version]
- Bustos-Korts, D.; Malosetti, M.; Chenu, K.; Chapman, S.; Boer, M.P.; Zheng, B.; Van Eeuwijk, F.A. From QTLs to Adaptation Landscapes: Using Genotype-To-Phenotype Models to Characterize G×E Over Time. Front. Plant Sci. 2019, 10, 1540. [Google Scholar] [CrossRef] [PubMed]
- Khahani, B.; Tavakol, E.; Shariati, V.; Fornara, F. Genome wide screening and comparative genome analysis for Meta-QTLs, ortho-MQTLs and candidate genes controlling yield and yield-related traits in rice. BMC Genom. 2020, 21, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.R.; Macdonald, S.J. Dissecting the genetic basis of variation in Drosophila sleep using a multiparental QTL mapping resource. Genes 2020, 11, 294. [Google Scholar] [CrossRef] [Green Version]
- Ravichandran, N.K.; Wijesinghe, R.E.; Shirazi, M.F.; Park, K.; Lee, S.-Y.; Jung, H.-Y.; Jeon, M.; Kim, J. In vivo monitoring on growth and spread of gray leaf spot disease in capsicum annuum leaf using spectral domain optical coherence tomography. J. Spectrosc. 2016. [Google Scholar] [CrossRef] [Green Version]
- Trojanowski, M.; Kraszewski, M.; Strąkowski, M.R.; Pluciński, J. Optical Coherence Tomography for nanoparticles quantitative characterization. Nanoimaging Nanospectrosc. III 2015. [Google Scholar] [CrossRef]
- Barreiro, P.; Zheng, C.; Sun, D.-W.; Sanchez, N.H.; Perez-Sanchez, J.; Ruiz-Cabello, J. Non-destructive seed detection in mandarins: Comparison of automatic threshold methods in FLASH and COMSPIRA MRIs. Postharvest. Biol. Technol. 2008, 47, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Van As, H. Intact plant MRI for the study of cell water relations, membrane permeability, cell-to-cell and long distance water transport. J. Exp. Bot. 2007, 58, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Meglinski, I.V.; Buranachai, C.; Terry, L.A. Plant photonics: Application of optical coherence tomography to monitor defects and rots in onion. Laser Phys. Lett. 2010, 7, 307–310. [Google Scholar] [CrossRef]
- Lee, C.; Lee, S.Y.; Jung, H.Y.; Kim, J. The application of optical coherence tomography in the diagnosis of marssonina blotch in apple leaves. J. Opt. Soc. Korea 2012, 16, 133–140. [Google Scholar] [CrossRef]
