
Identification of Drought Tolerance on the Main Agronomic Traits for Rice (Oryza sativa L. ssp. japonica) Germplasm in China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Material
2.2. Field Management
2.3. Drought Trials and Investigations in the Field
2.4. Large-Scale Germplasm Adjustments for Screening
2.5. Statistical Analysis
2.5.1. Phenotypic Data Analysis
2.5.2. Graphical Representation
3. Results
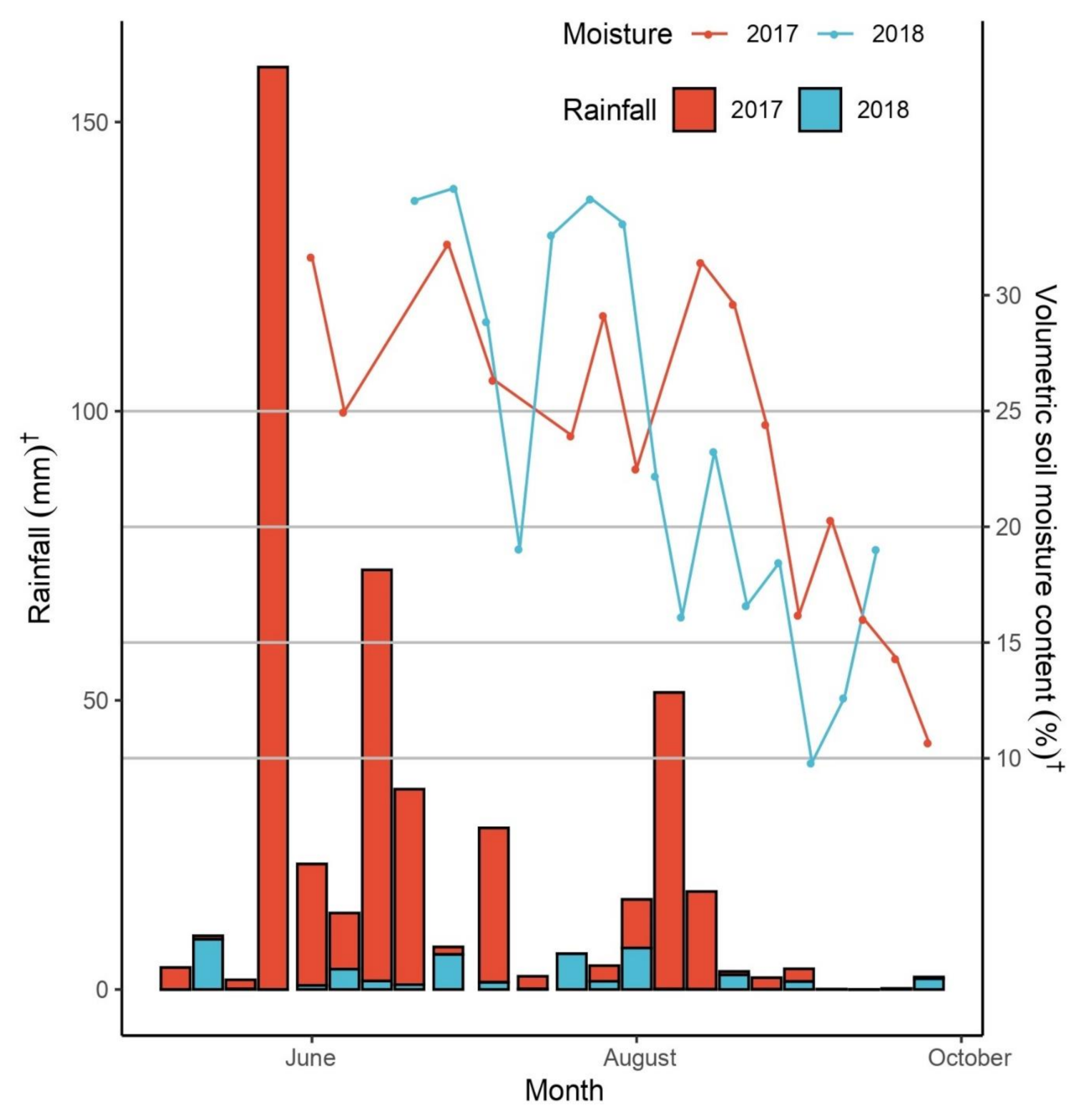
3.1. Meteorological and Soil Observations
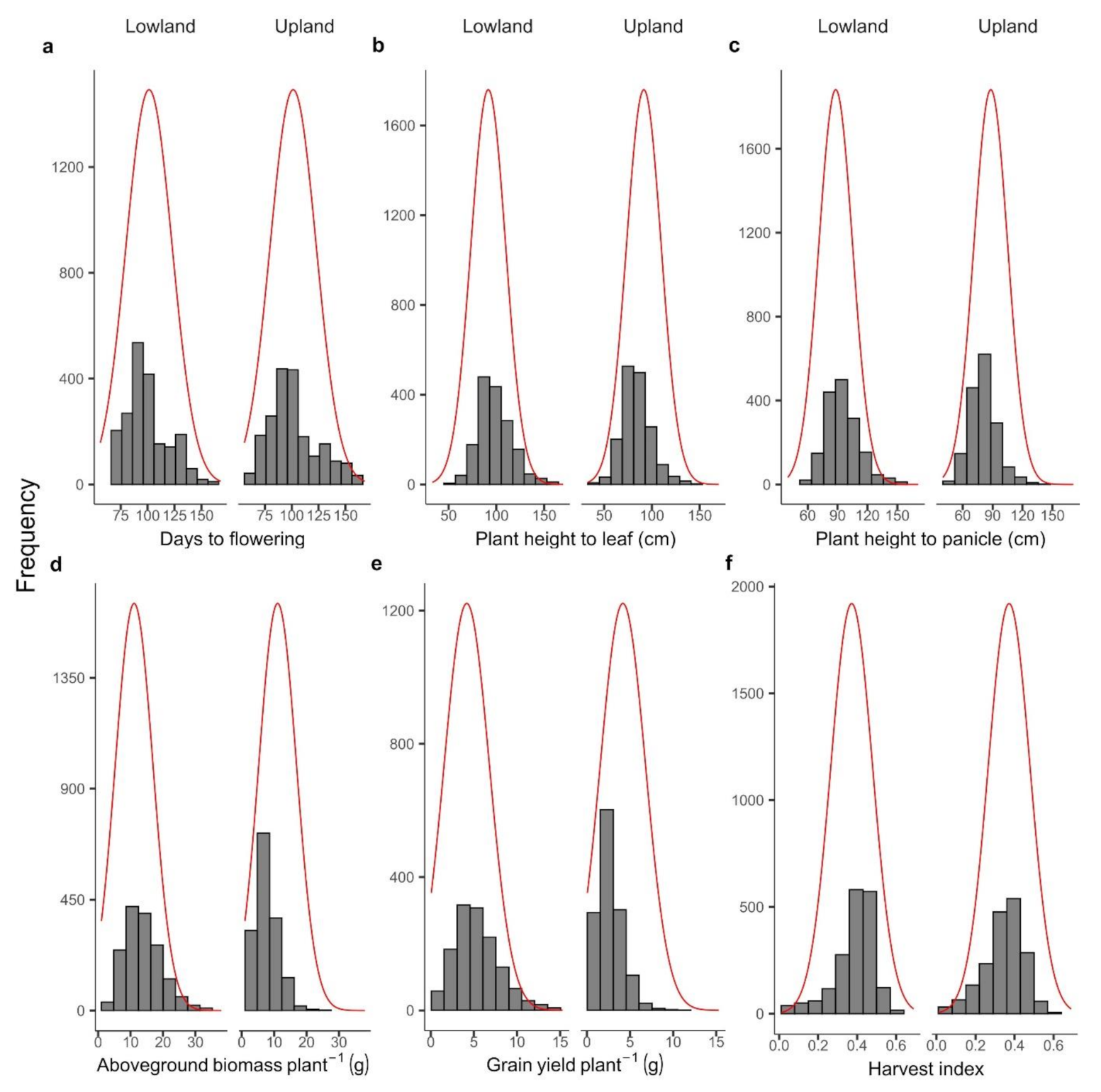
3.2. Variation of Main Agronomic Traits among Controls and Populations
3.3. Determination of Genetic Variation
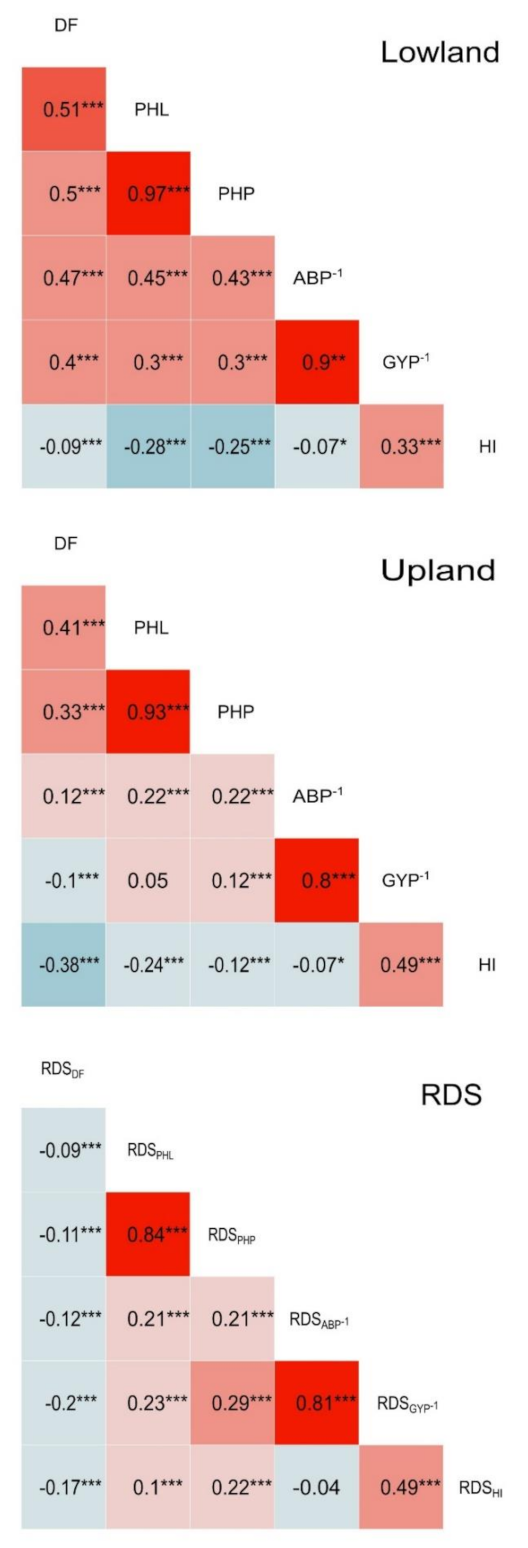
3.4. Correlational Studies of Traits in Lowland and Upland Conditions along with Their RDS Index
3.5. Relative Drought Stress Susceptibility (RDS) and Drought Resistance Grade (DRG) of Genotypes
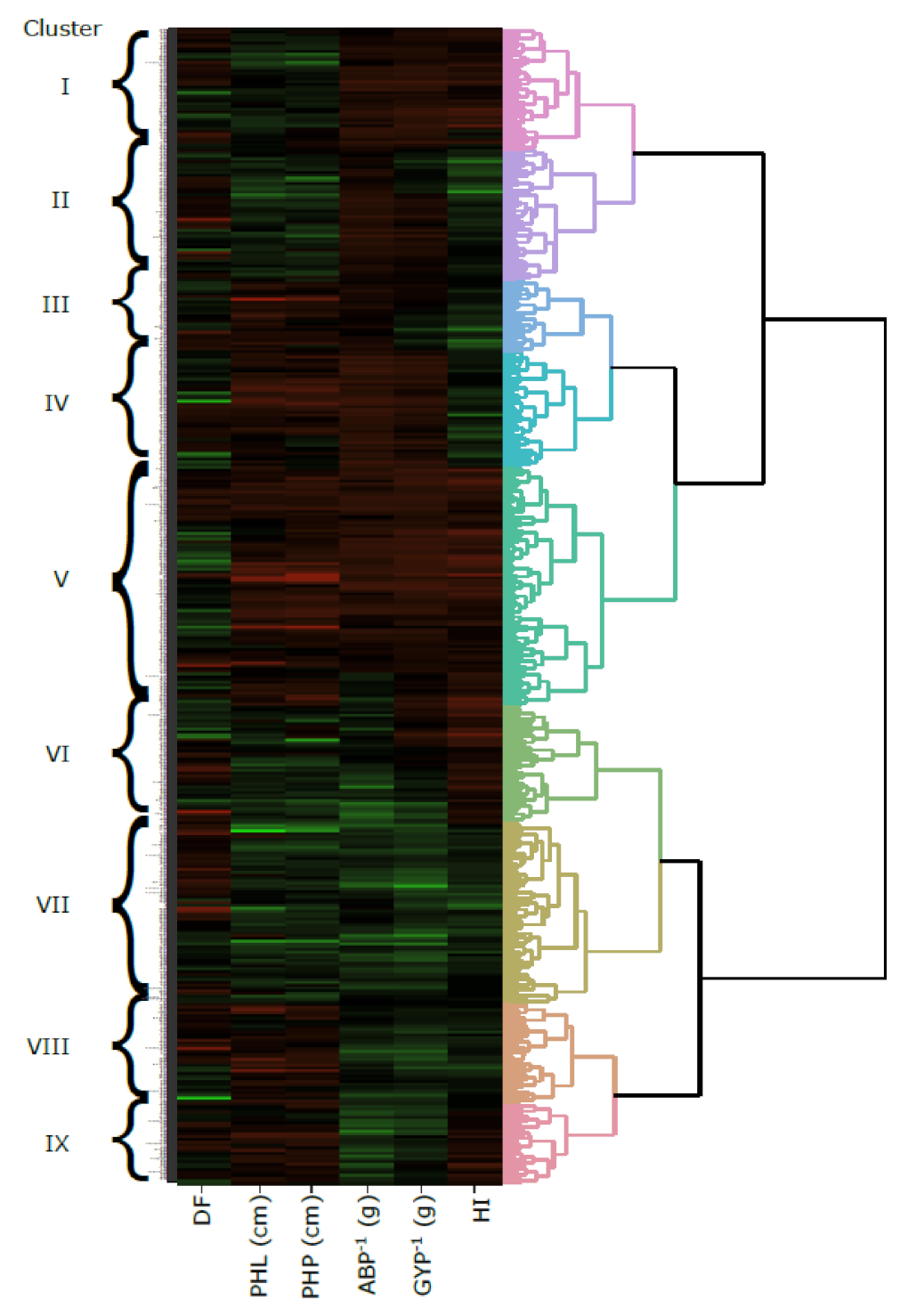
3.6. Agglomerative Hierarchical Clustering (AHC)
3.7. Integrated Elite Genotypes Selection
4. Discussion
4.1. Why Field Identification?
4.2. Augmented Randomized Complete Block Design (ARCBD) Application in Rice Phenotyping
4.3. Traits Investigated under Drought Stress
4.4. Integrated Relative Drought Stress Susceptibility (RDS-I) Derivation and Estimation
4.5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Estrela, T.; Vargas, E. Drought management plans in the European Union. The case of Spain. Water Resour. Manag. 2012, 26, 1537–1553. [Google Scholar] [CrossRef]
- Liu, J.; Diamond, J. China’s environment in a globalizing world. Nature 2006, 435, 1179–1186. [Google Scholar] [CrossRef]
- Rice around the world. In Rice Almanac; Global Rice Science Partnership, International Rice Research Institute: Los Banos, Philippines, 2013; pp. 79–260. ISBN 978-9712203008. Available online: http://books.irri.org/9789712203008_content.pdf (accessed on 16 August 2021).
- Belder, P.; Spiertz, J.H.J.; Bouman, B.A.M.; Lu, G.; Tuong, T.P. Nitrogen economy and water productivity of lowland rice under water-saving irrigation. Field Crop. Res. 2005, 93, 169–185. [Google Scholar] [CrossRef]
- Jockson, M.T. International Rice Genebank Operations Manual; International Rice Research Institute: Los Banos, Philippines, 2000. [Google Scholar]
- Kumar, A.; Bernier, J.; Verulkar, S.; Lafitte, H.R.; Atlin, G.N. Breeding for drought tolerance: Direct selection for yield, response to selection and use of drought-tolerant donors in upland and lowland-adapted populations. Field Crop. Res. 2008, 107, 221–231. [Google Scholar] [CrossRef]
- Jongdee, B.; Pantuwan, G.; Fukai, S.; Fischer, K. Improving drought tolerance in rainfed lowland rice: An example from Thailand. Agric. Water Manag. 2006, 80, 225–240. [Google Scholar] [CrossRef]
- Lafitte, H.R.; Li, Z.K.; Vijayakumar, C.H.M.; Gao, Y.M.; Shi, Y.; Xu, J.L.; Fu, B.Y.; Yu, S.B.; Ali, A.J.; Domingo, J.; et al. Improvement of rice drought tolerance through backcross breeding: Evaluation of donors and selection in drought nurseries. Field Crop. Res. 2006, 97, 77–86. [Google Scholar] [CrossRef]
- Bernier, J.; Atlin, G.N.; Serraj, R.; Kumar, A.; Spaner, D. Breeding upland rice for drought resistance. J. Sci. Food Agric. 2008, 88, 927–939. [Google Scholar] [CrossRef]
