
The Effect of Night Low Temperature on Agronomical Traits of Thirty-Nine Pepper Accessions (Capsicum annuum L.)

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Soil Preparations
2.3. Temperature Regulations
2.4. Diseases and Pest Controls
2.5. Data Collections
2.6. Data Analysis
3. Results
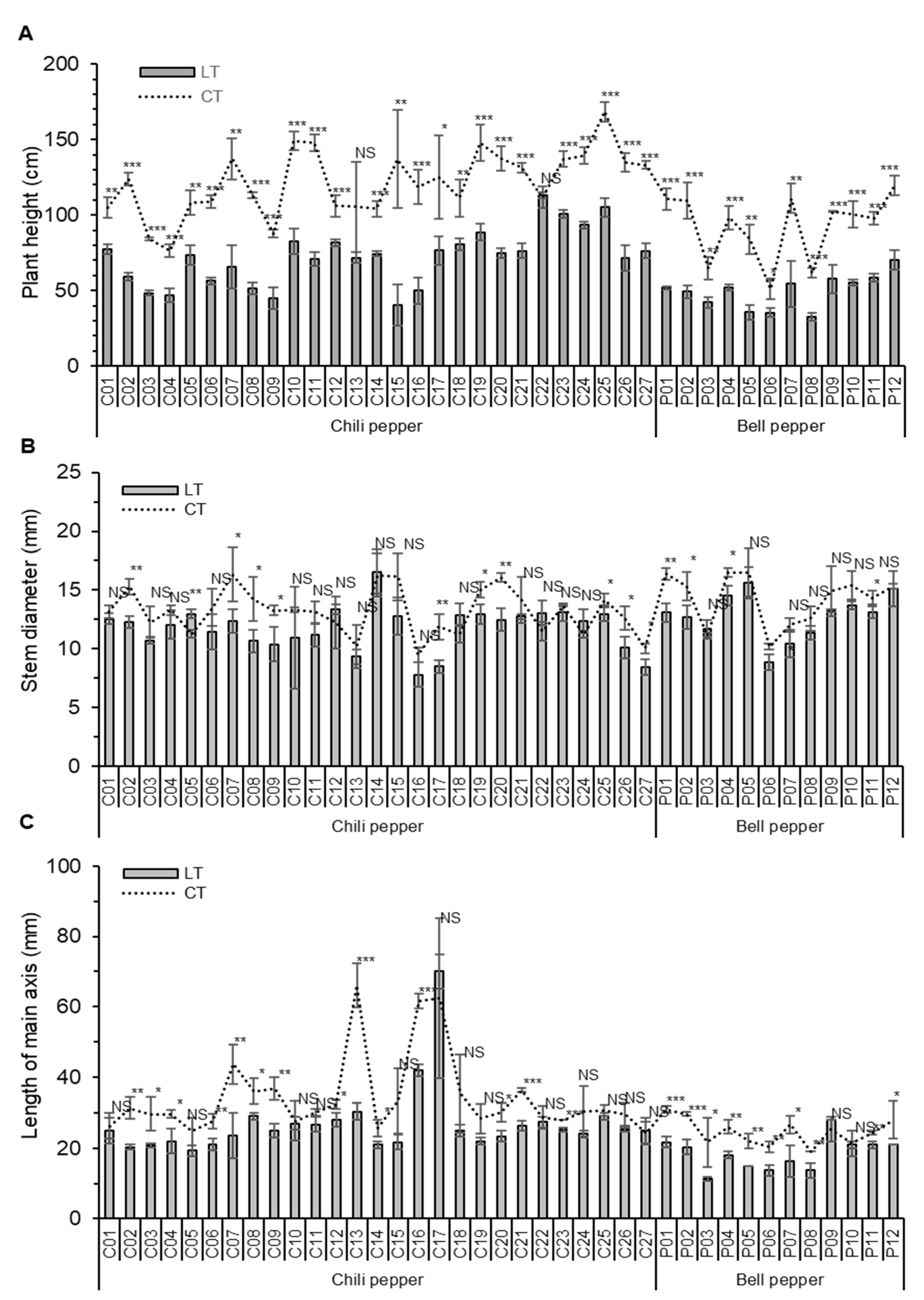
3.1. The Vegetative Traits with Chili and Bell Peppers
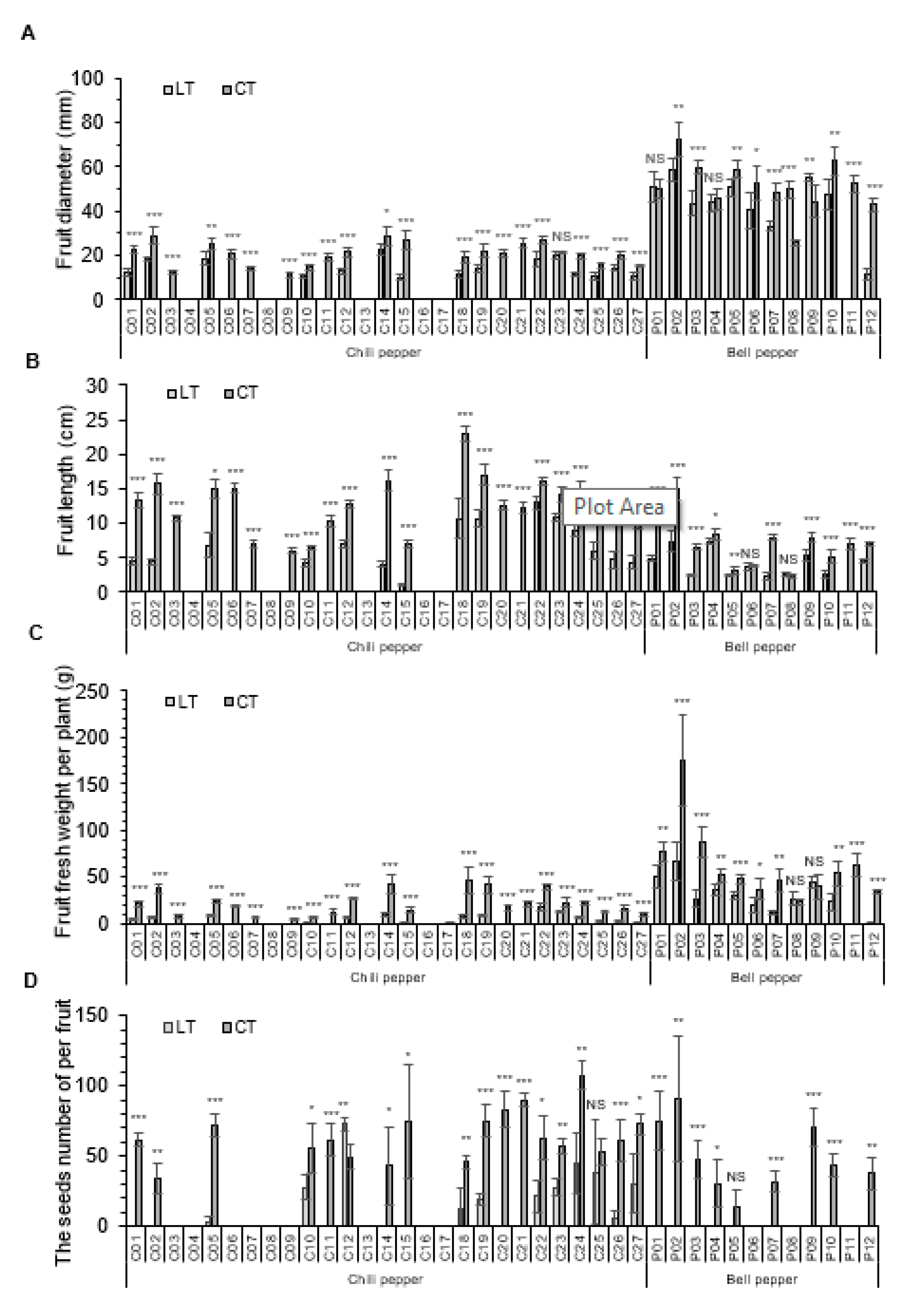
3.2. The Reproductive Traits of NFL, NFR, and FY with Chili and Bell Peppers
3.3. The Fruit Traits of FD, FL, FFW, and NSF with Chili and Bell Peppers
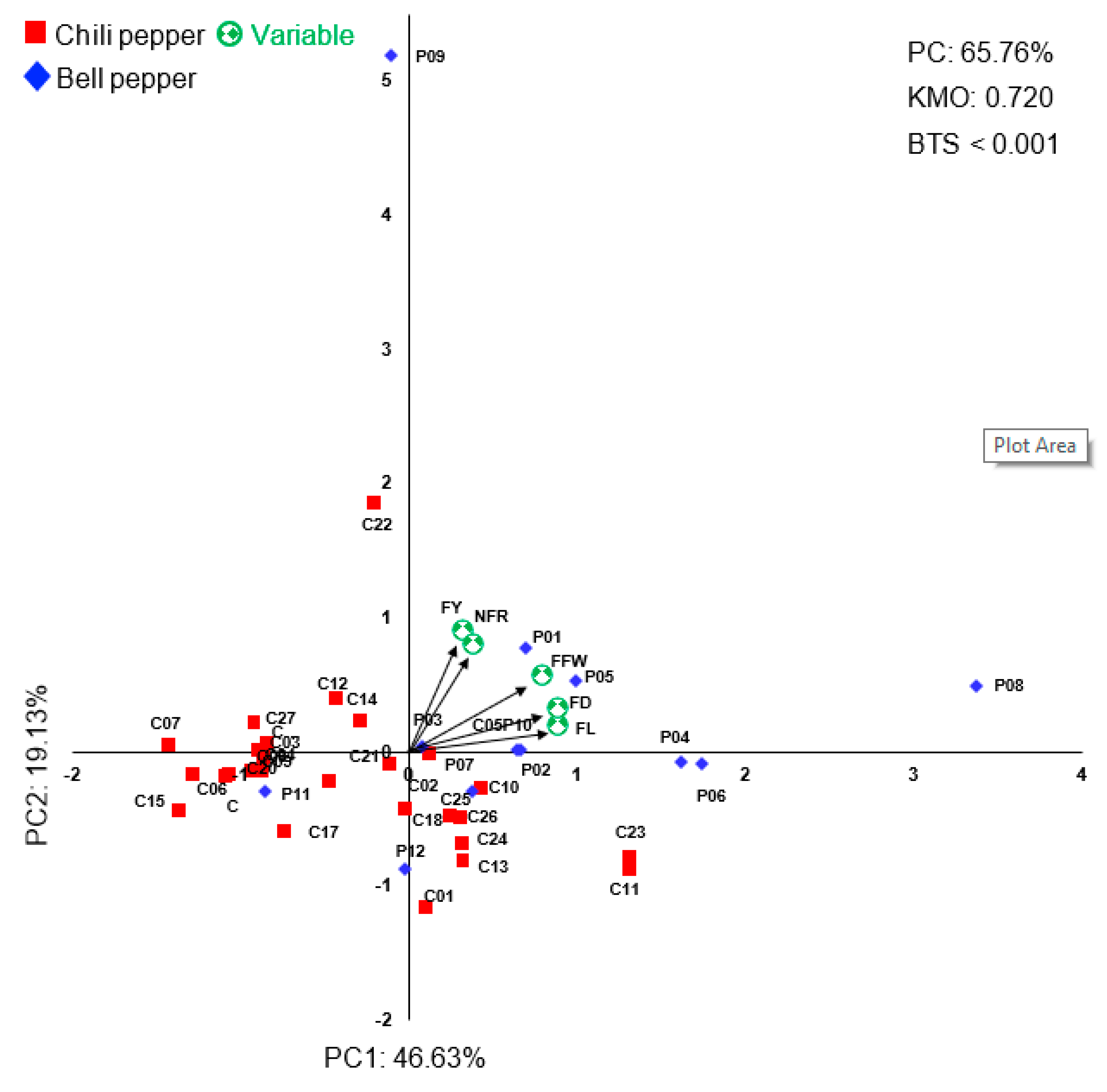
3.4. The Principal Component Analysis (PCA) of Agronomical Traits
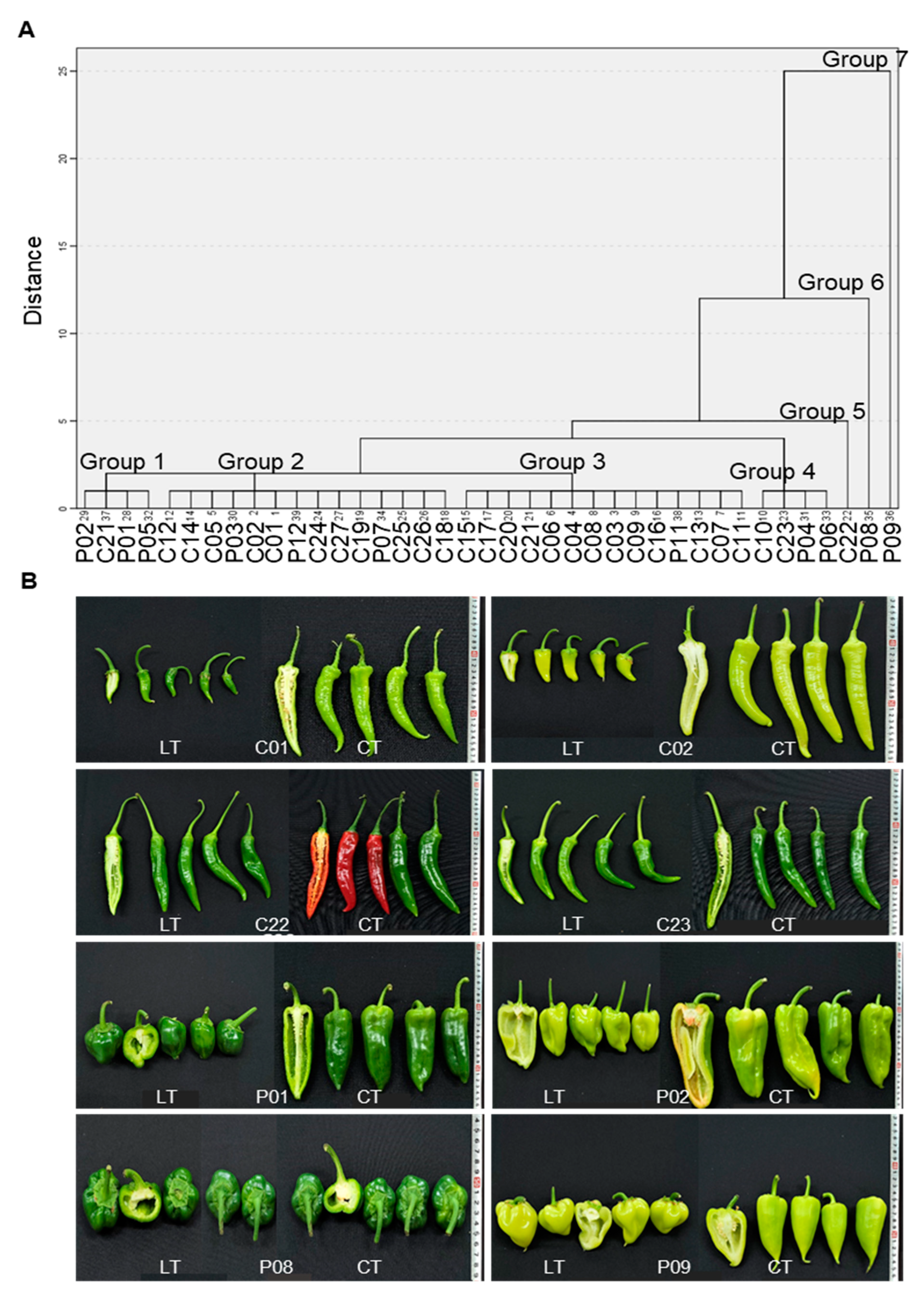
3.5. Clustering Analysis
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kothari, S.; Joshi, A.; Kachhwaha, S.; Ochoa-Alejo, N. Chilli peppers—A review on tissue culture and transgenesis. Biotechnol. Adv. 2010, 28, 35–48. [Google Scholar] [CrossRef]
- Bhutia, K.; Khanna, V.; Meetei, T.; Bhutia, N. Effects of climate change on growth and development of chilli. Agrotechnology 2018, 7, 2. [Google Scholar] [CrossRef]
- Prohens, J.; Nuez, F. Handbook of Plant Breeding. Vegetables II: Fabaceae, Liliaceae, Solanaceae and Umbelliferae; Springer: New York, NY, USA, 2008; Volume 3, pp. 30–40. [Google Scholar]
- Sun, T.; Xu, Z.; Wu, C.T.; Janes, M.; Prinyawiwatkul, W.; No, H. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L.). J. Food Sci. 2007, 72, 98–102. [Google Scholar] [CrossRef]
- Sarada, C.; Ratnam, M.; Naidu, L.; Ramana, C.; Rajani, A.; Vijaya, T. Chilli production and productivity in relation to Seasonal weather conditions in Guntur District of Andhra Pradesh. Int. J. Pure Appl. Biosci. 2015, 3, 207–213. [Google Scholar]