- Kutis, I.S.; Sapozhnikova, V.V.; Kuranov, R.V.; Kamenskii, V.A. Study of the morphological and functional state of higher plant tissues by optical coherence microscopy and optical coherence tomography. Russ. J. Plant Physiol. 2005, 52, 559–564. [Google Scholar] [CrossRef]
- Wijesinghe, R.E.; Lee, S.-Y.; Ravichandran, N.K.; Shirazi, M.F.; Kim, P.; Jung, H.-Y.; Jeon, M.; Kim, J. Biophotonic approach for the characterization of initial bitter-rot progression on apple specimens using optical coherence tomography assessments. Sci. Rep. 2018, 8, 15816. [Google Scholar] [CrossRef] [PubMed]
- Manattayil, J.K.; Ravichandran, N.K.; Wijesinghe, R.E.; Shirazi, M.F.; Lee, S.-Y.; Kim, P.; Jung, H.-Y.; Jeon, M.; Kim, J. Non-destructive classification of diversely stained capsicum annuum seed specimens of different cultivars using near-infrared imaging based optical intensity detection. Sensors 2018, 18, 2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Jeesook, E.; Kim, S.H. Crops Disease Diagnosing Using Image-Based Deep Learning Mechanism. In Proceedings of the 2018 International Conference on Computing and Network Communications (CoCoNet), Astana, Kazakhstan, 15–17 August 2018; pp. 23–26. [Google Scholar] [CrossRef]
- Shimada, Y.; Nakagawa, H.; Sadr, A.; Wada, I.; Nakajima, M.; Nikaido, T.; Otsuki, M.; Tagami, J.; Sumi, Y. Noninvasive cross-sectional imaging of proximal caries using swept-source optical coherence tomography (SS-OCT) in vivo. J. Biophotonics 2014, 7, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Yun, B.W.; Kim, K.M. Analysis of QTLs associated with the rice quality related gene by double haploid populations. Int. J. Genom. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffman, H.E. An improved technique for evaluat-ing resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]
- Kim, H.; Du, X.X.; Kim, S.; Kim, P.; Wijesinghe, R.E.; Yun, B.J.; Kim, K.M.; Jeon, M.; Kim, J. Non-invasive morphological characterization of rice leaf bulliform and aerenchyma cellular regions using low coherence interferometry. Appl. Sci. 2019, 9, 2104. [Google Scholar] [CrossRef] [Green Version]
- McCough, S.R.; Doerge, R.W. QTL mapping in rice. Trends Genet. 1995, 11, 482–487. [Google Scholar] [CrossRef]
