
Carbohydrate Assimilation and Translocation Regulate Grain Yield Formation in Wheat Crops (Triticum aestivum L.) under Post-Flowering Waterlogging

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Site
2.2. Experiment Design
2.3. Measurements and Methods
2.3.1. SPAD Value and Leaf Gas Exchange of Flag Leaf
2.3.2. Parameters of Wheat Grain Filling
- (1)
- Start date of the peak grain filling period: t1 = [A − ln(2 + 1.732)]/(−B).
- (2)
- End date of the peak grain filling period: t2 = [A + ln(2 + 1.732)]/(−B).
- (3)
- End of grain filling (Y up to 99% K) date: t3 = (4.595 − 12 + A)/(−B).
- (4)
- Date when the maximum grain filling rate appears: Tm = −A/B, and the maximum grain filling rate Vm = −BK/4.
- (5)
- T1, T2, and T3 represent the grain filling rates of the gradual, rapid, and slow increase stages, respectively. T1 = t1, T2 = t2 − t1, and T3 = t3 − t2.
- (6)
- Grain filling duration: T = t3, and mean grain filling rate: Va = K/t.
2.3.3. Dry Matter Accumulation and Distribution
2.3.4. Grain Yield and Components
2.4. Data Analysis
3. Results
3.1. SPAD of Flag Leaves
3.2. Net Photosynthetic Rate of Flag Leaves
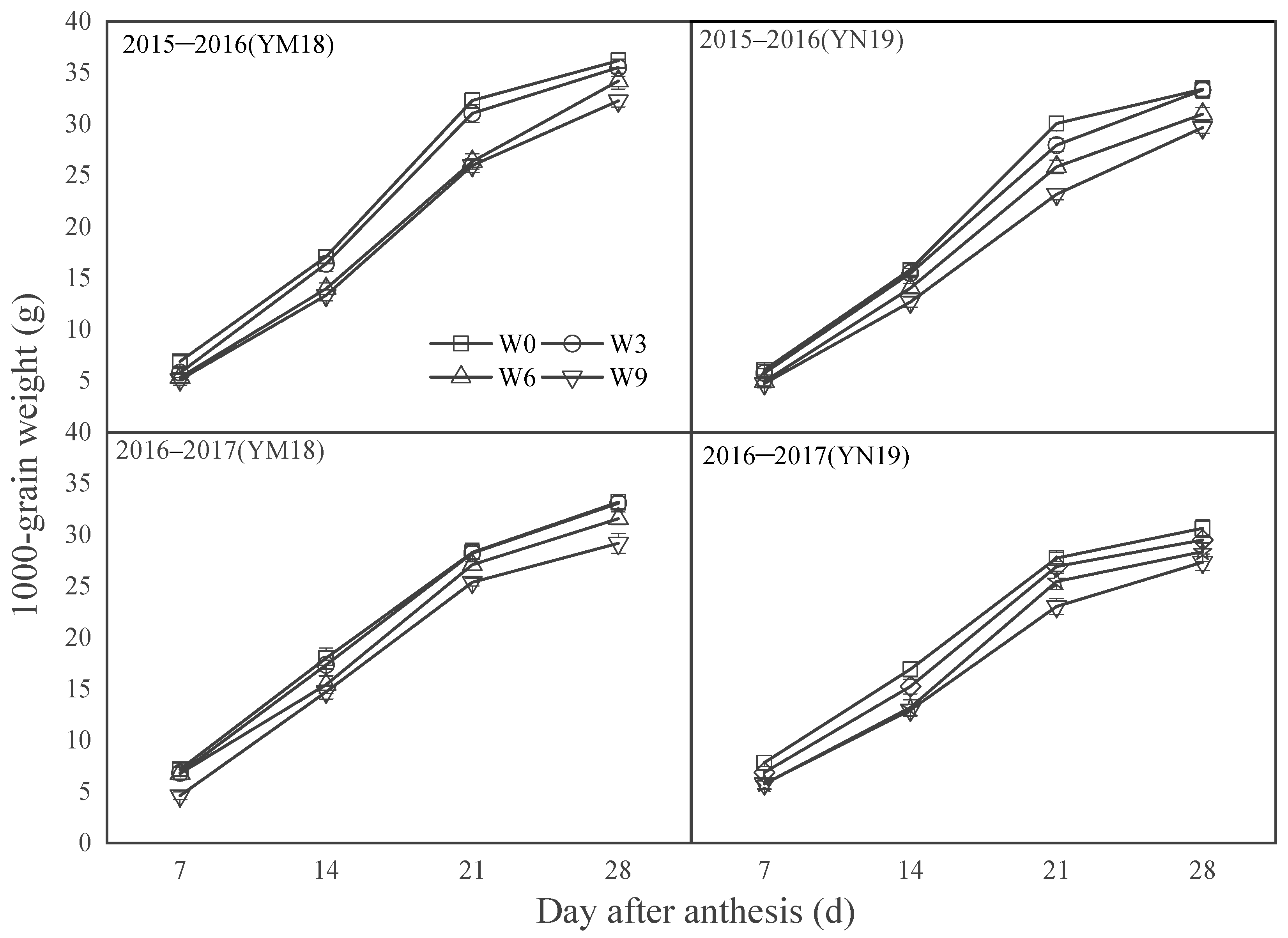
3.3. Grain Weight
3.4. Grain Filling Characteristic Parameters
3.5. Dry Matter Accumulation and Distribution
3.6. Grain Yield and Its Components
4. Discussion
4.1. Effect of Waterlogging on Photosynthetic Characteristics
4.2. Effect of Waterlogging on Grain Weight and Grain Filling Rate
4.3. Effect of Waterlogging on Dry Matter Distribution
4.4. Effect of Waterlogging on Yield and Components
5. Conclusions
6. Key Findings
Author Contributions
Funding
Conflicts of Interest
References
- Wang, X.; Liu, F.; Jiang, D. Priming: A promising strategy for crop production in response to future climate. J. Integr. Agric. 2017, 16, 2709–2716. [Google Scholar] [CrossRef]
- Arguello, M.N.; Mason, R.E.; Roberts, T.L.; Subramanian, N.; Acuna, A.; Addison, C.K.; Lozada, D.N.; Miller, R.G.; Gbur, E. Performance of soft red winter wheat subjected to field soil waterlogging: Grain yield and yield components. Field Crops Res. 2016, 194, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Fan, X.; Dai, T.; Cao, W. Nitrogen fertiliser rate and post-anthesis waterlogging effects on carbohydrate and nitrogen dynamics in wheat. Plant Soil 2008, 304, 301–314. [Google Scholar] [CrossRef]