- Cruz-Huerta, N.; Williamson, J.G.; Darnell, R.L. Low night temperature increases ovary size in sweet pepper cultivars. HortScience 2011, 46, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Toki, T.; Ogiwara, S.; Aoki, H. Effect of varying night temperature on the growth and yields in cucumber. Acta Hortic. 1978, 87, 233–238. [Google Scholar] [CrossRef]
- Horie, T.; de Wit, C.T.; Goudriaan, J.; Bensink, J. A formal template for the development of cucumber in its vegetative stage (I, II and III). In Proceedings of the Koninklijke Nederlandse Akademie Van Wetenschappen. Serie C: Biological and Medical Sciences; Wageningen University: Wageningen, The Netherlands, 1979; Volume 82, pp. 433–479. [Google Scholar]
- Nilwik, H. Growth analysis of sweet pepper (Capsicum annuum L.) 1. The influence of irradiance and temperature under glasshouse conditions in winter. Ann. Bot. 1981, 48, 129–136. [Google Scholar] [CrossRef]
- Ji, L.; Li, P.; Su, Z.; Li, M.; Guo, S. Cold-tolerant introgression line construction and low-temperature stress response analysis for bell pepper. Plant Signal. Behav. 2020, 15, 1773097. [Google Scholar] [CrossRef] [PubMed]
- Foolad, M.; Lin, G. Relationship between cold tolerance during seed germination and vegetative growth in tomato: Germplasm evaluation. J. Am. Soc. Hortic. Sci. 2000, 125, 679–683. [Google Scholar] [CrossRef]
- O’SULLIVAN, J.; Bouw, W. Pepper seed treatment for low-temperature germination. Can. J. Plant Sci. 1984, 64, 387–393. [Google Scholar] [CrossRef]
- Seo, J.-U.; Hwang, J.-M.; Oh, S.-M. Effects of night temperature treatment of raising seedlings before transplanting on growth and development of pepper. J. Bio-Env. Con. 2006, 15, 149–155. [Google Scholar]
