
The Role of Laccase from Zygomycetous Fungus Mortierella elasson in Humic Acids Degradation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain, Media and Culture Conditions
2.1.1. Inoculum Preparation
2.1.2. Induction of Laccase
2.2. Enzyme Purification
2.3. Enzyme Characterization
2.3.1. Enzyme Activity
2.3.2. Optimal pH and pH Stability
2.3.3. Temperature Optimum and Thermal Stability
2.3.4. Kinetic Constants of Substrate Oxidation
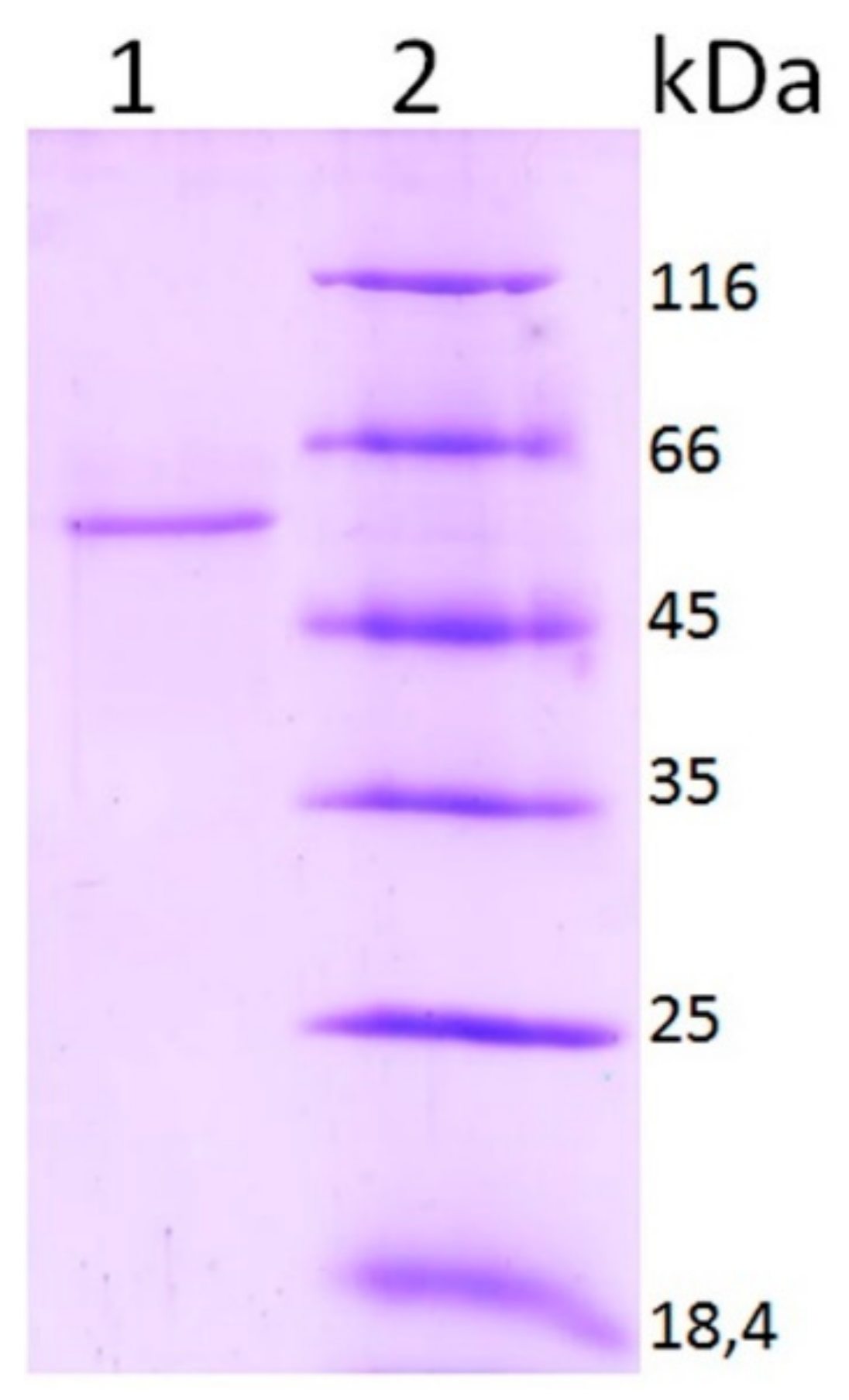
2.3.5. Molecular Weight Determination
2.4. Humic Acids Preparations
2.4.1. Extraction and Purification of HA
2.4.2. Characterization of the HA
2.5. Reaction of Laccase with Humic Acids
3. Results
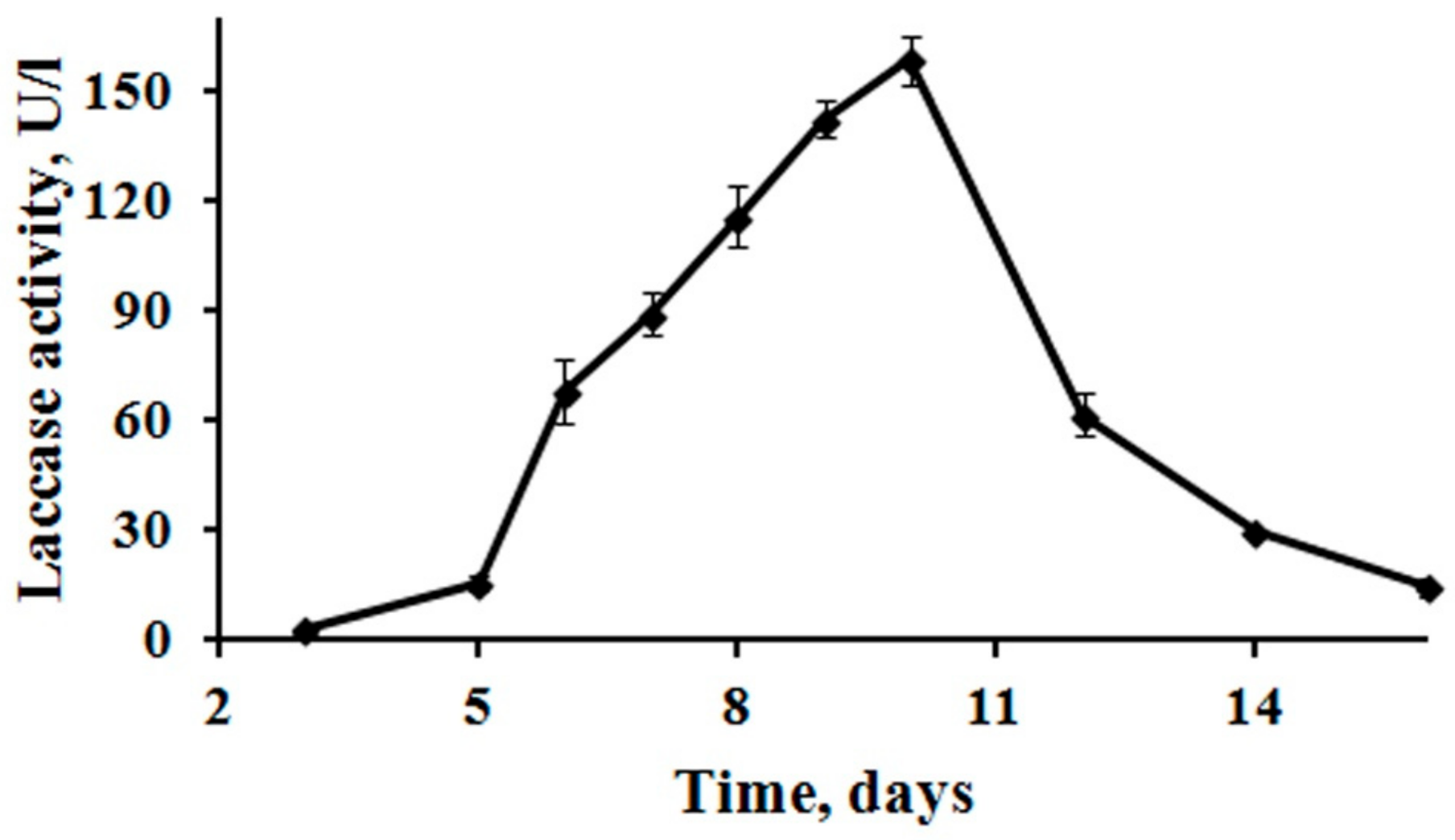
3.1. Production of Laccase
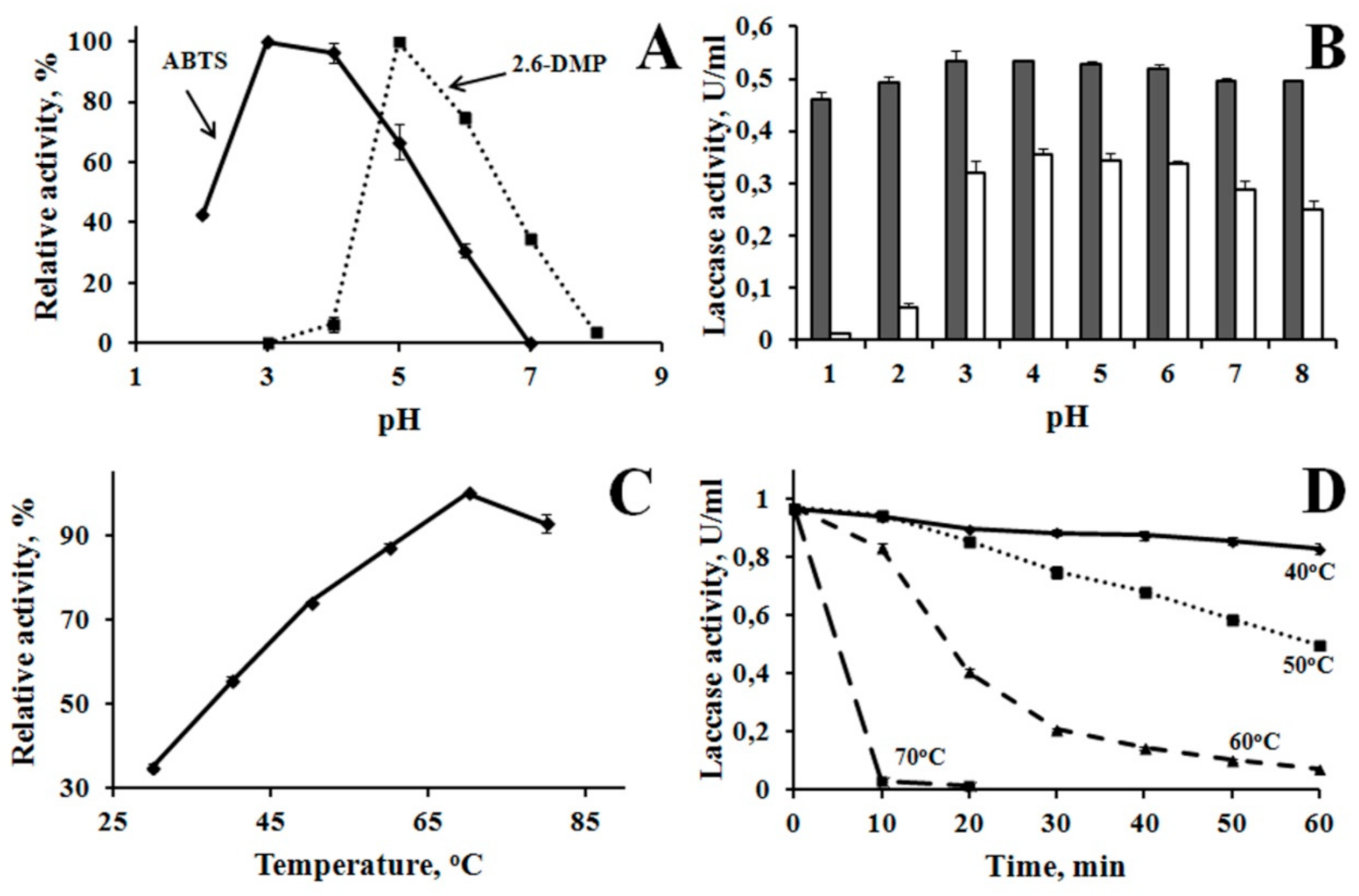
3.2. Properties of Laccase
3.3. Humic Acids
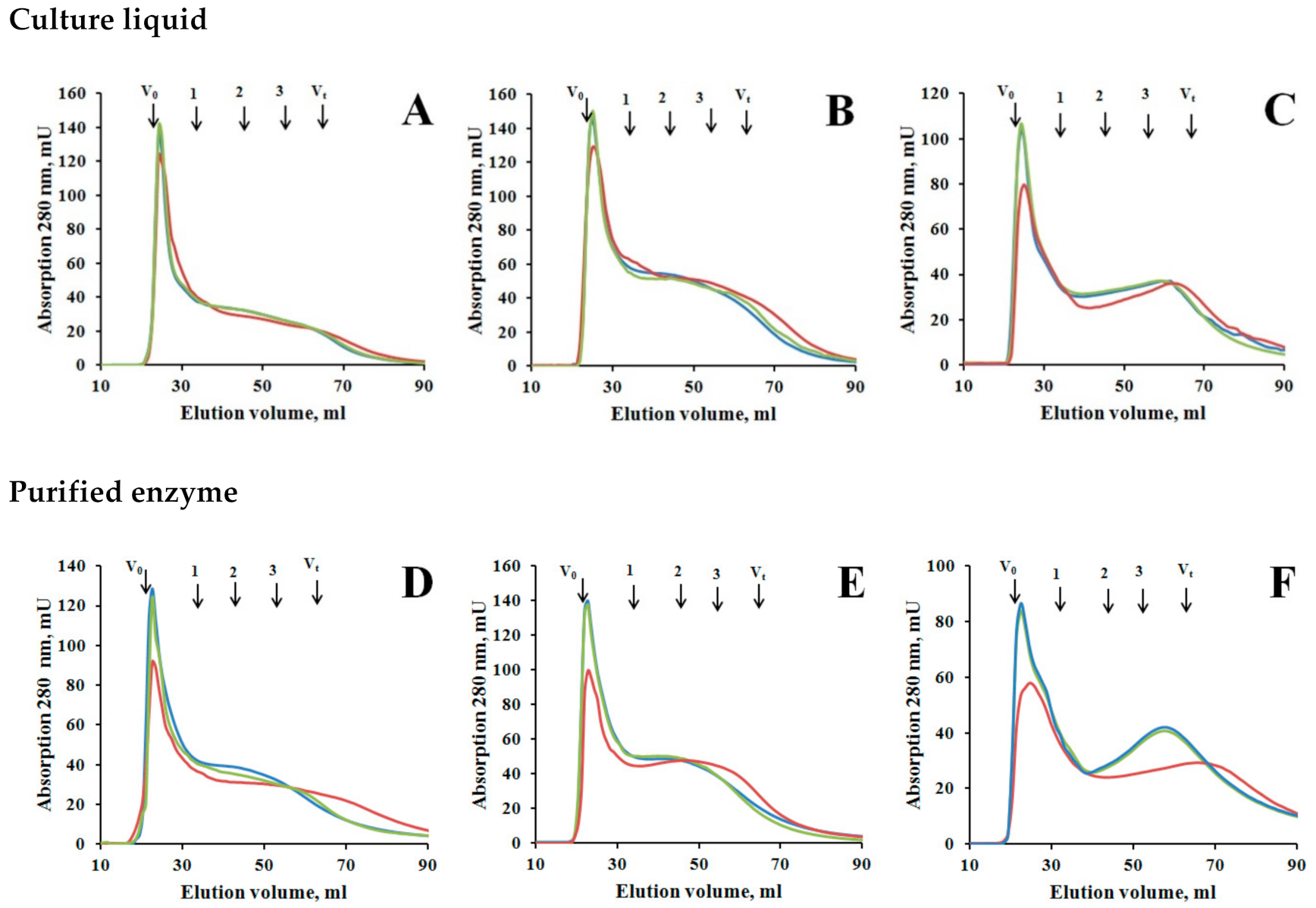
3.4. Interaction of Laccase with Humic Acids
4. Discussion
4.1. Production of Laccase by Mortierella Elasson
4.2. Properties of Laccase of M. elasson
4.3. Transformation of Humic Acids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Institutional Review Board Statement
Conflicts of Interest
References
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 2014, 65, 10–21. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kogel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenov, V.M.; Tulina, A.S.; Semenova, N.A.; Ivannikova, L.A. Humification and nonhumification pathways of the organic matter stabilization in soil: A review. Eurasian Soil Sci. 2013, 46, 355–368. [Google Scholar] [CrossRef]
- Stevenson, F.J. Humus Chemistry: Genesis, Composition, Reactions, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 1994; p. 512. [Google Scholar]
- Zavarzina, A.G.; Lisov, A.A.; Zavarzin, A.A.; Leontievsky, A.A. Fungal oxidoreductases and humification in forest soils. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 22, pp. 207–228. [Google Scholar] [CrossRef]
- Solomon, E.I.; Uma, M.S.; Machonkin, T.E. Multicopper oxidases and oxygenases. Chem. Rev. 1996, 96, 2563–2606. [Google Scholar] [CrossRef]