- Sundgren, T.K.; Uhlen, A.K.; Lillemo, M.; Briese, C.; Wojciechowski, T. Rapid seedling establishment and a narrow root stele promotes waterlogging tolerance in spring wheat. J. Plant Physiol. 2018, 227, 45–55. [Google Scholar] [CrossRef]
- Pedersen, O.; Perata, P.; Voesenek, L. Flooding and low oxygen responses in plants. Funct. Plant Biol. 2017, 44, iii–vi. [Google Scholar] [CrossRef] [PubMed]
- Amzallag, G.N. Maturation of integrated functions during development. I. Modifications of the regulatory network during transition periods in Sorghum bicolor. Plant. Cell Environ. 2001, 24, 337–345. [Google Scholar] [CrossRef]
- Najeeb, U.; Bange, M.P.; Tan, D.K.Y.; Atwell, B.J. Consequences of waterlogging in cotton and opportunities for mitigation of yield losses. AoB Plants 2015, 7, plv080. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wei, F.; Yu, S.; Yu, Z. Effect of waterlogging on senescence of winter wheat root system at booting stage. Chin. J. Appl. Ecol. 2000, 11, 723–726. [Google Scholar]
- Wany, A.; Kumari, A.; Gupta, K.J. Nitric oxide is essential for the development of aerenchyma in wheat roots under hypoxic stress. Plant Cell Environ. 2017, 40, 3003–3017. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Tao, Q.; Zhang, G. Effect of waterlogging on senescence of flag leaf and root of wheat Yangmai 5. Chin. J. Appl. Ecol. 2002, 13, 1519–1521. [Google Scholar]
- Qi, Y.H.; Mao, F.F.; Zhou, Z.Q.; Liu, D.C.; Yu, M.; Deng, X.Y.; Li, J.W.; Mei, F.Z. The release of cytochrome c and the regulation of the programmed cell death progress in the endosperm of winter wheat (Triticum aestivum L.) under waterlogging. Protoplasma 2018, 255, 1651–1655. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Y.; Zhou, Z.Q.; Yang, C.N.; Jiang, Z.; Li, J.; Cheng, X.; Guo, Y. Effects of waterlogging on amyloplasts and programmed cell death in endosperm cells of Triticum aestivum L. Protoplasma 2013, 250, 1091–1103. [Google Scholar] [CrossRef]
- Marti, J.; Savin, R.; Slafer, G.A. Wheat Yield as Affected by Length of Exposure to Waterlogging During Stem Elongation. J. Agron. Crop Sci. 2015, 201, 473–486. [Google Scholar] [CrossRef]
- Ghobadi, M.E.; Ghobadi, M.; Zebarjadi, A. Effect of waterlogging at different growth stages on some morphological traits of wheat varieties. Int. J. Biometeorol. 2017, 61, 635–645. [Google Scholar] [CrossRef]
- Najeeb, U.; Atwell, B.J.; Bange, M.P.; Tan, D. Aminoethoxyvinylglycine (AVG) ameliorates waterlogging-induced damage in cotton by inhibiting ethylene synthesis and sustaining photosynthetic capacity. Plant Growth Regul. 2015, 76, 83–98. [Google Scholar] [CrossRef]