- Bhatt, R.; Srinivasa Rao, N. Response of bell-pepper (Capsicum annuum) photosynthesis, growth, and flower and fruit setting to night temperature. Photosynthetica 1994, 28, 127–132. [Google Scholar]
- Sherzod, R.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Kim, J.H.; Nam, C.W.; Chae, W.B. Traits affecting low temperature tolerance in tomato and its application to breeding program. Plant Breed. Biotechnol. 2019, 7, 350–359. [Google Scholar] [CrossRef]
- Rajametov, S.N.; Lee, K.; Jeong, H.-B.; Cho, M.-C.; Nam, C.-W.; Yang, E.-Y. Physiological Traits of Thirty-Five Tomato Accessions in Response to Low Temperature. Agriculture 2021, 11, 792. [Google Scholar] [CrossRef]
- Oh, S.-Y.; Koh, S.C. Fruit Development and Quality of Hot Pepper (Capsicum annuum L.) under Various Temperature Regimes. Hortic. Sci. Technol. 2019, 37, 313–321. [Google Scholar]
- Mercado, J.; Mar Trigo, M.; Reid, M.; Valpuesta, V.; Quesada, M. Effects of low temperature on pepper pollen morphology and fertility: Evidence of cold induced exine alterations. J. Hortic. Sci. 1997, 72, 317–326. [Google Scholar] [CrossRef]
- Pressman, E.; Moshkovitch, H.; Rosenfeld, K.; Shaked, R.; Gamliel, B.; Aloni, B. Influence of low night temperatures on sweet pepper flower quality and the effect of repeated pollinations, with viable pollen, on fruit setting. J. Hortic. Sci. Biotechnol. 1998, 73, 131–136. [Google Scholar] [CrossRef]
- Shaked, R.; Rosenfeld, K.; Pressman, E. The effect of low night temperatures on carbohydrates metabolism in developing pollen grains of pepper in relation to their number and functioning. Sci. Hortic. 2004, 102, 29–36. [Google Scholar] [CrossRef]
- Kato, K. Flowering and fertility of forced green peppers at lower temperatures. J. Jpn. Soc. Hortic. Sci. 1989, 58, 113–121. [Google Scholar] [CrossRef]