- Baldrian, P. Fungal laccases-occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef] [Green Version]
- Gorbacheva, M.; Morozova, O.; Shumakovich, G.; Streltsov, A.; Shleev, S.; Yaropolov, A. Enzymatic oxidation of manganese ions catalysed by laccase. Bioorg. Chem. 2009, 37, 1–5. [Google Scholar] [CrossRef]
- Grinhut, T.; Hadar, Y.; Chen, Y. Degradation and transformation of humic substances by saprotrophic fungi: Processes and mechanisms. Fungal Biol. Rev. 2007, 21, 179–189. [Google Scholar] [CrossRef]
- Zavarzina, A.G.; Leontievsky, A.A.; Golovleva, L.A.; Trofimov, S.Y. Biotransformation of soil humic acids by blue laccase of Panus tigrinus 8/18: An in vitro study. Soil Biol. Biochem. 2004, 36, 359–369. [Google Scholar] [CrossRef]
- Zavarzina, A.G.; Lisov, A.V.; Leontievsky, A.A. The role of ligninolytic enzymes laccase and a versatile peroxidase of the white-rot fungus Lentinus tigrinus in biotransformation of soil humic matter: Comparative in vivo study. J. Geophys. Res. Biogeosciences 2018, 123, 2727–2742. [Google Scholar] [CrossRef]
- Thurston, C.F. The structure and function of fungal laccases. Microbiology 1994, 140, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.M.; Harel, E. Polyphenol oxidases in plants. Phytochemistry 1979, 18, 193–215. [Google Scholar] [CrossRef]
- Dittmer, N.T.; Gorman, M.J.; Kanost, M.R. Characterization of endogenous and recombinant forms of laccase-2, a multicopper oxidase from the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 2009, 39, 596–606. [Google Scholar] [CrossRef] [Green Version]
- Otto, B.; Schlosser, D. First laccase in green algae: Purification and characterization of an extracellular phenol oxidase from Tetracystis aeria. Planta 2014, 240, 1225–1236. [Google Scholar] [CrossRef]
- Ece, S.; Lambertz, C.; Fischer, R.; Commandeur, U. Heterologous expression of a Streptomyces cyaneus laccase for biomass modification applications. AMB Express 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Machczynski, M.C.; Vijgenboom, E.; Samyn, B.; Canters, G.W. Characterization of SLAC: A small laccase from Streptomyces coelicolor with unprecedented activity. Protein Sci. 2004, 13, 2388–2397. [Google Scholar] [CrossRef] [Green Version]
