
Genetic Mapping Reveals Novel Exotic and Elite QTL Alleles for Salinity Tolerance in Barley

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Genotyping
2.2. Experimental Site, Design, and Trial Management
2.3. Data Collection
2.4. Data Analysis
3. Results
3.1. Phenotypic Variations and Heritability Estimates
3.2. Identification of QTL
3.3. Co-Location of QTL under Salinity Conditions
4. Discussion
4.1. The Barley Population’s Wild Genetic Background and Salinity Tolerance
4.2. Stress Tolerance Index and Heritability
4.3. Candidate Genes with Potential Functions and Corresponding QTL under Salinity Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, H.; Shan, W.; Zhao, J.; Zhang, G.; Li, C.; Wu, F. Difference in response to aluminum stress among Tibetan wild barley genotypes. J. Plant Nutr. Soil Sci. 2011, 174, 952–960. [Google Scholar] [CrossRef]
- Islam, F.; Wang, J.; Farooq, M.A.; Yang, C.; Jan, M.; Mwamba, T.M.; Hannan, F.; Xu, L.; Zhou, W. Rice Responses and Tolerance to Salt Stress: Deciphering the Physiological and Molecular Mechanisms of Salinity Adaptation; Woodhead Publishing: Cambridge, UK; Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780128143322. [Google Scholar]
- Hassani, A.; Azapagic, A.; Shokri, N. Predicting long-term dynamics of soil salinity and sodicity on a global scale. Proc. Natl. Acad. Sci. USA 2021, 117, 33017–33027. [Google Scholar] [CrossRef] [PubMed]
- Sanower Hossain, M.; Sultan Ahmad Shah, J. Present Scenario of Global Salt Affected Soils, its Management and Importance of Salinity Research. Int. Res. J. Biol. Sci. Perspect. 2019, 1, 2663–5976. [Google Scholar]
- Abdel-Shafy, H.I.; Kamel, A.H. Groundwater in Egypt issue: Resources, location, amount, contamination, protection, renewal, future overview. Egypt. J. Chem. 2016, 59, 321–362. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Zhang, Y.; Sun, J.; Shao, H. Negative interactive effects between biochar and phosphorus fertilization on phosphorus availability and plant yield in saline sodic soil. Sci. Total Environ. 2016, 568, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Noaman, M.M.; El-Banna, M.N.; El-Gawad Nassar, M.A.A.; Boseely, M.A.E.-A. Evaluation of 16 Barley Genotypes under Calcareous Soil Conditions in Egypt. J. Agric. Sci. 2011, 3, 105. [Google Scholar] [CrossRef] [Green Version]
- Ouda, S.; Ewise, M.; Noreldin, T. Projection of productivity of cultivated crops in rain-fed areas in Egypt under climate change. Cogent Food Agric. 2016, 2, 1136256. [Google Scholar] [CrossRef]
- Dai, F.; Nevo, E.; Wu, D.; Comadran, J.; Zhou, M.; Qiu, L.; Chen, Z.; Beiles, A.; Chen, G.; Zhang, G. Tibet is one of the centers of domestication of cultivated barley. Proc. Natl. Acad. Sci. USA 2012, 109, 16969–16973. [Google Scholar] [CrossRef] [Green Version]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World: The Origin and Spread of Cultivated Plants in West Asia, Europe, and the Nile Valley; OUP E-Books; Oxford University Press: New York, NY, USA, 2000; ISBN 9780198503576. [Google Scholar]