- Luo, L.J. Breeding for water-saving and drought-resistance rice (WDR) in China. J. Exp. Bot. 2010, 61, 3509–3517. [Google Scholar] [CrossRef] [Green Version]
- Vikram, P.; Swamy, B.P.M.; Dixit, S.; Ahmed, H.U.; Teresa Sta Cruz, M.; Singh, A.K.; Kumar, A. qDTYI1.1, a major QTL for rice grain yield under reproductive-stage drought stress with a consistent effect in multiple elite genetic backgrounds. BMC Genet. 2011, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Wang, X.; Wang, H.; Xin, H.; Yang, X.; Yan, J.; Li, J.; Tran, L.-S.P.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; et al. Genome-wide analysis of ZmDREB genes and theirassociation with natural variation in drought tolerance at seedling stage of Zea mays L. PLoS Genet. 2013, 9, e1003790. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Wang, Y. Overexpression of taNAC2D displays opposite responses to abiotic stresses between seedling and mature stage of transgenic Arabidopsis. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef]
- Shamsudin, N.A.A.; Swamy, B.P.M.M.; Ratnam, W.; Cruz, M.T.S.; Sandhu, N.; Raman, A.K.; Kumar, A.; Abd, N.; Shamsudin, A.; Swamy, B.P.M.M.; et al. Pyramiding of drought yield QTLs into a high quality Malaysian rice cultivar MRQ74 improves yield under reproductive stage drought. Rice 2016, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.; Kolapo, K. Drought resistance in rice from conventional to molecular breeding: A review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef] [Green Version]
- Monneveux, P.; Jing, R.; Misra, S.C. Phenotyping for drought adaptation in wheat using physiological traits. Front. Physiol. 2012, 3, 429. [Google Scholar] [CrossRef] [Green Version]
- Passioura, J.B. Phenotyping for drought tolerance in grain crops: When is it useful to breeders? Funct. Plant Biol. 2012, 39, 851–859. [Google Scholar] [CrossRef]
- Lawas, L.M.F.; Shi, W.; Yoshimoto, M.; Hasegawa, T.; Hincha, D.K.; Zuther, E.; Jagadish, S.V.K. Combined drought and heat stress impact during flowering and grain filling in contrasting rice cultivars grown under field conditions. Field Crop. Res. 2018, 229, 66–77. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. Eight cycles of selection for drought tolerance in lowland tropical maize. II. Responses in reproductive behavior. Field Crop. Res. 1993, 31, 253–268. [Google Scholar] [CrossRef]
- Zhao, Y.; Karypis, G. Evaluation of hierarchical clustering algorithms for document datasets. In Proceedings of the Eleventh International Conference on Information and Knowledge Management, McLean, VA, USA, 4–9 November 2002; Association for Computing Machinery: New York, NY, USA, 2002; pp. 515–524. [Google Scholar]
- Kumar, S.S.; Dwivedi, S.K.; Basu, S.; Kumar, G.; Mishra, J.S.; Koley, T.K.; Rao, K.K.; Choudhary, A.K.; Mondal, S.; Kumar, S.S.; et al. Anatomical, agro-morphological and physiological changes in rice under cumulative and stage specific drought conditions prevailed in eastern region of India. Field Crop. Res. 2020, 245, 107658. [Google Scholar] [CrossRef]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.S.; Wu, B.; Wang, H.; Kang, D. Field screening of rice germplasm (Oryza sativa L. ssp. japonica) based on days to flowering for drought escape. Plants 2020, 9, 609. [Google Scholar] [CrossRef]