- Zheng, C.; Dong, J.; Liu, F.; Dai, T.; Jing, Q.; Cao, W. Effects of salt and waterlogging stresses and their combination on leaf photosynthesis, chloroplast ATP synthesis, and antioxidant capacity in wheat. Plant Sci. Int. J. Exp. Plant Biol. 2009, 176, 575–582. [Google Scholar] [CrossRef]
- Ding, J.; Huang, Z.; Zhu, M.; Li, C.; Zhu, X.; Guo, W.; Zhou, M. Does cyclic water stress damage wheat yield more than a single stress? PLoS ONE 2018, 13, e0195535. [Google Scholar] [CrossRef] [Green Version]
- Arduini, I.; Baldanzi, M.; Pampana, S. Reduced Growth and Nitrogen Uptake During Waterlogging at Tillering Permanently Affect Yield Components in Late Sown Oats. Front. Plant Sci. 2019, 10, 1087–1105. [Google Scholar] [CrossRef] [Green Version]
- Dickin, E.; Wright, D. The effects of winter waterlogging and summer drought on the growth and yield of winter wheat (Triticum aestivum L.). Eur. J. Agron. 2008, 28, 234–244. [Google Scholar] [CrossRef]
- Arduini, I.; Orlandi, C.; Pampana, S.; Masoni, A. Waterlogging at tillering affects spike and spikelet formation in wheat. Crop Pasture Sci. 2016, 67, 703–711. [Google Scholar] [CrossRef]
- Xiaoming, B. Study on identification stage and index of waterlogging tolerance in various wheat genotypes (Triticum aestivum L.). Acta Agricul Turae Shanghai 1997, 2, 7. [Google Scholar]
- Ding, J.; Liang, P.; Wu, P.; Zhu, M.; Li, C.; Zhu, X.; Guo, W. Identifying the critical stage near anthesis for waterlogging on wheat yield and its components in the Yangtze River Basin, China. Agronomy 2020, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- de San Celedonio, R.P.; Abeledo, L.G.; Miralles, D.J. Identifying the critical period for waterlogging on yield and its components in wheat and barley. Plant Soil 2014, 378, 265–277. [Google Scholar] [CrossRef]
- Araki, H.; Hamada, A.; Hossain, M.A.; Takahashi, T. Waterlogging at jointing and/or after anthesis in wheat induces early leaf senescence and impairs grain filling. Field Crops Res. 2012, 137, 27–36. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Hamada, A.; Hossain, A.; Takahashi, T.; Araki, H. Grain growth and leaf senescence in wheat exposed to waterlogging during jointing stage and post-anthesis. Jpn. J. Crop Sci. Extra Issue 2011, 80, 158. [Google Scholar]