- Qiu, L.; Wu, D.; Ali, S.; Cai, S.; Dai, F.; Jin, X.; Wu, F.; Zhang, G. Evaluation of salinity tolerance and analysis of allelic function of HvHKT1 and HvHKT2 in Tibetan wild barley. Theor. Appl. Genet. 2011, 122, 695–703. [Google Scholar] [CrossRef]
- Ashraf, M.Y.; Akhter, K.; Hussain, F.; Iqbal, J. Screening of different accessions of three potential grass species from cholistan desert for salt tolerance. Pakistan J. Bot. 2006, 38, 1589–1597. [Google Scholar]
- Allel, D.; Benamar, A.; Badri, M.; Abdelly, C. Evaluation of salinity tolerance indices in north african barley accessions at reproductive stage. Czech J. Genet. Plant Breed. 2019, 55, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.J.; Tucker, E.J.; Tester, M. Genetic analysis of abiotic stress tolerance in crops. Curr. Opin. Plant Biol. 2011, 14, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Shavrukov, Y.; Bovill, J.; Afzal, I.; Hayes, J.E.; Roy, S.J.; Tester, M.; Collins, N.C. HVP10 encoding V-PPase is a prime candidate for the barley HvNax3 sodium exclusion gene: Evidence from fine mapping and expression analysis. Planta 2013, 237, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Mano, Y.; Takeda, K. Mapping quantitative trait loci for salt tolerance at germination and the seedling stage in barley (Hordeum vulgare L.). Euphytica 1997, 94, 263–272. [Google Scholar] [CrossRef]
- Von Korff, M.; Wang, H.; Léon, J.; Pillen, K. Development of candidate introgression lines using an exotic barley accession (Hordeum vulgare ssp. spontaneum) as donor. Theor. Appl. Genet. 2004, 109, 1736–1745. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Schmalenbach, I.; von Korff, M.; Léon, J.; Kilian, B.; Rode, J.; Pillen, K. Association of barley photoperiod and vernalization genes with QTLs for flowering time and agronomic traits in a BC2DH population and a set of wild barley introgression lines. Theor. Appl. Genet. 2010, 120, 1559–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, M.A.; Schumann, H.; Pillen, K.; Naz, A.A.; Léon, J. AB-QTL analysis reveals new alleles associated to proline accumulation and leaf wilting under drought stress conditions in barley (Hordeum vulgare L.). BMC Genet. 2012, 13, 61. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, E.S.A.; El-Sobky, E.S.E.A.; Farag, H.I.A.; Yasin, M.A.T.; Attia, A.; Rady, M.O.A.; Awad, M.F.; Mansour, E. Sowing date and genotype influence on yield and quality of dual-purpose barley in a salt-affected arid region. Agronomy 2021, 11, 717. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall Inc.: Englewood Cliffs, NJ, USA, 1958; Volume 498, pp. 183–204. [Google Scholar]
- Zhang, H.; Schroder, J.L.; Pittman, J.J.; Wang, J.J.; Payton, M.E. Soil Salinity Using Saturated Paste and 1:1 Soil to Water Extracts. Soil Sci. Soc. Am. J. 2005, 69, 1146–1151. [Google Scholar] [CrossRef]