- Arregui, L.; Ayala, M.; Gómez-Gil, X.; Gutiérrez-Soto, G.; Hernández-Luna, C.E.; de Los Santos, M.H.; Levin, L.; Rojo-Domínguez, A.; Romero-Martínez, D.; Saparrat, M.C.; et al. Laccases: Structure, function, and potential application in water bioremediation. Microb. Cell Fact. 2019, 18, 1–33. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.F.; Wheeler, M.H.; Chang, Y.C.; Kwon-Chung, K.J. A developmentally regulated gene cluster involved in conidial pigment biosynthesis in Aspergillus fumigatus. J. Bacteriol. 1999, 181, 6469–6477. [Google Scholar] [CrossRef] [Green Version]
- Frases, S.; Salazar, A.; Dadachova, E.; Casadevall, A. Cryptococcus neoformans can utilize the bacterial melanin precursor homogentisic acid for fungal melanogenesis. Appl. Environ. Microbiol. 2007, 73, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Hu, J.; Regner, R.; Round, J.W.; Ralph, J.; Saddler, J.N.; Eltis, L.D. Enhanced delignification of steam-pretreated poplar by a bacterial laccase. Sci. Rep. 2017, 7, 42121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaitan, I.J.; Medina, S.C.; González, J.C.; Rodríguez, A.; Espejo, A.J.; Osma, J.F.; Sarria, V.; Alméciga-Díaz, C.J.; Sánchez, O.F. Evaluation of toxicity and degradation of a chlorophenol mixture by the laccase produced by Trametes pubescens. Bioresour. Technol. 2011, 102, 3632–3635. [Google Scholar] [CrossRef] [PubMed]
- Eichlerová, I.; Šnajdr, J.; Baldrian, P. Laccase activity in soils: Considerations for the measurement of enzyme activity. Chemosphere 2012, 88, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- De Nobili, M.; Bravo, C.; Chen, Y. The spontaneous secondary synthesis of soil organic matter components: A critical examination of the soil continuum model theory. Appl. Soil Ecol. 2020, 154, 103655. [Google Scholar] [CrossRef]