- Yasmin, S.; Zaka, A.; Imran, A.; Zahid, M.A.; Yousaf, S.; Rasul, G.; Arif, M.; Mirza, M.S. Plant growth promotion and suppression of bacterial leaf blight in rice by inoculated bacteria. PLoS ONE 2016, 11, e0160688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Naeem, M.; Iqbal, M. Breeding approaches for bacterial leaf blight resistance in rice (Oryza sativa L.), current status and future directions. Eur. J. Plant Pathol. 2014, 139, 27–37. [Google Scholar] [CrossRef]
- Muhammad, S.; Tahira, B.; Hafiz, U.F.; Zulqarnain, H.; Imad, N.; Abid, M.; Muhammad, A. Molecular screening of rice (Oryza sativa L.) germplasm for Xa4, xa5 and Xa21 bacterial leaf blight (BLB) resistant genes using linked marker approach. Afr. J. Biotechnol. 2016, 15, 2317–2324. [Google Scholar] [CrossRef] [Green Version]
- Borrill, P.; Harrington, S.A.; Uauy, C. Applying the latest advances in genomics and phenomics for trait discovery in polyploid wheat. Plant J. 2019, 97, 56–72. [Google Scholar] [CrossRef]
- Furbank, R.T.; Jimenez-Berni, J.A.; George-Jaeggli, B.; Potgieter, A.B.; Deery, D.M. Field crop phenomics: Enabling breeding for radiation use efficiency and biomass in cereal crops. New Phytol. 2019, 223, 1714–1727. [Google Scholar] [CrossRef] [Green Version]
- Bock, C.H.; Poole, G.H.; Parker, P.E.; Gottwald, T.R. Plant disease severity estimated visually, by digital photography and image analysis, and by hyperspectral imaging. CRC Crit. Rev. Plant Sci. 2010, 29, 59–107. [Google Scholar] [CrossRef]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated Digital Image Analysis for Rapid and Accurate Measurement of Leaf Area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef]
- Kumar, A.; Guha, A.; Bimolata, W.; Reddy, A.R.; Laha, G.S.; Sundaram, R.M.; Pandey, M.K.; Ghazi, I.A. Leaf gas exchange physiology in rice genotypes infected with bacterial blight: An attempt to link photosynthesis with disease severity and rice yield. Aust. J. Crop Sci. 2013, 7, 32–39. [Google Scholar]
- Hettinger, J.W.; De La Peña Mattozzi, M.; Myers, W.R.; Williams, M.E.; Reeves, A.; Parsons, R.L.; Haskell, R.C.; Petersen, D.C.; Wang, R.; Medford, J.I. Optical coherence microscopy. A technology for rapid, in vivo, non-destructive visualization of plants and plant cells. Plant Physiol. 2000, 123, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-R.; Yun, S.; Jan, R.; Kim, K.-M. Screening and Identification of Brown Planthopper Resistance Genes OsCM9 in Rice. Agronomy 2020, 10, 1865. [Google Scholar] [CrossRef]