- Zu, X.; Lu, Y.; Wang, Q.; Chu, P.; Miao, W.; Wang, H.; La, H. A new method for evaluating the drought tolerance of upland rice cultivars. Crop. J. 2017, 5, 488–498. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, L.; Li, T.; Wu, H.; Cui, H. Breeding report on the new upland rice variety of Jiaohan No. 1 with high yield and good quality. J. Henan Agric. Sci. 2011, 40, 64–65. [Google Scholar]
- Gong, X.; Liu, X.; Pan, Q.; Mi, G.; Chen, F.; Yuan, L. Combined physiological, transcriptome and genetic analysis reveals a molecular network of nitrogen remobilization in maize. J. Exp. Bot. 2020, 71, 5061–5073. [Google Scholar] [CrossRef]
- Buckler, E.S.; Holland, J.B.; Bradbury, P.J.; Acharya, C.B.; Brown, P.J.; Browne, C.; Ersoz, E.; Flint-Garcia, S.; Garcia, A.; Glaubitz, J.C.; et al. The genetic architecture of maize flowering time. Science 2009, 325, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Kump, K.L.; Bradbury, P.J.; Wisser, R.J.; Buckler, E.S.; Belcher, A.R.; Oropeza-Rosas, M.A.; Zwonitzer, J.C.; Kresovich, S.; McMullen, M.D.; Ware, D.; et al. Genome-wide association study of quantitative resistance to southern leaf blight in the maize nested association mapping population. Nat. Genet. 2011, 43, 163–168. [Google Scholar] [CrossRef]
- Tian, F.; Bradbury, P.J.; Brown, P.J.; Hung, H.; Sun, Q.; Flint-Garcia, S.; Rocheford, T.R.; Mcmullen, M.D.; Holland, J.B.; Buckler, E.S. Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat. Genet. 2011, 43, 159–162. [Google Scholar] [CrossRef]
- Huang, C.; Chen, Q.; Xu, G.; Xu, D.; Tian, J.; Tian, F. Identification and fine mapping of quantitative trait loci for the number of vascular bundle in maize stem. J. Integr. Plant Biol. 2016, 58, 81–90. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Volume 2. [Google Scholar]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. 2020. [Google Scholar]
- Aravind, J.; Mukesh, S.S.; Wankhede, D.P.; Kaur, V. AugmentedRCBD: Analysis of Augmented Randomized Complete Block Designs. 2020. [Google Scholar]
- IRRI. Standard Evaluation System for Rice, 5th ed.; International Rice Research Institute: Manila, Philippines, 2013; ISBN 9781612099897. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; ISBN 9783319242750. [Google Scholar]
- Kato, Y.; Katsura, K. Rice adaptation to aerobic soils: Physiological considerations and implications for agronomy. Plant Prod. Sci. 2014, 17, 1–12. [Google Scholar] [CrossRef]
- Murtagh, F.; Legendre, P. Ward’s hierarchical agglomerative clustering method: Which algorithms implement Ward’s criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q. Strategies for developing green super rice. Proc. Natl. Acad. Sci. USA 2007, 95, 1663–1668. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Caldo, R.; Sebastian, L.; Hernandez, J. Morphology-based genetic diversity analysis of ancestral lines of Philippine rice cultivars. Philipp. J. Crop. Sci. 1996, 21, 86–92. [Google Scholar]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajracharya, J.; Steele, K.A.; Jarvis, D.I.; Sthapit, B.R.; Witcombe, J.R. Rice landrace diversity in Nepal: Variability of agro-morphological traits and SSR markers in landraces from a high-altitude site. Field Crop. Res. 2006, 95, 327–335. [Google Scholar] [CrossRef]