- Rylski, I. Effect of night temperature on shape and size of sweet pepper (Capsicum annuum L.). Amer. Soc. Hort. Sci. J. 1973, 98, 149–152. [Google Scholar]
- Aloni, B.; Pressman, E.; Karni, L. The effect of fruit load, defoliation and night temperature on the morphology of pepper flowers and on fruit shape. Ann. Bot. 1999, 83, 529–534. [Google Scholar] [CrossRef] [Green Version]
- De Koning, A. The effect of different day/night temperature regimes on growth, development and yield of glasshouse tomatoes. J. Hortic. Sci. 1988, 63, 465–471. [Google Scholar] [CrossRef]
- Rural Development Administration (RDA). Data Book of Agricultural Products Income for the Improvement of Agricultural Management in 2019; Rural Development Administration: Jeonju, Korea, 2020. [Google Scholar]
- Elings, A.; Kempkes, F.; Kaarsemaker, R.; Ruijs, M.; Van De Braak, N.; Dueck, T. The Energy Balance and Energy-Saving Measures in Greenhouse Tomato Cultivation. Acta Hortic. 2005, 691, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Rylski, I.; Spigelman, M. Effects of different diurnal temperature combinations on fruit set of sweet pepper. Sci. Hortic. 1982, 17, 101–106. [Google Scholar] [CrossRef]
- Rylski, I. Investigations on the Influence of Suboptimal Temperatures on the Flowering, Fruit Setting and Development of Sweet Pepper (Capsicum annum L.). Ph. D. Thesis, Hebrew University of Jerusalem, Jerusalem, Israel, 1971; pp. 1–96. [Google Scholar]
- Rylski, E.; Kempler, H. Fruit set of sweet pepper (Capsicum annuum L.) under plastic covers. HortScience 1972, 7, 422–423. [Google Scholar]