- Trubitsina, L.I.; Lisov, A.V.; Belova, O.V.; Trubitsin, I.V.; Demin, V.V.; Konstantinov, A.I.; Zavarzina, A.G.; Leontievsky, A.A. Transformation of low molecular compounds and soil humic acid by two domain laccase of Streptomyces puniceus in the presence of ferulic and caffeic acids. PLoS ONE. 2020, 15, e0239005. [Google Scholar] [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Thorn, R.G.; Reddy, C.A.; Harris, D.; Paul, E.A. Isolation of saprophytic basidiomycetes from soil. Appl. Environ. Microbiol. 1996, 62, 4288–4292. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Choi, J.; Asiegbu, F.O.; Lee, Y.H. Comparative genomics platform and phylogenetic analysis of fungal laccases and multi-copper oxidases. Mycobiology 2020, 48, 373–382. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [Green Version]
- Bankolea, P.O.; Semple, K.T.; Jeon, B.H.; Govindwar, S.P. Biodegradation of fluorene by the newly isolated marine-derived fungus, Mucor irregularis strain bpo1 using response surface methodology. Ecotoxicol. Environ. Saf. 2020, 208, 111619. [Google Scholar] [CrossRef]
- Kanmani, P.; Karuppasamy, P. Studies on lignocellulose biodegradation of coir waste in solid state fermentation using Phanerocheate chrysosporium and Rhizopus stolonifer. Afr. J. Biotechnol. 2009, 8, 6880–6887. [Google Scholar] [CrossRef]
- Geethanjali, P.A.; Gowtham, H.G.; Jayashankar, M. Optimization of culture conditions for hyper-production of laccase from an indigenous litter dwelling fungus Mucor circinelloides GL1. Environ. Sustain. 2020, 3, 141–146. [Google Scholar] [CrossRef]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 2020, 11, 7. [Google Scholar] [CrossRef]
- Sideris, C.P.; Paxton, G.E. A new species of Mortierella. Mycologia 1929, 21, 175–177. [Google Scholar] [CrossRef]
- Heinfling, A.; Martinez, A.T.; Martinez, M.J.; Bergbauer, M.; Szewzyk, U. Purification and characterization of peroxidases from the dye-decolorizing fungus Bjerkandera adusta. FEMS Microbiol. Lett. 1998, 428, 43–50. [Google Scholar] [CrossRef]
- Wariishi, H.; Valli, K.; Gold, M.H. Manganese(II) oxidation by manganese peroxidase from the basidiomycete Phanerochaete chrysosporium. Kinetic mechanism and role of helators. J. Biol. Chem. 1992, 267, 23688–23695. [Google Scholar] [CrossRef]