- Yawen, Z.; Shiquan, S.; Zichao, L.; Zhongyi, Y.; Xiangkun, W.; Hongliang, Z.; Guosong, W. Ecogeographic and genetic diversity based on morphological characters of indigenous rice (Oryza sativa L.) in Yunnan, China. Genet. Resour. Crop. Evol. 2003, 50, 567–577. [Google Scholar] [CrossRef]
- Huang, M.; Xu, Y.; Wang, H. Field identification of morphological and physiological traits in two special mutants with strong tolerance and high sensitivity to drought stress in upland rice (Oryza sativa L.). J. Integr. Agric. 2019, 18, 970–981. [Google Scholar] [CrossRef]
- Mahalle, M.D.; Dey, P.C.; Chetia, S.K.; Baruah, A.R.; Ahmed, T.; Sarma, R.N.; Kaldate, R.C.; Kumar, A.; Singh, S.K.; Modi, M.K. Association mapping for yield traits under drought stress in Autumn rice germplasm collection of Assam. J. Plant Biochem. Biotechnol. 2020, 30, 26–36. [Google Scholar] [CrossRef]
- Bänziger, M.; Edmeades, G.O.; Beck, D.; Bellon, M. Secondary traits. In Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice; CIMMYT: Texcoco, Mexico, 2000; pp. 39–46. ISBN 970-648463. [Google Scholar]
- Araus, J.L.; Slafer, G.A.; Reynolds, M.P. Plant breeding and drought in C3 cereals: What should we breed for? Ann. Bot. 2002, 89, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Piveta, L.B.; Roma-Burgos, N.; Noldin, J.A.; Viana, V.E.; de Oliveira, C.; Lamego, F.P.; Avila, L.A. de Molecular and physiological responses of rice and weedy rice to heat and drought stress. Agriculture 2020, 11, 9. [Google Scholar] [CrossRef]
- Torres, R.O.; Mcnally, K.L.; Cruz, C.V.; Serraj, R.; Henry, A. Screening of rice genebank germplasm for yield and selection of new drought tolerance donors. Field Crop. Res. 2013, 147, 12–22. [Google Scholar] [CrossRef]
- Royo, C.; Álvaro, F.; Martos, V.; Ramdani, A.; Isidro, J.; Villegas, D.; García Del Moral, L.F. Genetic changes in durum wheat yield components and associated traits in Italian and Spanish varieties during the 20th century. Euphytica 2007, 155, 259–270. [Google Scholar] [CrossRef]
- Slafer, G.A.; Araus, J.L.J.L.; Royo, C.; García, L.F.; Moral, D.E.L.; Moral, L.F.G. Del Promising eco-physiological traits for genetic improvement of cereal yields in Mediterranean environments. Ann. Appl. Biol. 2005, 146, 61–70. [Google Scholar] [CrossRef]
- Lanna, A.C.; Coelho, G.R.C.; Moreira, A.S.; Terra, T.G.R.; Brondani, C.; Saraiva, G.R.; Lemos, F.D.S.; Guimarães, P.H.R.; Morais, O.P.; Vianello, R.P. Upland rice: Phenotypic diversity for drought tolerance. Sci. Agric. 2020, 78, 1–14. [Google Scholar]
- Bolaños, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crop. Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
- Arai-Sanoh, Y.; Ishimaru, T.; Ohsumi, A.; Kondo, M. Effects of soil temperature on growth and root function in rice. Plant Prod. Sci. 2010, 13, 235–242. [Google Scholar] [CrossRef]
- Rosielle, A.A.; Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environment. Crop. Sci. 1981, 21, 943–946. [Google Scholar] [CrossRef]
- Blum, A. Plant breeding and yield stability. In Plant Breeding For Stress Environments; CRC Press: Boca Raton, FL, USA, 1988; pp. 15–42. ISBN 9781315896618. [Google Scholar]