- SAS Institute. The SAS System for Windows; Release 9.2; SAS Inst.: Cary, NC, USA, 2011; Available online: http://www.sciepub.com/reference/166089 (accessed on 4 July 2021).
- Padi, F.K. Genotype × environment interaction and yield stability in a cowpea-based cropping system. Euphytica 2007, 158, 11–25. [Google Scholar] [CrossRef]
- Singh, R.K.; Chaudhary, B.D. Biometrical Methods in Quantitative Genetic Analysis; Kalyani Publishers: Ludhiana, India, 1979. [Google Scholar]
- Fernandez, G.C.J. Effective selection criteria for assessing plant stress tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and other Food Crops in Temperature and Water Stress, Shanhua, Taiwan, 13–16 August 1992; pp. 257–270. [Google Scholar] [CrossRef]
- Payne, R.W.; Murray, D.A.; Harding, S.A.; Baird, D.B.; Soutar, D.M. Genstat® for WindowsTM, 15th ed.; VSN International Ltd.: Oxford, UK, 2012. [Google Scholar]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Lacolla, G.; Cucci, G. Reclamation of sodic-saline soils. Barley crop response. Ital. J. Agron. 2008, 3, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Harlan, J.R.; Zohary, D. Distribution of Wild Wheats and Barley. Science 1966, 153, 1074–1080. [Google Scholar] [CrossRef]
- Nevo, E.; Chen, G. Drought and salt tolerances in wild relatives for wheat and barley improvement. Plant Cell Environ. 2010, 33, 670–685. [Google Scholar] [CrossRef]
- Sayed, M.A.; Hamada, A.; Lèon, J.; Naz, A.A. Genetic mapping reveals novel exotic QTL alleles for seminal root architecture in barley advanced backcross double haploid population. Euphytica 2017, 213, 1–16. [Google Scholar] [CrossRef]
- Abd El-Aty, M.; Amer, K.; Eldegwy, I.; El-Akhdar, A. Genetic Studies on Yield and Its Components in Some Barley Crosses. J. Plant Prod. 2011, 2, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Saade, S.; Brien, C.; Pailles, Y.; Berger, B.; Shahid, M.; Russell, J.; Waugh, R.; Negrão, S.; Tester, M. Dissecting new genetic components of salinity tolerance in two-row spring barley at the vegetative and reproductive stages. PLoS ONE 2020, 15, e0236037. [Google Scholar] [CrossRef]
- Jabbari, M.; Fakheri, B.A.; Aghnoum, R.; Darvishzadeh, R.; Mahdi Nezhad, N.; Ataei, R.; Koochakpour, Z.; Razi, M. Association analysis of physiological traits in spring barley (Hordeum vulgare L.) under water-deficit conditions. Food Sci. Nutr. 2021, 9, 1761–1779. [Google Scholar] [CrossRef]
- Thabet, S.G.; Moursi, Y.S.; Sallam, A.; Karam, M.A.; Alqudah, A.M. Genetic associations uncover candidate SNP markers and genes associated with salt tolerance during seedling developmental phase in barley. Environ. Exp. Bot. 2021, 188, 104499. [Google Scholar] [CrossRef]