- Timofeevski, S.L.; Reading, N.S.; Aust, S.D. Mechanisms for protection against inactivation of manganese peroxidase by hydrogen peroxide. Arch. Biochem. Biophys. 1998, 356, 287–295. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. In Methods of Biochemical Analysis; Glick, D., Ed.; Interscience: New York, NY, USA, 1954; Volume 1, pp. 357–408. [Google Scholar]
- Saito, T.; Hong, P.; Kato, K.; Okazaki, M.; Inagaki, H.; Maeda, S.; Yokogawa, Y. Purification and characterization of an extracellular laccase of a fungus (family Chaetomiaceae) isolated from soil. Enzym. Microb. Technol. 2003, 33, 520–526. [Google Scholar] [CrossRef]
- Zavarzina, A.G.; Kravchenko, E.G.; Konstantinov, A.I.; Perminova, I.V.; Chukov, S.N.; Demin, V.V. Comparison of the properties of humic acids extracted from soils by alkali in the presence and absence of oxygen. Eurasian Soil Sci. 2019, 52, 880–891. [Google Scholar] [CrossRef]
- Zavarzina, A.G.; Demin, V.V. Acid–base properties of humic acids as seen from potentiometric titration data. Eurasian Soil Sci. 1999, 32, 1246–1256. [Google Scholar]
- Zavarzina, A.G.; Demin, V.V.; Nifantieva, T.I.; Shkinev, V.M.; Danilova, T.V.; Spivakov, B.Y. Extraction of humic acids and their fractions in polyethylene glycol-based aqueous biphasic systems. Anal. Chim. Acta 2002, 452, 95–103. [Google Scholar] [CrossRef]
- Hertkorn, N.; Permin, A.; Perminova, I.; Kovalevskii, D.; Yudov, M.; Petrosyan, V.; Kettrup, A. Comparative analysis of partial structures of a peat humic and fulvic acid using one- and two-dimensional nuclear magnetic resonance spectroscopy. J. Environ. Qual. 2002, 31, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Galhaup, C.; Haltrich, D. Enhanced formation of laccase activity by the white-rot fungus Trametes pubescens in the presence of copper. Appl. Microbiol. Biotechnol. 2001, 56, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Giardina, P.; Palmieri, G.; Scaloni, A.; Fontanella, B.; Faraco, V.; Cennamo, G.; Sannia, G. Protein and gene structure of a blue laccase from Pleurotus ostreatus. Biochem. J. 1999, 341, 655–663. [Google Scholar] [CrossRef]
- D’Souza, T.M.; Merritt, C.S.; Reddy, C.A. Lignin-modifying enzymes of the white rot basidiomycete Ganoderma lucidum. Appl. Environ. Microbiol. 1999, 65, 5307–5313. [Google Scholar] [CrossRef] [Green Version]