- Kim, E.; Yun, S.; Park, J.; Kim, K.-M. Identification of F3H, Major Secondary Metabolite-Related Gene That Confers Resistance against Whitebacked Planthopper through QTL Mapping in Rice. Plants 2021, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Sulewski, P. Normal distribution with plasticizing component. Commun. Stat.-Theory Methods 2020, 1–30. [Google Scholar] [CrossRef]
- Javed, T.; Shabbir, R.; Ali, A.; Afzal, I.; Zaheer, U.; Gao, S.J. Transcription factors in plant stress responses: Challenges and potential for sugarcane improvement. Plants 2020, 9, 491. [Google Scholar] [CrossRef]
- Liu, J.; Ding, G.; Gai, Z.; Zhang, W.; Han, Y.; Li, W. Changes in the gene expression profile of Arabidopsis thaliana under chromium stress. Ecotoxicol. Environ. Saf. 2020, 193, 110302. [Google Scholar] [CrossRef] [PubMed]
- Bhadouriya, S.L.; Mehrotra, S.; Basantani, M.K.; Loake, G.J.; Mehrotra, R. Role of Chromatin Architecture in Plant Stress Responses: An Update. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Meraj, T.A.; Fu, J.; Raza, M.A.; Zhu, C.; Shen, Q.; Xu, D.; Wang, Q. Transcriptional Factors Regulate Plant Stress Responses through Mediating Secondary Metabolism. Genes 2020, 11, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kliebenstein, D.J. Making new molecules-evolution of structures for novel metabolites in plants. Curr. Opin. Plant Biol. 2013, 16, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.D.; Chen, Z.W.; Deng, Y.; Lan, T.; Guan, H.Z.; Duan, Y.L.; Zhou, Y.C.; Lin, M.C.; Wu, W.R. Fine Mapping of qBlsr5a, a QTL Controlling Resistance to Bacterial Leaf Streak in Rice. Acta Agron. Sin. 2008, 34, 587–590. [Google Scholar]
- Tang, D.; Wu, W.; Li, W.; Lu, H.; Worland, A.J. Mapping of QTLs conferring resistrance to bacterial leaf streak in rice. Theor. Appl. Genet. 2000, 101, 286–291. [Google Scholar] [CrossRef]
- Xiao, J.; Li, J.; Yuan, L.; Tanksley, S.D. Identification of QTLs affecting traits of agronomic importance in a recombinant inbred population derived from a subspecific rice cross. Theor. Appl. Genet. 1996, 92, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: Latest discoveries and future prospects. Plant Cell Rep. 2021. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Parents | DH z Population | ||||||
|---|---|---|---|---|---|---|---|---|