- Edmeades, G.O.; Bolaños, J.; Lafitte, H.R.; Rajaram, S.; Pfeiffer, W.; Fischer, R.A. Traditional approaches to breeding for drought resistance in cereals. In Drought Resistance in Cereals; Baker, F.W.G., Ed.; CAB International: Wallingford, UK, 1989; pp. 27–52. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
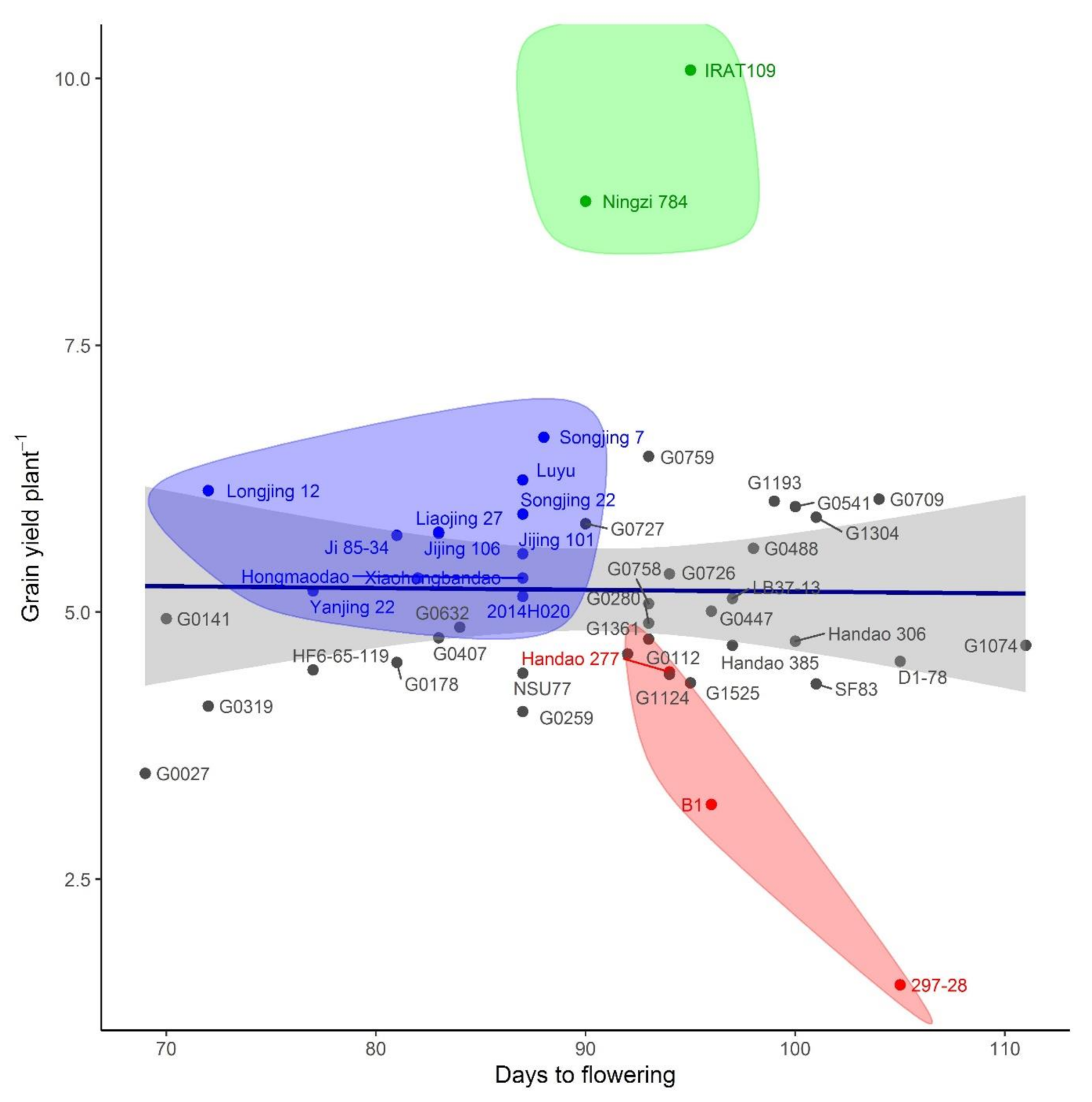{kind=link}
| Ecotype | Lowland | Upland | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Origin | Exotic | Line | Variety | Cultivar | Line | Variety | Cultivar | ||
| China | 195 | 1351 | 203 | 31 | 28 | 92 | 1900 | ||
| Anhui | 8 | 8 | |||||||
| Beijing | 37 | 31 | 8 | 76 | |||||
| Guizhou | 6 | 6 | |||||||
| Hebei | 31 | 20 | 3 | 54 | |||||
| Heilongjiang | 192 | 37 | 2 | 13 | 244 | ||||
| Henan | 67 | 4 | 71 | ||||||
| Inner Mongolia | 4 | 4 | 8 | ||||||
| Jiangsu | 176 | 150 | 1 | 1 | 328 | ||||
| Jilin | 263 | 51 | 1 | 15 | 330 | ||||
| Liaoning | 247 | 41 | 12 | 36 | 336 | ||||
| Ningxia | 85 | 13 | 98 | ||||||
| Shandong | 19 | 60 | 25 | 16 | 120 | ||||
| Tianjin | 2 | 2 | 4 | ||||||
| Xinjiang | 17 | 9 | 3 | 29 | |||||
| Yunnan | 188 | 188 | |||||||
| Brazil | 1 | 1 | |||||||
| Ivory Coast | 2 | 2 | |||||||
| Japan | 56 | 1 | 57 | ||||||
| South Korea | 70 | 70 | |||||||
| Total | 126 | 195 | 1352 | 203 | 30 | 31 | 92 | 2030 | |
| Trait | Description |
|---|---|
| DF | Days to flowering was recorded as the number of days from sowing to the time when inflorescences had emerged above the flag leaf sheath for more than half of the individuals of a landrace. |
| PHL (cm) | Measured height from ground to the highest leaf tip with a meter rod. |
| PHP (cm) | Measured height from ground to panicle tip with a meter rod. |
| ABP−1 (g) | Shoot dry weight, including the grain yield and straw, were weighed for each plant after being dried in an oven at 105 °C for 30 min and at 80 °C for 23.5 h. |
| GYP−1 (g) | Total grain weight plant−1 was weighed after drying at 105 °C for 30 min and then 80 °C for 23.5 h in an oven. |
| HI | The harvest index was computed as the ratio of filled spikelet weight to total aboveground biomass. |
| Trait | G | C | Y | G × C | G × Y | C × Y | G × C × Y |
|---|---|---|---|---|---|---|---|
| DF | 0.000 | 0.000 | 0.000 | 0.000 | 0.012 | 0.000 | 0.979 |
| PHL | 0.000 | 0.000 | 0.997 | 0.002 | 0.015 | 0.000 | 0.535 |
| PHP | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.004 |
| ABP−1 | 0.000 | 0.000 | 0.006 | ||||
| GYP−1 | 0.000 | 0.000 | 0.001 | ||||
| HI | 0.000 | 0.000 | 0.221 |
| Factor | DF | PHL (cm) | PHP (cm) | ABP −1 (g) | GYP −1 (g) | HI |
|---|---|---|---|---|---|---|
| Genotype | 100.07 ± 20.88 | 91.52 ± 19.10 | 88.21 ± 18.28 | 11.09 ± 5.79 | 4.17 ± 2.61 | 0.372 ± 0.11 |
| Control | 98.69 ± 8.47 | 93.44 ± 18.37 | 89.15 ± 19.61 | 11.09 ± 4.66 | 4.06 ± 2.3 | 0.348 ± 0.12 |
| B1 | 96.04 ± 6.53 | 75.34 ± 7.86 | 68.12 ± 7.61 | 9.48 ± 2.84 | 3.49 ± 1.4 | 0.36 ± 0.07 |
| HD-277 | 94.39 ± 5.67 | 96.41 ± 10.37 | 94.68 ± 10.33 | 12.88 ± 4.44 | 5.52 ± 1.97 | 0.43 ± 0.07 |
| 297-28 | 106.2 ± 7.82 | 109.24 ± 15.83 | 105.3 ± 15.89 | 11 ± 5.53 | 3.24 ± 2.6 | 0.26 ± 0.12 |
| Condition (C) | ||||||
| Lowland | 99.17 ± 17.94 | 98.62 ± 16.82 | 95.63 ± 15.93 | 13.93 ± 6.04 | 5.39 ± 2.82 | 0.39 ± 0.11 |