- Eggert, C.; Temp, U.; Eriksson, K.E. The ligninolytic system of the white rot fungus Pycnoporus cinnabarinus: Purification and characterization of the laccase. Appl. Environ. Microbiol. 1996, 62, 1151–1158. [Google Scholar] [CrossRef] [Green Version]
- Elisashvili, V.; Kachlishvili, E.; Khardziani, T.; Agathos, S.N. Effect of aromatic compounds on the production of laccase and manganese peroxidase by white-rot basidiomycetes. J. Ind. Microbiol. Biotechnol. 2010, 37, 1091–1096. [Google Scholar] [CrossRef]
- Arora, D.S.; Rampal, P. Laccase production by some Phlebia species. J. Basic Microbiol. 2002, 42, 295–301. [Google Scholar] [CrossRef]
- Schneider, W.; Fontana, R.; Mendonça, S.; Siqueira, F.G.; Dillon, A.J.; Camassola, M. High level production of laccases and peroxidases from the newly isolated white-rot basidiomycete Marasmiellus palmivorus VE111 in a stirred-tank bioreactor in response to different carbon and nitrogen sources. Process Biochem. 2018, 69, 1–11. [Google Scholar] [CrossRef]
- Pointing, S.; Jones, E.; Vrijmoed, L. Optimization of laccase production by Pycnoporus sanguineus in submerged liquid culture. Mycologia 2000, 92, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Buddhika, U.V.A.; Savocchia, S.; Steel, C.C. Copper induces transcription of BcLCC2 laccase gene in phytopathogenic fungus Botrytis cinerea. Mycology 2020, 11, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, A.; Falcón, M.A.; Carnicero, A.; Perestelo, F.; De la Fuente, G.; Trojanowski, J. Laccase activities of Penicillium chrysogenum in relation to lignin degradation. Appl. Microbiol. Biotechnol. 1996, 45, 399–403. [Google Scholar] [CrossRef]
- Assavanig, A.; Amornikitticharoen, B.; Ekpaisal, N.; Meevootisom, V.; Flegel, T.W. Isolation, characterization and function of laccase from Trichoderma. Appl. Microbiol. Biotechnol. 1992, 38, 198–202. [Google Scholar] [CrossRef]
- Góralczyk-Bińkowska, A.; Jasińska, A.; Długoński, A.; Płociński, P.; Długoński, J. Laccase activity of the ascomycete fungus Nectriella pironii and innovative strategies for its production on leaf litter of an urban park. PLoS ONE 2020, 15, e0231453. [Google Scholar] [CrossRef] [Green Version]