| Cheongcheong | Nagdong | |||||||
| Control | Infection | Control | Infection | Control | Infection | |||
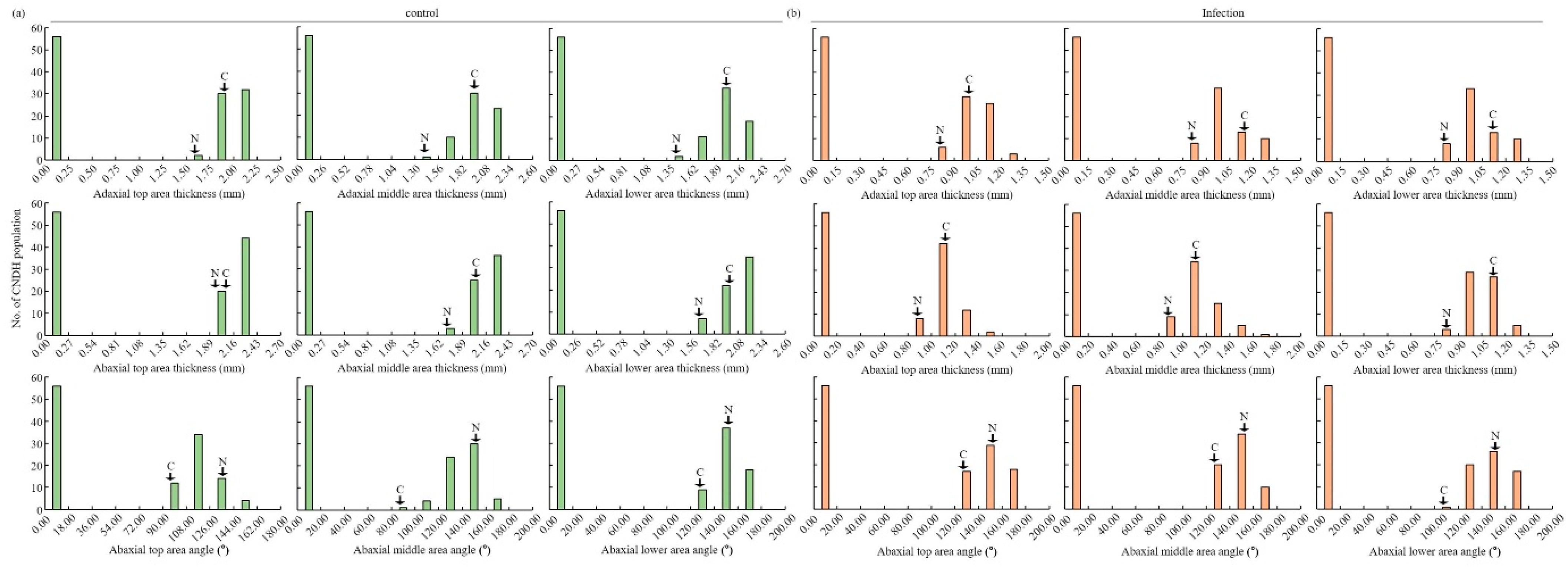
| Adaxial | Thickness (mm) | Top | 1.97 ± 0.83 | 1.10 ± 0.81 | 1.40 ± 0.68 | 0.68 ± 0.74 | 0.93 ± 0.88 | 0.48 ± 0.45 |
| Middle | 2.02 ± 0.81 | 1.14 ± 0.90 | 1.20 ± 0.69 | 0.69 ± 0.92 | 0.93 ± 0.88 | 0.47 ± 0.45 | ||
| Lower | 2.10 ± 0.93 | 1.13 ± 0.86 | 1.17 ± 0.92 | 0.64 ± 0.67 | 0.95 ± 0.90 | 0.49 ± 0.46 | ||
| Abaxial | Thickness (mm) | Top | 2.08 ± 0.79 | 1.31 ± 0.81 | 1.76 ± 0.79 | 0.60 ± 0.91 | 1.04 ± 0.98 | 0.49 ± 0.47 |
| Middle | 2.08 ± 0.81 | 1.50 ± 0.91 | 1.38 ± 0.62 | 0.63 ± 0.84 | 1.01 ± 0.95 | 0.51 ± 0.50 | ||
| Lower | 2.06 ± 0.89 | 1.16 ± 0.73 | 1.44 ± 0.91 | 0.71 ± 0.69 | 0.97 ± 0.91 | 0.49 ± 0.46 | ||
| Angle (°) | Top | 72.83 ± 0.92 | 108.06 ± 0.79 | 139.0 ± 0.80 | 151.74 ± 0.65 | 54.21 ± 51.97 | 69.64 ± 66.08 | |
| Middle | 71.90 ± 0.82 | 105.64 ± 0.81 | 158.13 ± 0.93 | 156.21 ± 0.93 | 64.93 ± 61.88 | 67.95 ± 64.34 | ||
| Lower | 106.98 ± 0.74 | 78.13 ± 0.85 | 158.55 ± 0.91 | 145.58 ± 0.91 | 71.39 ± 67.51 | 64.60 ± 61.61 | ||
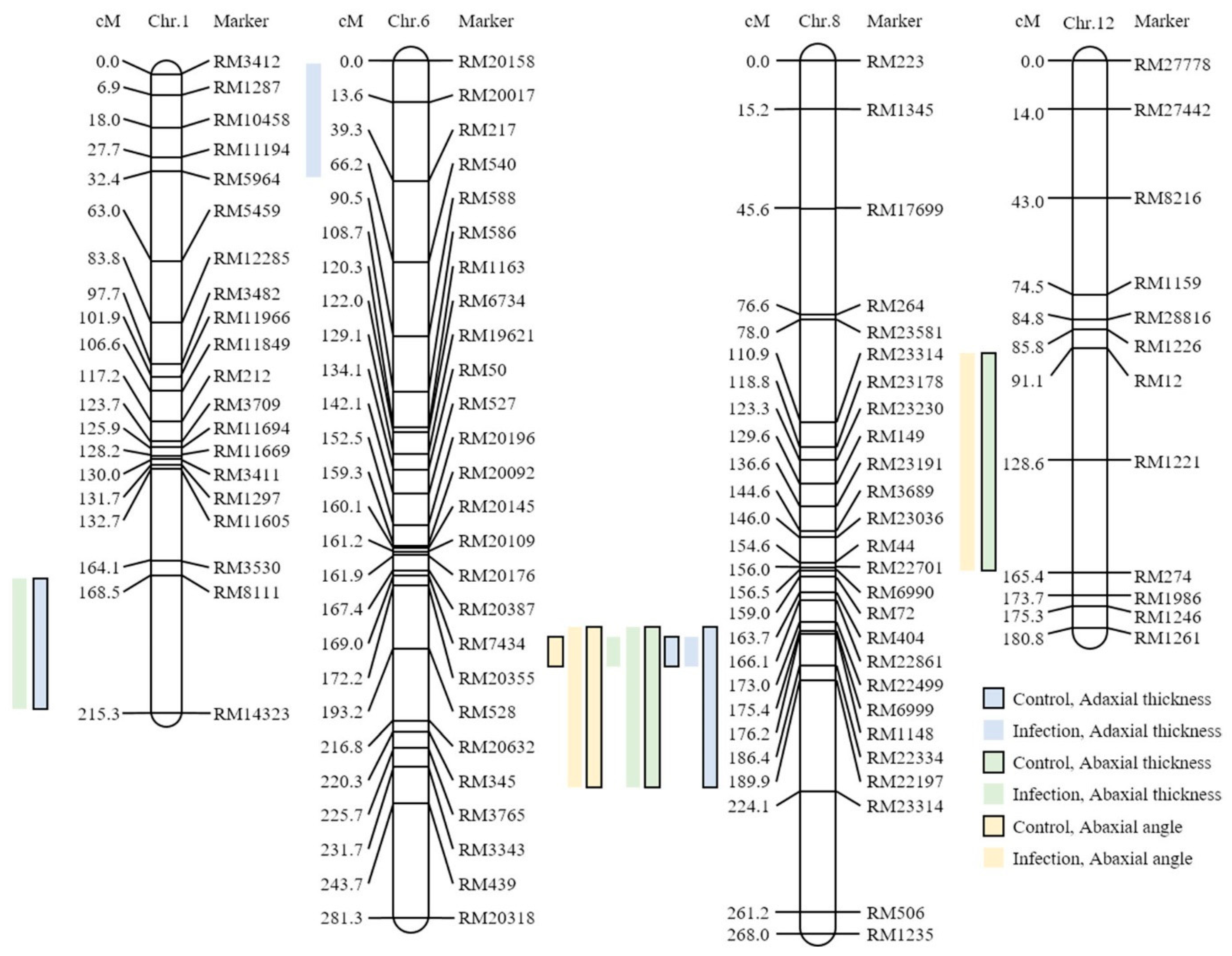
| Characteristics | QTL | Chromosome | Interval Markers z | LOD | Additive Effect y | R2 x | Increasing Effects w | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Adaxial | Thickness | Top | Control | qco-1t-8 | 8 | RM22499-RM23314 | 4.85 | −0.38 | 0.33 | Nagdong |
| Infection | qio-1t-8 | 8 | RM6999-RM22334 | 3.52 | −0.18 | 0.36 | Nagdong | |||
| Middle | Control | qco-2t-1 | 1 | RM8111-RM14323 | 52.23 | 0.85 | 0.97 | Cheongcheong | ||
| qco-2t-8 | 8 | RM22499-RM22334 | 4.58 | −0.36 | 0.38 | Nagdong | ||||
| Infection | qio-2t-8 | 8 | RM6999-RM22334 | 4.81 | −0.20 | 0.39 | Nagdong | |||
| Lower | Control | qco-3t-1 | 1 | RM8111-RM14323 | 48.49 | 0.86 | 0.95 | Cheongcheong | ||
| qco-3t-8 | 8 | RM6999-RM23314 | 4.36 | −0.37 | 0.34 | Nagdong | ||||
| Infection | qio-3t-6 | 6 | RM20158-RM217 | 2.96 | 0.14 | 0.31 | Cheongcheong | |||
| Abaxial | Thickness | Top | Control | qco-1bt-8 | 8 | RM22499-RM22334 | 4.68 | −0.43 | 0.36 | Nagdong |
| Infection | qio-1bt-1 | 1 | RM8111-RM14323 | 34.85 | 0.45 | 0.93 | Cheongcheong | |||
| qio-1bt-8 | 8 | RM22499-RM22334 | 3.86 | −0.19 | 0.36 | Nagdong | ||||