| Upland | 100.27 ± 21.02 | 84.29 ± 14.24 | 80.77 ± 13.09 | 7.99 ± 3.77 | 2.83 ± 1.55 | 0.36 ± 0.11 |
| Year (Y) | ||||||
| 2017 | 103.8 ± 20.8 | 91.02 ± 19.96 | 88.57 ± 19.3 | |||
| 2018 | 96.05 ± 19.63 | 92.18 ± 18.13 | 87.94 ± 17.35 | 11.09 ± 5.75 | 4.16 ± 2.6 | 0.371 ± 0.11 |
| HSD0.05 | ||||||
| Sc | 7.00 | 7.11 | 7.21 | 4.82 | 2.17 | 0.11 |
| Sb | 55.96 | 64.45 | 65.29 | 30.63 | 13.77 | 0.60 |
| Sv | 68.54 | 74.43 | 75.43 | 35.35 | 15.94 | 0.65 |
| Svc | 34.53 | 37.44 | 37.94 | 17.90 | 8.05 | 0.45 |
| C | 0.276 | 0.409 | 0.379 | 0.267 | 0.118 | 0.0055 |
| Y | 0.276 | NS | 0.378 | |||
| C × Y | 0.553 | 0.821 | 0.758 |
| Trait | Year | C | Mean | Min | Max | CV (%) | SEM | SD (±) | GCV (%) | PCV (%) | H2 (%) | GA (%) | HSD0.05 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Days to flowering | 2017 | L | 104.1 | 67 | 174 | 9.23 | 0.44 | 19.73 | 16.36 | 18.78 | 75.91 | 30.63 | 50.16 |
| U | 101.9 | 61 | 170 | 4.96 | 0.51 | 21.84 | 20.76 | 21.36 | 94.52 | 42.43 | 31.41 | ||
| 2018 | L | 94.24 | 67 | 138 | 3.93 | 0.37 | 16.16 | 18.6 | 19.01 | 95.72 | 35.38 | 18.90 | |
| U | 96.92 | 55 | 155 | 6.19 | 0.47 | 19.64 | 21.63 | 22.51 | 92.29 | 41.54 | 29.37 | ||
| Plant height to leaf (cm) | 2017 | L | 99.67 | 43.8 | 187 | 6.82 | 0.5 | 19.43 | 17.45 | 18.74 | 86.64 | 33.39 | 35.90 |
| U | 81 | 33 | 149.33 | 8.77 | 0.43 | 16.47 | 19.41 | 21.35 | 82.63 | 29.48 | 35.28 | ||
| 2018 | L | 97.2 | 42 | 178.33 | 7.09 | 0.41 | 16.94 | 18.6 | 19.9 | 87.36 | 34.85 | 34.95 | |
| U | 85.84 | 45 | 139.67 | 9.56 | 0.42 | 15 | 27.03 | 31.76 | 72.43 | 2.27 | 39.15 | ||
| Plant height to panicle (cm) | 2017 | L | 97.62 | 44 | 191 | 6.55 | 0.48 | 18.59 | 16.92 | 18.15 | 86.93 | 31.77 | 33.68 |
| U | 78.2 | 25.6 | 147.67 | 8.68 | 0.4 | 15.57 | 18.91 | 20.86 | 82.17 | 27.66 | 33.70 | ||
| 2018 | L | 93.25 | 54 | 185 | 6.84 | 0.4 | 16.39 | 18.58 | 19.79 | 88.15 | 33.56 | 32.30 | |
| U | 82.41 | 45 | 139.67 | 9.27 | 0.38 | 14.1 | 13.17 | 16.07 | 67.12 | 18.34 | 36.36 | ||
| Aboveground biomass plant −1 (g) | L | 13.93 | 2.50 | 48.66 | 33.58 | 0.15 | 6.04 | 24.81 | 41.83 | 35.18 | 4.23 | 24.05 | |
| U | 7.99 | 1.22 | 26.47 | 28.19 | 0.1 | 3.77 | 32.36 | 42.28 | 58.57 | 4.08 | 10.48 | ||
| Grain yield plant −1 (g) | L | 5.39 | 0.1 | 17.36 | 36.1 | 0.07 | 2.82 | 34.52 | 50.22 | 47.26 | 2.64 | 10.07 | |
| U | 2.83 | 0.05 | 10.65 | 39.18 | 0.04 | 1.55 | 33.08 | 49.58 | 44.5 | 1.29 | 5.03 | ||
| Harvest index | L | 0.39 | 0.01 | 0.69 | 15.36 | 0.002 | 0.11 | 22.23 | 27.07 | 67.43 | 0.15 | 0.31 | |
| U | 0.36 | 0.01 | 0.65 | 24.6 | 0.003 | 0.11 | 16.22 | 28.99 | 31.28 | 0.07 | 0.48 |
| Variable | Factor | DF | PHL | PHP | ABP−1 | GYP−1 | HI |
|---|---|---|---|---|---|---|---|
| RDS | 2017 | 2.12 ± 7.89 NS | 17.52 ± 12.69 *** | 18.32 ± 13.31 *** | |||
| 2018 | −3.32 ± 8.59 *** | 9.53 ± 12.82 *** | 10.3 ± 13.21 *** | 42.72 ± 27.91 *** | 52.45 ± 26.57 *** | 16.56 ± 26.44 *** | |
| Pooled | −0.3 ± 8.25 *** d | 13.82 ± 12.76 *** c | 14.39 ± 13.26 *** c | 42.78 ± 27.91 *** b | 52.34 ± 26.57 *** a | 16.19 ± 26.44 *** c | |
| Weighted Factor Calculation | Shared RDS variation (%) | 0.21 | 9.89 | 10.29 | 30.60 | 37.44 | 11.58 |
| Original ranking | 6 | 5 | 4 | 2 | 1 | 3 | |
| Practical ranking | 5 | 2 | 4 | 1 | 3 | 6 | |
| Weighted factor (w; %) | 9.89 | 30.60 | 10.29 | 37.44 | 11.58 | 0.21 | |