- Xu, F. Effects of redox potential and hydroxide inhibition on the pH activity profile of fungal laccases. J. Biol. Chem. 1997, 272, 924–928. [Google Scholar] [CrossRef] [Green Version]
- Heinzkill, M.; Bech, L.; Halkier, T.; Schneider, P.; Anke, T. Characterization of laccases and peroxidases from woodrotting fungi (family Coprinaceae). Appl. Environ. Microbiol. 1998, 64, 1601–1606. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.R.; Luo, Z.H.; Kwok-Kei Chow, R.; Vrijmoed, L.L. Purification and characterization of an extracellular laccase from the anthracene-degrading fungus Fusarium solani MAS2. Bioresour. Technol. 2010, 65, 4943–4948. [Google Scholar] [CrossRef]
- Nagai, M.; Kawata, M.; Watanabe, H.; Ogawa, M.; Saito, K.; Takesawa, T.; Kanda, K.; Sato, T. Important role of fungal intracellular laccase for melanin synthesis: Purification and characterization of an intracellular laccase from Lentinula edodes fruit bodies. Microbiology 2003, 149, 2455–2462. [Google Scholar] [CrossRef]
- Garcia, T.A.; Santiago, M.F.; Ulhoa, C.J. Studies on the Pycnoporus sanguineus CCT-4518 laccase purified by hydrophobic interaction chromatography. Appl. Microbiol. Biotechnol. 2007, 75, 311–318. [Google Scholar] [CrossRef]
- Fakoussa, R.; Frost, P. In vivo-decolorization of coal-derived humic acids by laccase-excreting fungus Trametes versicolor. Appl. Microbiol. Biotechnol. 1999, 52, 60–65. [Google Scholar] [CrossRef]
- Zahmatkesh, M.; Spanjers, H.; Toran, M.J.; Blánquez, P.; van Lier, J.B. Bioremoval of humic acid from water by white rot fungi: Exploring the removal mechanisms. AMB Express 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chefetz, B.; Chen, Y.; Hadar, Y. Purification and characterization of laccase from Chaetomium thermophilium and its role in humification. Appl. Environ. Microbiol. 1998, 64, 3175–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olbrich, A.C.; Schild, J.N.; Urlacher, V.B. Correlation between the T1 copper reduction potential and catalytic activity of a small laccase. J. Inorg. Biochem. 2019, 201, 110843. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Km, μM | kkat, s−1 | kkat/Km, μM−1 × s−1 |