- Wang, C. Alleviation of chilling injury in tropical and subtropical fruits. In III International Symposium on Tropical and Subtropical Fruits; International Society for Horticultural Science: Bierbeek, Belgium, 2004; Volume 864, pp. 267–273. [Google Scholar]
- Barchi, L.; Lefebvre, V.; Sage-Palloix, A.-M.; Lanteri, S.; Palloix, A. QTL analysis of plant development and fruit traits in pepper and performance of selective phenotyping. Theor. Appl. Genet. 2009, 118, 1157–1171. [Google Scholar] [CrossRef] [PubMed]
- Yarnes, S.C.; Ashrafi, H.; Reyes-Chin-Wo, S.; Hill, T.A.; Stoffel, K.M.; Van Deynze, A. Identification of QTLs for capsaicinoids, fruit quality, and plant architecture-related traits in an interspecific Capsicum RIL population. Genome 2013, 56, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Tang, X.-F.; Ma, N.-N.; Wang, L.-Y.; Meng, Q.-W. Heterology expression of the sweet pepper CBF3 gene confers elevated tolerance to chilling stress in transgenic tobacco. J. Plant Physiol. 2011, 168, 1804–1812. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.-M.; Zhang, H.-F.; Liu, S.-Y.; Wang, X.-K.; Zhang, Y.-M.; Meng, Y.-C.; Luo, D.; Chen, R.-G. The NAC transcription factor CaNAC064 is a regulator of cold stress tolerance in peppers. Plant Sci. 2020, 291, 110346. [Google Scholar] [CrossRef]
- Kong, X.-M.; Zhou, Q.; Zhou, X.; Wei, B.-D.; Ji, S.-J. Transcription factor CaNAC1 regulates low-temperature-induced phospholipid degradation in green bell pepper. J. Exp. Bot. 2020, 71, 1078–1091. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.-K.; Sunkar, R. Gene Regulation During Cold Stress Acclimation in Plants. Methods Mol. Biol. 2010, 639, 39–55. [Google Scholar] [PubMed] [Green Version]
- Bakker, J.; Van Uffelen, J. The effects of diurnal temperature regimes on growth and yield of glasshouse sweet pepper. Neth. J. Agri. Sci. 1988, 36, 201–208. [Google Scholar]