- Monostori, I.; Árendás, T.; Hoffman, B.; Galiba, G.; Gierczik, K.; Szira, F.; Vágújfalvi, A. Relationship between SPAD value and grain yield can be affected by cultivar, environment and soil nitrogen content in wheat. Euphytica 2016, 211, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Mu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Jing, Q.; Cao, W. Long-term low radiation decreases leaf photosynthesis, photochemical efficiency and grain yield in winter wheat. J. Agron. Crop Sci. 2010, 196, 38–47. [Google Scholar] [CrossRef]
- Wu, X.; Tang, Y.; Li, C.; Wu, C. Characterization of the rate and duration of grain filling in wheat in southwestern China. Plant Prod. Sci. 2018, 21, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Li, J.F.; Fan, Z.R.; Zhang, Y.Q.; Wang, Z.; Gao, X.; Shi, J. Logistic equation fitting Xinjiang spring wheat filling dynamics. Xinjiang Agric. Sci. 2019, 56, 2006–2014. [Google Scholar]
- Cheng, X.X.; Yu, M.; Zhang, N.; Zhou, Z.Q.; Xu, Q.T.; Mei, F.Z.; Qu, L.H. Reactive oxygen species regulate programmed cell death progress of endosperm in winter wheat (Triticum aestivum L.) under waterlogging. Protoplasma 2016, 253, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Kumutha, D.; Ezhilmathi, K.; Sairam, R.K.; Srivastava, G.C.; Deshmukh, P.S.; Meena, R.C. Waterlogging induced oxidative stress and antioxidant activity in pigeonpea genotypes. Biol. Plant. 2009, 53, 75–84. [Google Scholar] [CrossRef]
- Hui, Z.; Dai, T.B.; Dong, J.; Jing, Q.; Cao, W.X. Effects of drought and waterlogging on flag leaf post-anthesis photosynthetic characteristics and assimilates translocation in winter wheat under high temperature. Chin. J. Appl. Ecol. 2007, 18, 333–338. [Google Scholar]
- Pampana, S.; Masoni, A.; Arduini, I. Grain yield of durum wheat as affected by waterlogging at tillering. Cereal Res. Commun. 2016, 44, 706–716. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Araki, H.; Takahashi, T. Poor grain filling induced by waterlogging is similar to that in abnormal early ripening in wheat in Western Japan. Field Crops Res. 2011, 123, 100–108. [Google Scholar] [CrossRef]
- Setter, T.L.; Waters, I. Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil 2003, 253, 1–34. [Google Scholar] [CrossRef]
- Hayashi, T.; Yoshida, T.; Fujii, K.; Mitsuya, S.; Tsuji, T.; Okada, Y.; Hayashi, E.; Yamauchi, A. Maintained root length density contributes to the waterlogging tolerance in common wheat (Triticum aestivum L.). Field Crops Res. 2013, 152, 27–35. [Google Scholar] [CrossRef]