| Middle | Control | qco-2bt-8 | 8 | RM22499-RM22334 | 5.22 | −0.45 | 0.38 | Nagdong | ||
| Infection | qio-2bt-8 | 8 | RM6999-RM23314 | 3.48 | −0.19 | 0.31 | Nagdong | |||
| Lower | Control | qco-3bt-8 | 8 | RM6999-RM22334 | 3.18 | −0.34 | 0.35 | Nagdong | ||
| qco-3bt-12 | 12 | RM12-RM247 | 57.78 | 0.88 | 0.96 | Cheongcheong | ||||
| Infection | qio-3bt-8 | 8 | RM6999-RM23314 | 5.09 | −0.22 | 0.37 | Nagdong | |||
| Angle | Top | Control | qc0-1ba-8 | 8 | RM22499-RM22334 | 3.67 | −20.35 | 0.34 | Nagdong | |
| Infection | qio-1ba-8 | 8 | RM22499-RM22334 | 4.58 | −29.36 | 0.36 | Nagdong | |||
| Middle | Control | qco-2ba-8 | 8 | RM6999-RM22334 | 3.71 | −24.79 | 0.35 | Nagdong | ||
| Infection | qio-2ba-8 | 8 | RM22499-RM22334 | 3.39 | −24.64 | 0.35 | Nagdong | |||
| qio-2ba-12 | 12 | RM12-RM247 | 59.32 | 62.10 | 0.97 | Cheongcheong | ||||
| Lower | Control | qco-3ba-8 | 8 | RM6999-RM22334 | 3.66 | −26.07 | 0.34 | Nagdong | ||
| Infection | qio-3ba-8 | 8 | RM6999-RM22334 | 3.66 | −24.96 | 0.35 | Nagdong | |||
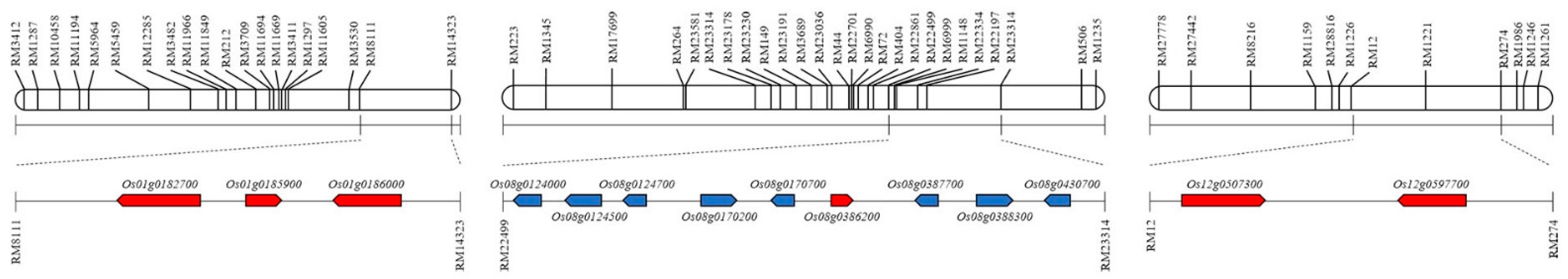
| Chromosome | Marker Interval | Locus | Description |
|---|---|---|---|
| 3 | RM15749-RM15689 | Os03g0686900 | Glycoside hydrolase, family 13, N-terminal domain containing protein |
| Os03g0692000 | Glycosyl transferase, family 14 protein | ||
| Os03g0692500 | Galectin, galactose-binding lectin family protein | ||
| Os03g0693600 | Similar to Indole-3-acetate beta-glucosyltransferase (EC 2.4.1.121) | ||
| Os03g0693700 | Similar to Oxalate oxidase 1 (EC 1.2.3.4) (Germin) | ||
| Os03g0693800 | Similar to Oxalate oxidase 1 (EC 1.2.3.4) (Germin) | ||
| Os03g0693900 | Similar to Oxalate oxidase 1 (EC 1.2.3.4) (Germin) | ||
| Os03g0694000 | Similar to Oxalate oxidase 1 (EC 1.2.3.4) (Germin) | ||
| Os03g0695600 | Proteasome subunit beta type 2 (EC 3.4.25.1) (20S proteasome alpha subunit D) | ||
| Os03g0695700 | Similar to Chloride channel protein CLC-d (AtCLC-d) | ||
| Os03g0696300 | CCAAT-binding transcription factor, subunit B family protein | ||
| Os03g0698800 | Zinc finger, CCCH-type domain containing protein | ||
| Os03g0698900 | Alkaline phytoceramidase family protein | ||
| Os03g0701200 | Similar to Sugar-starvation induced protein (Fragment) | ||
| Os03g0702000 | UDP-glucuronosyl/UDP-glucosyltransferase family protein | ||
| Os03g0702500 | UDP-glucuronosyl/UDP-glucosyltransferase family protein | ||