- Mwando, E.; Angessa, T.T.; Han, Y.; Zhou, G.; Li, C. Quantitative Trait Loci Mapping for Vigour and Survival Traits of Barley Seedlings after Germinating under Salinity Stress. Agronomy 2021, 11, 103. [Google Scholar] [CrossRef]
- Mwando, E.; Han, Y.; Angessa, T.T.; Zhou, G.; Hill, C.B.; Zhang, X.Q.; Li, C. Genome-Wide Association Study of Salinity Tolerance During Germination in Barley (Hordeum vulgare L.). Front. Plant Sci. 2020, 11, 212–226. [Google Scholar] [CrossRef] [Green Version]
- Hordeum_Vulgare–Ensembl Genomes 51. Available online: https://plants.ensembl.org/Hordeum_vulgare/Info/Index (accessed on 1 August 2021).
- Oyiga, B.C.; Palczak, J.; Wojciechowski, T.; Lynch, J.P.; Naz, A.A.; Léon, J.; Ballvora, A. Genetic components of root architecture and anatomy adjustments to water-deficit stress in spring barley. Plant Cell Environ. 2020, 43, 692–711. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-X.; Blumwald, E. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 2001, 19, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Remy, E.; Niño-González, M.; Godinho, C.P.; Cabrito, T.R.; Teixeira, M.C.; Sá-Correia, I.; Duque, P. Heterologous expression of the yeast Tpo1p or Pdr5p membrane transporters in Arabidopsis confers plant xenobiotic tolerance. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Niño-González, M.; Novo-Uzal, E.; Richardson, D.N.; Barros, P.M.; Duque, P. More Transporters, More Substrates: The Arabidopsis Major Facilitator Superfamily Revisited. Mol. Plant 2019, 12, 1182–1202. [Google Scholar] [CrossRef] [Green Version]
- Vysotskaya, L.; Hedley, P.E.; Sharipova, G.; Veselov, D.; Kudoyarova, G.; Morris, J.; Jones, H.G. Effect of salinity on water relations of wild barley plants differing in salt tolerance. AoB Plants 2010, 2010, plq006. [Google Scholar] [CrossRef]
- Fan, Y.; Shabala, S.; Ma, Y.; Xu, R.; Zhou, M. Using QTL mapping to investigate the relationships between abiotic stress tolerance (drought and salinity) and agronomic and physiological traits. BMC Genom. 2015, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gorham, J. Salt tolerance in the triticeae: K/Na discrimination in Aegilops species. J. Exp. Bot. 1990, 41, 615–621. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Gupta, N.K.; Miyazaki, J.; Baho, M.N.; Chalmers, K.J.; Tester, M.; Langridge, P.; Collins, N.C. HvNax3—A locus controlling shoot sodium exclusion derived from wild barley (Hordeum vulgare ssp. spontaneum). Funct. Integr. Genom. 2010, 10, 277–291. [Google Scholar] [CrossRef]
- Forster, B.P. Mutation genetics of salt tolerance in barley: An assessment of Golden Promise and other semi-dwarf mutants. Euphytica 2001, 120, 317–328. [Google Scholar] [CrossRef]
- Forster, B.P.; Phillips, M.S.; Miller, T.E.; Baird, E.; Powell, W. Chromosome location of genes controlling tolerance to salt (NaCl) and vigour in hordeum vulgare and H. chilense. Heredity 1990, 65, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Von Korff, M.; Wang, H.; Léon, J.; Pillen, K. AB-QTL analysis in spring barley: II. Detection of favourable exotic alleles for agronomic traits introgressed from wild barley (H. vulgare ssp. spontaneum). Theor. Appl. Genet. 2006, 112, 1221–1231. [Google Scholar] [CrossRef] [Green Version]