- Egli, D.B. Seed Biology and Yield of Grain Crops; Centre for Agriculture and Biosciences International: Oxfordshire, UK, 2017. [Google Scholar]
- Abid, M.; Shao, Y.; Liu, S.; Wang, F.; Gao, J.; Jiang, D.; Tian, Z.; Dai, T. Pre-drought priming sustains grain development under post-anthesis drought stress by regulating the growth hormones in winter wheat (Triticum aestivum L.). Planta 2017, 246, 509–524. [Google Scholar] [CrossRef]
- Zhao, H.; Dai, T.; Jing, Q.; Jiang, D.; Cao, W. Leaf senescence and grain filling affected by post-anthesis high temperatures in two different wheat cultivars. Plant Growth Regul. 2007, 51, 149–158. [Google Scholar] [CrossRef]
- Zhang, J.; Xue, X.; Li, N.; Li, H.; Zhang, L.; Song, J. Effects of Water Stress on Photosynthetic Characteristics, Dry Matter Production and Yield of Winter Wheat at Flowering Stage. J. Arid Meteorol. 2019, 37, 447–453. [Google Scholar]
- Ma, S.; Han, X.; Wang, Y.; Huang, Z.; Zhang, W.; Fan, Y.; Ma, Y. Effect of waterlogging stress after anthesis on the root morphology, physiology, dry matter distribution and yield in wheat. J. Nanjing Agric. Univ. 2021, 44, 614–621. [Google Scholar]
- Hays, D.B.; Do, J.H.; Mason, R.E.; Morgan, G.; Finlayson, S.A. Heat stress induced ethylene production in developing wheat grains induces kernel abortion and increased maturation in a susceptible cultivar. Plant Sci. 2007, 172, 1113–1123. [Google Scholar] [CrossRef]
- Wang, X.; Liu, D.; Wei, M.; Man, J. Spraying 6-BA could alleviate the harmful impacts of waterlogging on dry matter accumulation and grain yield of wheat. PeerJ 2020, 8, e8193. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Liu, J.; Dai, T.; Jing, Q.; Cao, W.; Jiang, D. Alterations in photosynthesis and antioxidant enzyme activity in winter wheat subjected to post-anthesis water-logging. Photosynthetica 2008, 46, 21–27. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Season | Organic Matter (g·kg−1) | Total N (g·kg−1) | Available P (mg·kg−1) | Available K (mg·kg−1) | Available N (mg·kg−1) |
|---|---|---|---|---|---|
| 2015–2016 | 18.5 | 1.2 | 12.1 | 127.4 | 133.5 |
| 2016–2017 | 17.3 | 1.2 | 11.7 | 142.9 | 118.5 |
| Treatments | 7 Days after Flowering | 14 Days after Flowering | 21 Days after Flowering | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SPAD | Pn | GW | SPAD | Pn | GW | SPAD | Pn | GW | |