| Os03g0703000 | Similar to Beta-glucosidase | ||
| Os03g0703100 | Similar to Beta-glucosidase | ||
| Os03g0703200 | Protein kinase-like domain containing protein | ||
| Os03g0704700 | Oxysterol-binding protein family protein | ||
| Os03g0706900 | Zinc finger, RING-type domain containing protein | ||
| Os03g0707600 | OsGAI | ||
| Os03g0708100 | Phytanoyl-CoA dioxygenase family protein | ||
| Os03g0708900 | Zinc finger, RanBP2-type domain containing protein | ||
| Os03g0710100 | Protein kinase-like domain containing protein | ||
| Os03g0710500 | Similar to Luminal binding protein 2 precursor (BiP2) | ||
| 8 | RM149-RM23191 | Os08g0439000 | Phosphofructokinase family protein |
| Os08g0439900 | Mitochondrial glycoprotein family protein | ||
| Os08g0440100 | Similar to Temperature stress-induced lipocalin | ||
| Os08g0442300 | Similar to Calcineurin-like protein | ||
| Os08g0452500 | Auxin responsive SAUR protein family protein | ||
| Os08g0452900 | Non-protein coding transcript, unclassifiable transcript | ||
| Os08g0453200 | Dormancyauxin associated family protein | ||
| Os08g0459700 | Similar to Adenosine diphosphate glucose pyrophosphatase precursor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.-X.; Park, J.-R.; Kim, H.; Saleah, S.A.; Yun, B.-J.; Jeon, M.; Kim, K.-M. Quantitative Trait Locus Analysis of Microscopic Phenotypic Characteristic Data Obtained Using Optical Coherence Tomography Imaging of Rice Bacterial Leaf Blight Infection in the Field. Agronomy 2021, 11, 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081630
Du X-X, Park J-R, Kim H, Saleah SA, Yun B-J, Jeon M, Kim K-M. Quantitative Trait Locus Analysis of Microscopic Phenotypic Characteristic Data Obtained Using Optical Coherence Tomography Imaging of Rice Bacterial Leaf Blight Infection in the Field. Agronomy. 2021; 11(8):1630. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081630
Chicago/Turabian StyleDu, Xiao-Xuan, Jae-Ryoung Park, Hyeree Kim, Sm Abu Saleah, Byoung-Ju Yun, Mansik Jeon, and Kyung-Min Kim. 2021. "Quantitative Trait Locus Analysis of Microscopic Phenotypic Characteristic Data Obtained Using Optical Coherence Tomography Imaging of Rice Bacterial Leaf Blight Infection in the Field" Agronomy 11, no. 8: 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081630