- Ohtaka, K.; Yoshida, A.; Kakei, Y.; Fukui, K.; Kojima, M.; Takebayashi, Y.; Yano, K.; Imanishi, S.; Sakakibara, H. Difference Between Day and Night Temperatures Affects Stem Elongation in Tomato (Solanum lycopersicum) Seedlings via Regulation of Gibberellin and Auxin Synthesis. Front. Plant Sci. 2020, 11, 1947. [Google Scholar] [CrossRef] [PubMed]
- Phatak, S. Top and root temperature effects on tomato flowering. J. Am. Soc. Hortic. Sci. 1966, 88, 527–531. [Google Scholar]
- Goodstal, F.J.; Kohler, G.R.; Randall, L.B.; Bloom, A.J.; Clair, D.A.S. A major QTL introgressed from wild Lycopersicon hirsutum confers chilling tolerance to cultivated tomato (Lycopersicon esculentum). Theor. Appl. Genet. 2005, 111, 898–905. [Google Scholar] [CrossRef]
- Patterson, B.D.; Reid, M.S. Genetic and environmental influences on the expression of chilling injury. In Chilling Injury of Horticultural Crops; CRC Press: Boca Raton, FL, USA, 1990; pp. 87–112. [Google Scholar]
- Issarakraisila, M.; Considine, J. Effects of temperature on pollen viability in mango cv. ‘Kensington’. Ann. Bot. 1994, 73, 231–240. [Google Scholar] [CrossRef]
- Satake, T.; Hayase, H. Male sterility caused by cooling treatment at the young microspore stage in rice plants: V. Estimations of pollen developmental stage and the most sensitive stage to coolness. Jpn. J. Crop Sci. 1970, 39, 468–473. [Google Scholar] [CrossRef]
- Polowick, P.; Sawhney, V. Temperature effects on male fertility and flower and fruit development in Capsicum annuum L. Sci. Hortic. 1985, 25, 117–127. [Google Scholar] [CrossRef]
- Sawhney, V.K.; Shukla, A. Male sterility in flowering plants: Are plant growth substances involved? Am. J. Bot. 1994, 81, 1640–1647. [Google Scholar] [CrossRef]