- Laurie, D.A.; Pratchett, N.; Bezant, J.H.; Snape, J.W. RFLP mapping of five major genes and eight quantitative trait loci controlling flowering time in a winter X spring barley (Hordeum vulgare L.) cross. Genome 1995, 38, 575–585. [Google Scholar] [CrossRef]
- Zhou, G.; Johnson, P.; Ryan, P.R.; Delhaize, E.; Zhou, M. Quantitative trait loci for salinity tolerance in barley (Hordeum vulgare L.). Mol. Breed. 2012, 29, 427–436. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| 2017–2018 | 2018–2019 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Atmax | Atmin | FP | RH% | Prec. | Atmax | Atmin | FP | RH% | Train | |
| November | 22.9 | 12.6 | 8.9 | 61.2 | 27.4 | 23.9 | 14.1 | 9.8 | 58.6 | 6.4 |
| December | 20.7 | 11.3 | 6.4 | 57.8 | 0.8 | 18.5 | 9.6 | 7.0 | 65.3 | 8.5 |
| January | 17.5 | 7.9 | 5.8 | 66.1 | 14.8 | 17.4 | 7.1 | 1.8 | 52.6 | 0.7 |
| February | 21.6 | 11.2 | 5.3 | 52.1 | 26.7 | 18.8 | 8.2 | 3.6 | 54.8 | 7.4 |
| March | 26.3 | 12.5 | 5.2 | 42.5 | 0.4 | 21.1 | 9.2 | 4.8 | 52.9 | 8.2 |
| April | 27.5 | 14.1 | 7.9 | 46.4 | 50.5 | 25.3 | 12.3 | 5.8 | 44.5 | 1.1 |
| Characteristics | Soil | Well 1 | Well 2 |
|---|---|---|---|
| Soil particles distribution | |||
| Sand (%) | 81.53 | ||
| Silt (%) | 9.77 | ||
| Clay (%) | 8.70 | ||
| Textural class | Sandy loam | ||
| Calcium carbonate (CaCO3, %) | 57.99 | ||
| pH | 7.8 | 7.96 | 7.66 |
| Electrical conductivity (dS/m) | 8.82 | 9.35 (6500 ppm) | 13.50 (9500 ppm) |
| Saturation soluble extract cations and anions (mg/100 g) | |||
| Calcium (Ca++) | 25.2 | 23.5 | 19.3 |
| Magnesium (Mg++) | 5.7 | 14.5 | 18.8 |
| Sodium (Na+) | 57.8 | 66.1 | 105.1 |
| Carbonate (CO3=) | 0.0 | 0.0 | 0.0 |
| Bicarbonate (HCO3-) | 6.2 | 6.50 | 7.5 |
| Chloride (Cl–) | 61.9 | 57.66 | 93.1 |
| Sulphate (SO4=) | 26.4 | 29.23 | 38.7 |
| Source | DF | HD | PH | NSPLM | GWPLM | BWPLM | HI | TGW |
|---|---|---|---|---|---|---|---|---|
| Year (Y) | 1 | 280.56 ** | 10,030 ** | 59,374 ** | 453,690 ** | 4,171,113 ** | 33.81 * | 251.19 ** |
| Y (Rep) | 4 | 1.33 | 17.9 | 245.8 | 2121.2 | 13,568.8 | 4.32 | 9.61 |
| Salinity levels (SL) | 1 | 50,035 ** | 108,881 ** | 4,812,213 ** | 33,022,079 ** | 217,923,213 ** | 11,749 ** | 50,535 ** |
| Y × SL | 1 | 101.67 | 66.8 * | 1202.5 * | 47,309 ** | 183,453 ** | 68.06 ** | 20.51 ** |
| Y × SL (Rep) | 4 | 244.32 | 5.6 | 123.4 | 512.7 | 2106.9 | 1.78 | 0.83 |
| Genotypes (G) | 302 | 405.50 ** | 586.6 ** | 29,951 ** | 68,532 ** | 580,988 ** | 21.16 ** | 220.7 ** |
| G × Y | 302 | 4.63 ** | 16.7 ** | 1029.7 ** | 1473.8 ** | 15,905 ** | 7.38 ** | 0.04 |
| G × SL | 302 | 9.80 ** | 209.6 ** | 2766.8 ** | 13,605 ** | 98,932 ** | 4.03 | 10.33 ** |