| Minimum | 2017 | −56.16 | −49.46 | −65.50 | |||
| 2018 | −60.22 | −61.90 | −80.69 | −66.06 | −120.60 | −111.98 | |
| Pooled | −60.22 | −61.90 | −80.69 | −66.06 | −120.60 | −110.98 | |
| Maximum | 2017 | 53.59 | 67.28 | 74.54 | |||
| 2018 | 29.63 | 46.84 | 52.42 | 91.75 | 98.73 | 98.60 | |
| Pooled | 30.67 | 57.00 | 49.96 | 91.75 | 98.73 | 98.60 | |
| Range | 2017 | 109.76 | 116.74 | 140.03 | |||
| 2018 | 89.84 | 108.75 | 133.11 | 157.81 | 219.33 | 210.57 | |
| Pooled | 90.88 | 118.90 | 130.65 | 157.81 | 219.33 | 209.57 | |
| Variance (s2) | 2017 | 62.21 | 161.13 | 177.07 | |||
| 2018 | 73.87 | 164.43 | 174.54 | 779.19 | 705.76 | 699.27 | |
| Pooled | 68.04 | 162.78 | 175.81 | 779.19 | 705.76 | 699.27 | |
| Standard Deviation (SD) | 2017 | 7.89 | 12.69 | 13.31 | |||
| 2018 | 8.59 | 12.82 | 13.21 | 27.91 | 26.57 | 26.44 | |
| Pooled | 8.25 | 12.76 | 13.26 | 27.91 | 26.57 | 26.44 |
| DRG | G. N. | DF | PHL | PHP | ABP⁻¹ | GYP⁻¹ | HI | Mean | Cluster | Class | Ecotype | Origin a |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 0.91 | 2.08 | 1.16 | −22.53 | −10.50 | −0.037 | −28.92 | VII | A | RU | North, Ivory Coast |
| 1–3 | 5 | 0.06 | −1.78 | 0.03 | −14.44 | −2.25 | 0.025 | −18.36 | VI, VII | A | North | |
| 3 | 18 | 0.05 | 1.49 | 0.59 | −10.72 | −0.27 | 0.035 | −8.81 | VI, VII, VIII, IX | A, B | TL, IU, TU | North, Centre, South |
| 3–5 | 67 | 0.20 | 2.25 | 0.96 | −2.76 | 1.73 | 0.039 | 2.42 | V, VI, VII, VIII, IX | A, B, C | TL, IU, RU, TU | North, Centre, South, South Korea, Japan |
| 5 | 141 | 0.12 | 3.12 | 1.09 | 4.70 | 2.93 | 0.025 | 11.98 | V, VI, VII, VIII, IX | A, B, C | TL, IU, RU, TU | North, Centre, South, South Korea, Japan |
| 5–7 | 250 | 0.04 | 3.45 | 1.18 | 12.64 | 4.93 | 0.023 | 22.27 | I, II, III, V, VI, VII, VIII, IX | A, B, C, D | TL, IU, RU, TU | North, Centre, South, South Korea, Japan |
| 7 | 328 | −0.07 | 4.39 | 1.48 | 19.54 | 7.00 | 0.030 | 32.36 | I, II, III, IV, V, VI, VII, VIII, IX | A, B, C, D | TL, IU, RU, TU | North, Centre, South, South Korea, Japan |
| 7–9 | 247 | −0.14 | 5.46 | 1.93 | 25.95 | 8.82 | 0.041 | 42.06 | I, II, III, IV, V, VIII | B, C, D | TL, IU, RU, TU | North, Centre, South, South Korea, Japan, Brazil |
| 9 | 60 | −0.29 | 7.90 | 2.87 | 29.69 | 10.02 | 0.064 | 50.26 | I, IV, V | C, D | TL, RU, TU | North, Centre, South, South Korea, Japan |
| SD | 0.32 | 2.55 | 0.76 | 17.51 | 5.99 | 0.03 | ||||||
| SEM | 0.11 | 0.85 | 0.25 | 5.84 | 2.00 | 0.01 |
| Cluster | G. N. | Statistic | DF | PHL | PHP | ABP−1 | GYP−1 | HI | Total | RDS-I Performance | Class | DRG with G. (%) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VI | 113 | Mean | −0.14 | 2.12 | 0.81 | 6.18 | 5.57 | 0.08 | 14.62 | Dry biomass (ABP−1 and GYP−1) and plant height (PHL and PHP) were lowest or lower, thus genotypes have strong drought resistance (all the genotypes in top DRGs 1 and 1–3 and the majority of them from DRGs 3, 3–5 and 5 fall in these clusters) | A | 1 | 100 |
| Min | −2.42 | −6.51 | −2.85 | −23.28 | −1.91 | 0.03 | −22.77 | 1–3 | 100 | ||||
| Max | 2.6 | 5.5 | 2.14 | 16.3 | 11 | 0.18 | 31.66 | 3 | 77.78 | ||||
| VII | 176 | Mean | 0.21 | 1.76 | 0.58 | 7.42 | 2.20 | 0.00 | 12.18 | 3–5 | 68.66 | ||