| Years | <0.001 | 0.240 | 0.002 | <0.001 | <0.001 | 0.068 | <0.001 | 0.023 | 0.001 |
| Cultivar | 0.001 | 0.043 | 0.708 | <0.001 | 0.008 | 0.001 | <0.001 | <0.001 | <0.001 |
| Waterlogging | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Years × Cultivar | 0.691 | 0.095 | 0.248 | 0.001 | 0.001 | 0.099 | 0.624 | <0.001 | 0.307 |
| Years × Waterlogging | 0.434 | 0.938 | 0.489 | 0.092 | 0.540 | 0.839 | <0.001 | 0.062 | 0.005 |
| Cultivar × Waterlogging | 0.412 | 0.342 | 0.411 | <0.001 | 0.974 | 0.967 | 0.002 | 0.939 | 0.235 |
| Years × Cultivar × Waterlogging | 0.777 | 0.295 | 0.501 | 0.267 | 0.831 | 0.613 | 0.081 | 0.568 | 0.302 |
| Growing Season | Cultivar | Treatment | Logistic Equation | R2 | Tm (d) | Vm (mg·grain−1·d−1) | T (d) | Va (mg·grain−1·d−1) | T1 (d) | T2 (d) | T3 (d) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2015–2016 | YM18 | W0 | Y = 38.6399/(1 + e2.7097−0.172703t) | 0.9916 | 15.7 | 1.7 | 42.3 | 0.9 | 8.1 | 14.7 | 19.0 |
| W3 | Y = 37.9719/(1 + e2.9108−0.182388t) | 0.9936 | 16.0 | 1.7 | 41.2 | 0.9 | 8.7 | 15.0 | 18.0 | ||
| W6 | Y = 35.2239/(1 + e4.1522−0.278026t) | 0.9897 | 14.9 | 2.5 | 31.5 | 1.1 | 10.2 | 13.9 | 11.8 | ||
| W9 | Y = 33.1104/(1 + e5.1911−0.355704t) | 0.9728 | 14.6 | 2.9 | 27.5 | 1.2 | 10.9 | 13.6 | 9.2 | ||
| YN19 | W0 | Y = 34.7171/(1 + e3.4524−0.242375t) | 0.9950 | 14.2 | 2.1 | 33.2 | 1.1 | 8.8 | 13.3 | 13.5 | |
| W3 | Y = 34.6754/(1 + e3.2611−0.221364t) | 0.9986 | 14.7 | 1.9 | 35.5 | 1.0 | 8.8 | 13.7 | 14.8 | ||
| W6 | Y = 32.1739/(1 + e3.4408−0.229923t) | 0.9993 | 15.0 | 1.9 | 35.0 | 0.9 | 9.2 | 14.0 | 14.3 | ||
| W9 | Y = 31.0240/(1 + e3.2146−0.206115t) | 0.9969 | 15.6 | 1.6 | 37.9 | 0.8 | 9.2 | 14.6 | 15.9 | ||
| 2016–2017 | YM18 | W0 | Y = 36.2162/(1 + e2.6678−0.187727t) | 0.9991 | 14.2 | 1.7 | 38.7 | 0.9 | 7.2 | 13.2 | 17.5 |
| W3 | Y = 35.8357/(1 + e2.7933−0.194550t) | 0.9996 | 14.4 | 1.7 | 38.0 | 0.9 | 7.6 | 13.4 | 16.9 | ||
| W6 | Y = 34.1698/(1 + e2.8480−0.194831t) | 0.9980 | 14.6 | 1.7 | 38.2 | 0.9 | 7.9 | 13.6 | 16.8 | ||
| W9 | Y = 30.6619/(1 + e3.3618−0.234306t) | 0.9998 | 14.4 | 1.8 | 34.0 | 0.9 | 8.7 | 13.4 | 14.0 | ||
| YN19 | W0 | Y = 33.3013/(1 + e2.5785−0.191689t) | 0.9962 | 13.5 | 1.6 | 37.4 | 0.9 | 6.6 | 12.5 | 17.1 | |
| W3 | Y = 32.7110/(1 + e2.7763−0.195348t) | 0.9925 | 14.2 | 1.6 | 37.7 | 0.9 | 7.5 | 13.2 | 16.8 | ||
| W6 | Y = 30.6996/(1 + e3.1501−0.214451t) | 0.9927 | 14.7 | 1.7 | 36.1 | 0.9 | 8.6 | 13.7 | 15.3 | ||