| G × Y × SL | 302 | 1.00 | 17.1 ** | 497.2 ** | 750.2 ** | 6236 ** | 4.87 | 0.26 |
| Error | 2416 | 1.18 | 2.15 | 48.98 | 61.93 | 2807.4 | 3.99 | 0.235 |
| GCV% | 7.97 | 12.47 | 21.31 | 31.92 | 32.23 | 3.45 | 11.85 | |
| PCV% | 8.23 | 15.24 | 23.11 | 35.75 | 36.23 | 7.05 | 12.19 | |
| Heritability (H2b) | 93.97 | 66.99 | 85.06 | 79.72 | 79.15 | 23.98 | 94.51 |
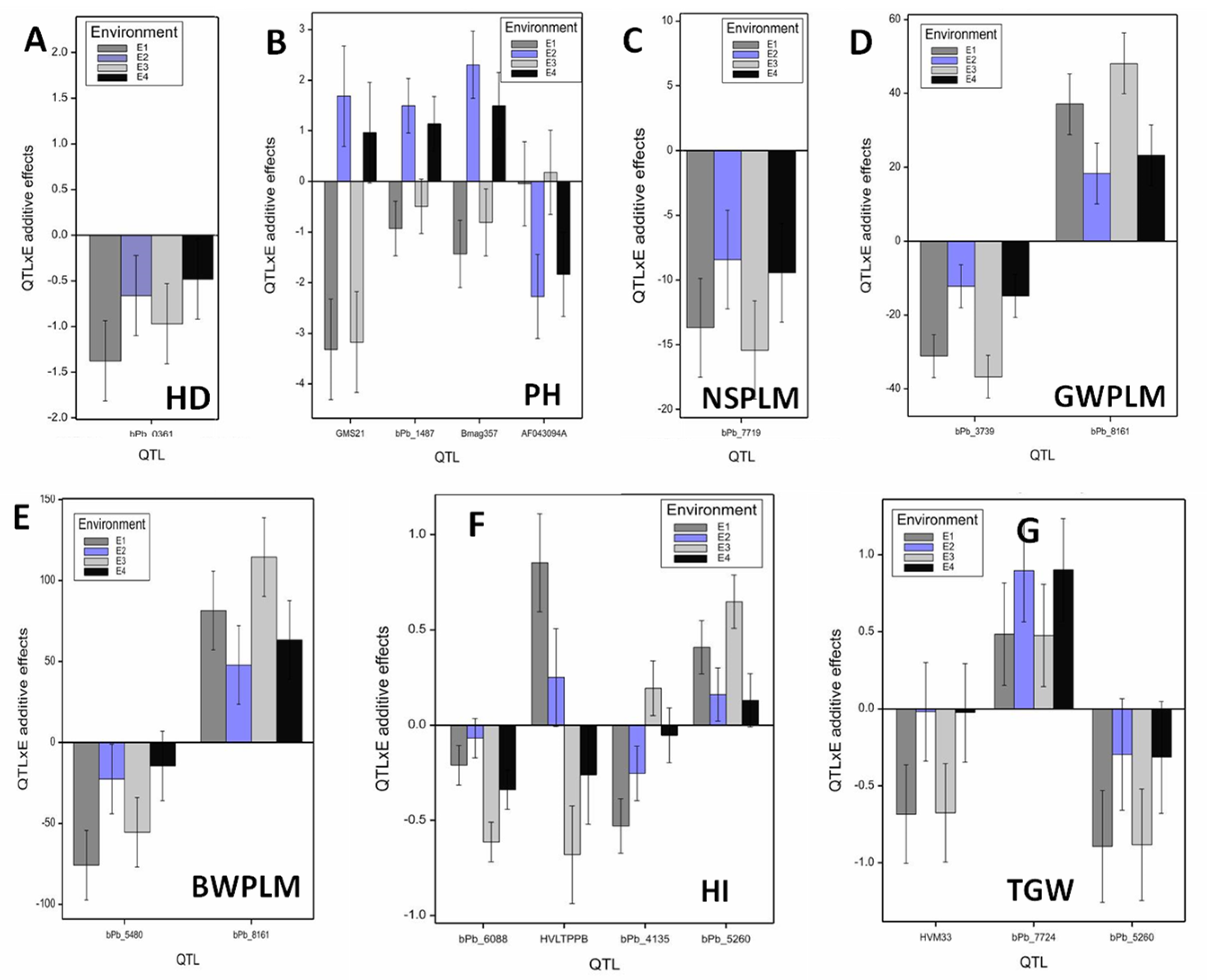
| Trait | (1) QTL | (2) Ch | (3) Marker | (4) Posi (cM) | (5) Effect | (6) F-Stat. | (7) LOD | (8) R2 | (9) Addi | (10) Allele | (11) S.E. | (12) QTL by E Additive Effects and S.E. | (15) S.E.-D | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (13) E1 | E2 | E3 | E4 | |||||||||||||||||
| (14) A1 | R2 | A2 | R2 | A3 | R2 | A4 | R2 | |||||||||||||
| HD | qHD.3H | 3H | bPb-0361 | 165.5 | M × E | 15.1 ** | 11.6 | 2.13 | −0.79 | ISR 42–8 | 0.45 | −1.32 | 4.90 | −0.57 | 1.00 | −0.91 | 2.20 | −0.38 | 0.40 | 0.13 |
| qHD.4H.a | 4H | HVPAZXG | 44.0 | M | 10.45 ** | 2.9 | 5.20 | −1.36 | Scarlett | 0.42 | Only main effect | |||||||||
| qHD.4H.b | 4H | bPb-6640 | 60.5 | M | 13.13 ** | 3.5 | 5.41 | −1.35 | Scarlett | 0.37 | Only main effect | |||||||||
| qHD.7H | 7H | bPb-5260 | 115.6 | M | 14.92 ** | 4.0 | 9.49 | −1.79 | Scarlett | 0.46 | Only main effect | |||||||||
| PH | qPH.1H.a | 1H | GMS21 | 14 | M × E | 21.72 ** | 12.9 | 9.48 | 0.96 | Scarlett | 1.00 | −3.32 | 17.00 | 1.69 | 3.60 | −3.17 | 16.00 | 0.96 | 1.30 | 0.81 |
| qPH.1H.b | 1H | bPb-1487 | 147.2 | M × E | 15.03 ** | 8.5 | 1.60 | 0.30 | ISR 42–8 | 0.54 | −0.93 | 1.30 | 1.49 | 2.80 | −0.49 | 0.40 | 1.14 | 1.90 | 0.44 | |
| qPH.3H | 3H | HV13GEIII | 150 | M | 40.98 ** | 9.2 | 22.51 | 3.31 | ISR 42–8 | 0.52 | Only main effect | |||||||||
| qPH.5H.a | 5H | Bmag357 | 68 | M × E | 22.42 ** | 13.2 | 3.55 | 0.39 | ISR 42–8 | 0.66 | −1.43 | 3.20 | 2.31 | 6.80 | −0.81 | 1.00 | 1.50 | 3.20 | 0.54 | |
| qPH.5H.b | 5H | AF043094A | 156 | M × E | 6.83 ** | 3.8 | 2.91 | 0.99 | Scarlett | 0.83 | −0.05 | 0.02 | −2.27 | 6.60 | 0.18 | 0.10 | −1.84 | 4.90 | 0.67 | |
| qPH.7H | 7H | bPb-1793 | 137.2 | M | 9.26 ** | 2.6 | 6.13 | 1.73 | Scarlett | 0.567 | Only main effect | |||||||||
| qNSPM.3H | 3H | HVM33 | 83 | M | 5.91 * | 1.8 | 1.48 | 9.031 | ISR 42–8 | 3.71 | Only main effect | |||||||||
| NSPLM | qNSPM.4H.a | 4H | MGB396 | 95 | M | 13.14 ** | 3.5 | 6.66 | 12.92 | Scarlett | 3.56 | Only main effect | ||||||||
| qNSPM.4H.b | 4H | bPb-7719 | 96.78 | M × E | 5.32 ** | 3.6 | 3.40 | 9.78 | Scarlett | 3.86 | 11.72 | 4.10 | 6.46 | 1.70 | 13.46 | 5.30 | 7.48 | 2.50 | 2.19 | |
| GWPLM | qGWPLM.4H | 4H | bPb-3739 | 96.31 | M, M × E | 17.79 ** | 13.9 | 8.43 | 23.71 | Scarlett | 5.80 | −31.10 | 9.00 | 12.20 | 6.10 | 36.72 | 11.30 | 14.81 | 7.30 | 4.15 |
| qGWPLM.7H | 7H | bPb-8161 | 2.22 | M, M × E | 12.45 ** | 9.4 | 15.98 | 31.70 | ISR 42–8 | 8.24 | 37.13 | 12.80 | 18.32 | 13.80 | 48.11 | 19.40 | 23.24 | 17.90 | 5.90 | |