| Min | −1.04 | −13.18 | −2.43 | −24.71 | −13.99 | −0.11 | −35.85 | 5 | 65.25 | ||||
| Max | 1.9 | 8.42 | 2.58 | 17.39 | 6.07 | 0.09 | 32.91 | 5–7 a | 47.2 | ||||
| VIII | 97 | Mean | 0.00 | 6.13 | 2.07 | 8.88 | 3.05 | 0.01 | 20.13 | Comparatively, dry biomass was lower but plant height was higher. Hence, drought resistance of genotypes varied with moderate (DRG 5) to somewhat strong resistance (DRG 3–5 and 3) | B | 3 | 22.22 |
| Min | −3.41 | 2.41 | 0.86 | −10.79 | −2.29 | −0.07 | −5.2 | 3–5 | 29.85 | ||||
| Max | 1.93 | 17.44 | 4.36 | 15.88 | 5.87 | 0.03 | 37.87 | 5 | 34.04 | ||||
| IX | 76 | Mean | 0.10 | 5.83 | 2.02 | 1.56 | 3.85 | 0.06 | 13.42 | 5–7 | 33.2 | ||
| Min | −1.15 | 3.64 | 0.86 | −19.81 | −0.09 | 0.02 | −10.82 | 7 | 5.18 | ||||
| Max | 3.04 | 12.81 | 4.71 | 12.92 | 6.62 | 0.13 | 30.25 | 7–9 | 0.4 | ||||
| I | 117 | Mean | −0.05 | 2.69 | 1.04 | 22.90 | 8.51 | 0.06 | 35.16 | Contrary to class B above, dry biomass and plant height were higher and lower, respectively. Therefore, drought resistance of the majority of genotypes was weak (DRG 7) | C | ||
| Min | −1.96 | −2.58 | −1.69 | 16.36 | 6.28 | −0.19 | 20.16 | 5–7 | 13.6 | ||||
| Max | 2.33 | 5.91 | 2.01 | 33.22 | 11.36 | 0.18 | 48.45 | 7 | 46.04 | ||||
| II | 126 | Mean | 0.06 | 2.00 | 0.65 | 22.31 | 6.60 | −0.01 | 31.61 | 7–9 | 22.67 | ||
| Min | −1.55 | −3.94 | −1.90 | 16.00 | 1.99 | −0.17 | 20.18 | 9 | 3.33 | ||||
| Max | 2.09 | 6.91 | 1.96 | 31.61 | 10.00 | 0.03 | 45.33 | ||||||
| III | 69 | Mean | −0.08 | 6.42 | 1.96 | 19.44 | 5.60 | −0.02 | 33.33 | Both dry biomass and plant height were highest or higher (highly drought susceptible); thus, drought resistance of genotypes was weak to weakest (DRG 7 to 9) | D | ||
| Min | −1.47 | 4.44 | 0.4 | 16.03 | 0.95 | −0.14 | 24.14 | 3–5 | 1.49 | ||||
| Max | 1.58 | 16.44 | 4.02 | 24.27 | 7.22 | 0.03 | 47.79 | 5 | 0.71 | ||||
| IV | 113 | Mean | −0.08 | 6.23 | 2.03 | 26.68 | 8.24 | 0.00 | 43.10 | 5–7 | 6 | ||
| Min | −3.3 | 4.21 | −0.14 | 20.85 | 6.33 | −0.12 | 34.06 | 7 | 45.12 | ||||
| Max | 2.44 | 10.89 | 3.8 | 34.31 | 10.6 | 0.05 | 58.52 | 7–9 | 76.92 | ||||
| V | 231 | Mean | −0.22 | 6.22 | 2.35 | 22.07 | 8.87 | 0.08 | 39.38 | 9 | 96.67 | ||
| Min | −4.11 | 2.69 | 1.19 | −16.46 | 6.17 | 0.04 | 4.99 | ||||||
| Max | 2.07 | 14.2 | 5.15 | 33.48 | 11.45 | 0.2 | 55.38 | ||||||
| SD | 0.13 | 2.14 | 0.71 | 9.18 | 2.44 | 0.04 | |||||||
| SEM | 0.04 | 0.71 | 0.24 | 3.06 | 0.81 | 0.01 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, M.S.; Wu, B.; Wang, H.; Kang, D. Identification of Drought Tolerance on the Main Agronomic Traits for Rice (Oryza sativa L. ssp. japonica) Germplasm in China. Agronomy 2021, 11, 1740. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091740
Ahmad MS, Wu B, Wang H, Kang D. Identification of Drought Tolerance on the Main Agronomic Traits for Rice (Oryza sativa L. ssp. japonica) Germplasm in China. Agronomy. 2021; 11(9):1740. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091740
Chicago/Turabian StyleAhmad, Muhammad Shafiq, Bingrui Wu, Huaqi Wang, and Dingming Kang. 2021. "Identification of Drought Tolerance on the Main Agronomic Traits for Rice (Oryza sativa L. ssp. japonica) Germplasm in China" Agronomy 11, no. 9: 1740. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091740