- Zhigila, D.A.; AbdulRahaman, A.A.; Kolawole, O.S.; Oladele, F.A. Fruit morphology as taxonomic features in five varieties of Capsicum annuum L. Solanaceae. J. Bot. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Paran, I.; Van Der Knaap, E. Genetic and molecular regulation of fruit and plant domestication traits in tomato and pepper. J. Exp. Bot. 2007, 58, 3841–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilarinho, L.B.O.; da Silva, D.J.H.; Greene, A.; Salazar, K.D.; Alves, C.; Eveleth, M.; Nichols, B.; Tehseen, S.; Khoury, J.K.; Johnson, J.V. Inheritance of fruit traits in Capsicum annuum: Heirloom cultivars as sources of quality parameters relating to pericarp shape, color, thickness, and total soluble solids. J. Am. Soc. Hortic. Sci. 2015, 140, 597–604. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajametov, S.N.; Lee, K.; Jeong, H.-B.; Cho, M.-C.; Nam, C.-W.; Yang, E.-Y. The Effect of Night Low Temperature on Agronomical Traits of Thirty-Nine Pepper Accessions (Capsicum annuum L.). Agronomy 2021, 11, 1986. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11101986
Rajametov SN, Lee K, Jeong H-B, Cho M-C, Nam C-W, Yang E-Y. The Effect of Night Low Temperature on Agronomical Traits of Thirty-Nine Pepper Accessions (Capsicum annuum L.). Agronomy. 2021; 11(10):1986. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11101986
Chicago/Turabian StyleRajametov, Sherzod Nigmatullayevich, Kwanuk Lee, Hyo-Bong Jeong, Myeong-Cheoul Cho, Chun-Woo Nam, and Eun-Young Yang. 2021. "The Effect of Night Low Temperature on Agronomical Traits of Thirty-Nine Pepper Accessions (Capsicum annuum L.)" Agronomy 11, no. 10: 1986. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11101986