| BWPLM | qBWPLM.4H.a | 4H | bPb-5480 | 72.2 | M × E | 6.9 ** | 4.8 | 3.20 | 42.15 | Scarlett | 21.49 | −75.91 | 6.70 | −22.52 | 2.20 | −55.47 | 3.20 | −14.68 | 0.70 | 14.63 |
| qBWPLM.4H.b | 4H | bPb-3739 | 96.3 | M | 19.98 ** | 5.0 | 9.55 | 68.17 | Scarlett | 15.25 | Only main effect | |||||||||
| qBWPLM.7H | 7H | bPb-8161 | 2.2 | M, M × E | 7.51 ** | 5.3 | 11.10 | 76.74 | ISR 42–8 | 24.31 | 81.44 | 7.70 | 47.77 | 9.80 | 114.45 | 13.60 | 63.28 | 13.30 | 16.55 | |
| HI | qHI.2H | 2H | bPb-6088 | 81.7 | M, M × E | 10.48 ** | 7.8 | 4.80 | 0.31 | Scarlett | 0.10 | −0.21 | 1.40 | −0.07 | 0.30 | −0.61 | 9.20 | −0.34 | 8.30 | 0.11 |
| qHI.3H | 3H | HVLTPPB | 25 | M × E | 9.08 ** | 6.6 | 10.68 | 0.04 | ISR 42–8 | 0.26 | 0.85 | 22.10 | 0.25 | 4.30 | −0.68 | 11.30 | −0.26 | 5.00 | 0.27 | |
| qHI.5H | 5H | bPb-4135 | 43.5 | M × E | 6.78 ** | 4.7 | 3.53 | 0.16 | Scarlett | 0.14 | −0.53 | 8.60 | −0.25 | 4.40 | 0.19 | 0.90 | −0.05 | 0.20 | 0.15 | |
| qHI.7H | 7H | bPb-5260 | 115.6 | M, M × E | 6.55 ** | 4.5 | 4.58 | 0.34 | ISR 42–8 | 0.14 | 0.41 | 5.10 | 0.16 | 1.70 | 0.65 | 10.30 | 0.13 | 1.20 | 0.15 | |
| TGW | qTGW.3H.a | 3H | HVM33 | 83 | M × E | 19.44 ** | 15.3 | 1.16 | 0.35 | Scarlett | 0.32 | −0.68 | 2.30 | −0.02 | 0.01 | −0.68 | 2.30 | −0.03 | 0.01 | 0.11 |
| qTGW.3H.b | 3H | bPb-7724 | 179.5 | M × E | 8.30 ** | 6.0 | 2.83 | 0.69 | ISR 42–8 | 0.33 | 0.48 | 1.20 | 0.90 | 4.40 | 0.48 | 1.20 | 0.90 | 4.50 | 0.11 | |
| qTGW.7H | 7H | bPb-5260 | 115.6 | M × E | 12.34 ** | 9.3 | 2.25 | 0.60 | Scarlett | 0.36 | −0.89 | 3.90 | −0.30 | 0.50 | −0.88 | 4.00 | −0.32 | 0.60 | 0.12 | |
| Trait | (1) QTL | (2) Chr. | (3) Marker | (4) Posi (cM) | (5) Effect | (6) F-Stat. | (7) LOD | (8) R2 | (9) Additive | (10) Allele | (11) S.E. | (12) QTL by E Additive Effects and S.E. | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (13) Y1 | R2 | Y2 | R2 | (14) SE. D | ||||||||||||
| HD | qSTI.HD.3H | 3H | bPb-0789 | 148.8 | M × E | 19.0 ** | 7.1 | 8.5 | 0.05 | Scarlett | 0.013 | −0.053 | 10.6 | −0.041 | 6.3 | 0.003 |
| qSTI.HD.4H | 4H | HVPAZXG | 44 | M × E | 13.9 ** | 4.2 | 2.7 | 0.03 | ISR 42-8 | 0.013 | 0.022 | 1.8 | 0.031 | 3.5 | 0.002 | |
| PH | qSTI.PH.3H | 3H | HV13GEIII | 150 | M | 15.2 ** | 4.0 | 8.7 | 0.07 | ISR 42-8 | 0.019 | Only main effect | ||||
| qSTI.PH.6H | 6H | bPb-4125 | 84.8 | M | 13.0 ** | 3.5 | 5.5 | 0.06 | ISR 42-8 | 0.016 | Only main effect | |||||
| NSPLMM | qSTI.NSPLM.1H | 1H | S53707 | 18 | M × E | 7.9 ** | 3.4 | 9.1 | 0.06 | ISR 42-8 | 0.054 | 0.173 | 12.4 | 0.108 | 5.8 | 0.021 |
| qSTI.NSPLM.4H | 4H | bPb-4216 | 69.66 | M | 13.8 ** | 3.7 | 10.5 | −0.15 | Scarlett | 0.041 | Only main effect | |||||
| qSTI.NSPLM.7H | 7H | bPb-9914 | 103.37 | M | 10.2 ** | 2.9 | 7.6 | −0.13 | Scarlett | 0.040 | Only main effect | |||||
| GWPLM | qSTI.GWPLM.1H.a | 1H | bPb-0405 | 7.21 | M × E | 9.2 ** | 6.8 | 17.8 | 0.33 | ISR 42-8 | 0.093 | 0.381 | 20.6 | 0.288 | 14.9 | 0.022 |