|---|---|---|---|
| ABTS | 32.7 ± 1.7 | 41.6 ± 1.5 | 1.27 |
| K4[Fe(CN)6] | 107 ± 4.3 | 40.5 ± 1.6 | 0.38 |
| 2,6-dimethoxyphenol | 35.9 ± 2.1 | 1.65 ± 0.007 | 4.6 × 10−2 |
| Guaiacol | 49.3 ± 6.5 | 0.93 ± 0.01 | 1.8 × 10−2 |
| Hydroquinone | 413 ± 30 | 9.43 ± 0.3 | 2.2 × 10−2 |
| Sample | Content, Mass%/Atomic% | Atomic Ratios | ||||||
|---|---|---|---|---|---|---|---|---|
| Ash, % | C | H | N | O | H:C | O:C | C:N | |
| 1 Retisol HA | 2.6 | 52.9/35.3 | 5.2/42.1 | 4.5/2.6 | 40.0/20.0 | 1.2 | 0.6 | 13.7 |
| Compost HA | 2.0 | 51.8/38.4 | 4.0/35.5 | 5.2/3.3 | 41.0/22.8 | 0.9 | 0.6 | 11.7 |
| 2 Peat HA | 1.9 | 51.2/35.9 | 4.8/40.7 | 2.8/1.7 | 41.2/21.7 | 1.1 | 0.6 | 21.1 |
| Spectral Region, ppm/Structural Group | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Aliphatic Fragments | Aromatic Fragments | Carboxyls, Ketones and Quinones | ||||||||
| Sample | 0–48 | 48–64 | 64–90 | 90–109 | 109–145 | 145–164 | 164–187 | 187–220 | 2 Car/Cal | |
| CHn | CHnO | CHO | OCO | Car | CarO | COO | C=O | |||
| 1 Retisol HA | 13.0 | 10.0 | 13.0 | 3.0 | 21.0 | 13.0 | 20.0 | 8.0 | 0.9 | |
| Compost HA | 18.0 | 8.6 | 9.1 | 4.3 | 26.1 | 9.5 | 18.9 | 5.5 | 0.9 | |
| Peat HA | 17.8 | 9.6 | 9.7 | 8.6 | 26.8 | 8.3 | 15.2 | 3.9 | 0.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisov, A.; Belova, O.; Zavarzina, A.; Konstantinov, A.; Leontievsky, A. The Role of Laccase from Zygomycetous Fungus Mortierella elasson in Humic Acids Degradation. Agronomy 2021, 11, 2169. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112169
Lisov A, Belova O, Zavarzina A, Konstantinov A, Leontievsky A. The Role of Laccase from Zygomycetous Fungus Mortierella elasson in Humic Acids Degradation. Agronomy. 2021; 11(11):2169. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112169
Chicago/Turabian StyleLisov, Alexander, Oksana Belova, Anna Zavarzina, Andrey Konstantinov, and Alexey Leontievsky. 2021. "The Role of Laccase from Zygomycetous Fungus Mortierella elasson in Humic Acids Degradation" Agronomy 11, no. 11: 2169. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112169