| qSTI.GWPLM.1H.b | 1H | bPb-1604 | 55.77 | M | 12.0 ** | 2.6 | 8.7 | 0.23 | ISR 42-8 | 0.077 | Only main effect | |||||
| qSTI.GWPLM.4H | 4H | bPb-3512 | 67.92 | M | 7.0 ** | 3.3 | 4.9 | −0.17 | Scarlett | 0.050 | Only main effect | |||||
| qSTI.GWPLM.7H | 7H | bPb-6706 | 58.17 | M | 18.3 ** | 2.1 | 14.4 | 0.30 | Scarlett | 0.113 | Only main effect | |||||
| BWPLM | qSTI.BWPLM.1H | 1H | bPb-0405 | 7.21 | M × E | 10.7 ** | 4.7 | 16.0 | 0.31 | ISR 42-8 | 0.090 | 0.352 | 18.5 | 0.269 | 13.5 | 0.027 |
| qSTI.BWPLM.4H | 4H | bPb-3512 | 67.92 | M | 11.3 ** | 3.1 | 4.8 | −0.17 | Scarlett | 0.050 | Only main effect | |||||
| qSTI.BWPLM.7H.a | 7H | bPb-6706 | 58.17 | M × E | 7.2 ** | 3.1 | 15.4 | 0.30 | ISR 42-8 | 0.113 | 0.349 | 18.2 | 0.259 | 12.5 | 0.034 | |
| qSTI.BWPLM.7H.b | 7H | bPb-5260 | 115.56 | M | 9.5 ** | 2.7 | 5.6 | −0.18 | Scarlett | 0.059 | Only main effect | |||||
| HI | qSTI.HI.1H.a | 1H | bPb-0405 | 7.21 | M | 15.7 | 4.1 | 14.9 | 0.04 | ISR 42-8 | 0.009 | Only main effect | ||||
| qSTI.HI.1H.b | 1H | bPb-2240 | 123.09 | M | 18.0 ** | 4.6 | 4.9 | −0.02 | Scarlett | 0.005 | Only main effect | |||||
| qSTI.HI.2H | 2H | Bmag381 | 97 | M | 13.4 ** | 3.6 | 4.0 | 0.02 | ISR 42-8 | 0.005 | Only main effect | |||||
| qSTI.HI.4H | 4H | Mlo | 127.5 | M | 17.6 ** | 4.6 | 5.8 | 0.02 | ISR 42-8 | 0.006 | Only main effect | |||||
| qSTI.HI.7H | 7H | HVSS1 | 49 | M × E | 7.3 ** | 3.2 | 20.4 | 0.04 | ISR 42-8 | 0.017 | 0.018 | 4.0 | 0.064 | 36.8 | 0.017 | |
| TGW | qSTI.TGW.2H | 2H | HVM36 | 26.5 | M | 12.4 ** | 3.4 | 7.2 | −0.07 | Scarlett | 0.020 | Only main effect | ||||
| qSTI.TGW.3H | 3H | bPb-4472 | 1.48 | M × E | 5.0 * | 2.2 | 3.3 | −0.05 | Scarlett | 0.032 | −0.049 | 3.4 | −0.046 | 3.1 | 0.001 | |
| qSTI.TGW.4H.a | 4H | GMS89 | 57 | M × E | 14.4 ** | 6.2 | 7.8 | −0.07 | Scarlett | 0.017 | −0.074 | 7.9 | −0.073 | 7.7 | 0.001 | |
| qSTI.TGW.4H.b | 4H | Mlo | 127.5 | M × E | 9.1 ** | 4.0 | 3.0 | 0.05 | ISR 42-8 | 0.018 | 0.046 | 3.1 | 0.045 | 2.9 | 0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, M.A.; Nassar, S.M.; Moustafa, E.S.; Said, M.T.; Börner, A.; Hamada, A. Genetic Mapping Reveals Novel Exotic and Elite QTL Alleles for Salinity Tolerance in Barley. Agronomy 2021, 11, 1774. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091774
Sayed MA, Nassar SM, Moustafa ES, Said MT, Börner A, Hamada A. Genetic Mapping Reveals Novel Exotic and Elite QTL Alleles for Salinity Tolerance in Barley. Agronomy. 2021; 11(9):1774. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091774
Chicago/Turabian StyleSayed, Mohammed Abdelaziz, Saad Mohamed Nassar, Ehab Soudi Moustafa, Mohamed Tharwat Said, Andreas Börner, and Alhosein Hamada. 2021. "Genetic Mapping Reveals Novel Exotic and Elite QTL Alleles for Salinity Tolerance in Barley" Agronomy 11, no. 9: 1774. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091774