| W9 | Y = 29.5724/(1 + e2.8629−0.192369t) | 0.9981 | 14.9 | 1.4 | 38.8 | 0.8 | 8.0 | 13.9 | 17.0 |
| Growing Season | Cultivar | Treatment | Grain | Stem + Sheath | Spike Axis + Glume | |||
|---|---|---|---|---|---|---|---|---|
| Amount (kg·hm−2) | Proportion of the Total (%) | Amount (kg·hm−2) | Proportion of the Total (%) | Amount (kg·hm−2) | Proportion of the Total (%) | |||
| 2015–2016 | YM18 | W0 | 7380.8 a | 47.1 a | 6929.7 a | 44.2 a | 1370.7 d | 8.7 d |
| W3 | 7064.4 a | 45.9 a | 6796.6 a | 44.2 a | 1530.2 c | 9.9 c | ||
| W6 | 6391.8 b | 42.3 b | 6377.8 b | 42.2 b | 2329.4 b | 15.4 b | ||
| W9 | 6073.4 c | 41.5 b | 5875.2 c | 40.1 c | 2697.8 a | 18.4 a | ||
| YN19 | W0 | 5603.7 a | 39.8 a | 7431.0 a | 52.8 b | 1032.9 d | 7.3 d | |
| W3 | 5382.5 a | 38.6 a | 7185.8 a | 51.5 b | 1390.3 c | 10.0 c | ||
| W6 | 4508.1 b | 34.4 b | 6920.9 b | 52.8 b | 1686.3 b | 12.9 b | ||
| W9 | 4039.5 c | 31.4 c | 7001.5 a | 54.5 a | 1804.5 a | 14.0 a | ||
| 2016–2017 | YM18 | W0 | 8011.2 a | 47.5 a | 7042.7 a | 41.8 a | 1783.4 c | 10.6 c |
| W3 | 7807.3 a | 47.0 a | 6764.4 a | 40.7 a | 2016.5 b | 12.3 b | ||
| W6 | 7231.1 b | 46.6 a | 6451.9 b | 41.1 a | 2035.4 b | 12.8 b | ||
| W9 | 6887.9 c | 44.8 b | 6138.0 c | 39.9 a | 2326.7 a | 15.2 a | ||
| YN19 | W0 | 7548.6 a | 46.7 a | 7045.5 a | 43.5 a | 1566.1 b | 9.7 d | |
| W3 | 7440.7 a | 45.4 a | 7040.7 a | 43.0 a | 1888.1 a | 11.5 c | ||
| W6 | 6745.5 b | 43.7 b | 6778.8 b | 43.9 a | 1915.9 a | 12.4 b | ||
| W9 | 6038.8 c | 42.3 b | 6271.6 c | 43.9 a | 1952.6 a | 13.7 a | ||
| Cultivar | Growing Season | Treatment | Number of Spikes (×104·hm−2) | Kernels Per Spike | 1000-Grain Weight (g) | Yield (kg·hm−2) |
|---|---|---|---|---|---|---|
| YM18 | 2015–2016 | W0 | 561.7 a | 38.0 a | 36.4 a | 7380.8 a |
| W3 | 553.6 a | 37.6 a | 36.2 a | 7064.4 a | ||
| W6 | 550.9 a | 36.4 a | 34.5 b | 6391.8 b | ||
| W9 | 540.0 a | 34.9 b | 32.0 c | 6073.4 c | ||
| 2016–2017 | W0 | 587.4 a | 39.2 a | 35.9 a | 8011.3 a | |
| W3 | 576.9 a | 38.3 a | 35.5 a | 7807.3 a | ||
| W6 | 575.7 a | 37.0 b | 33.5 b | 7231.1 b | ||
| W9 | 578.3 a | 35.3 b | 30.6 c | 6857.9 c | ||
| YN19 | 2015–2016 | W0 | 520.9 a | 31.6 a | 34.1 a | 5603.7 a |
| W3 | 505.4 a | 30.7 a | 33.8 a | 5382.5 a | ||
| W6 | 497.6 a | 30.5 a | 31.5 b | 4508.1 b | ||
| W9 | 495.2 a | 28.8 b | 29.8 c | 4039.5 c | ||
| 2016–2017 | W0 | 594.0 a | 38.3 a | 33.0 a | 7548.6 a | |
| W3 | 591.8 a | 37.8 a | 32.6 a | 7440.7 a | ||
| W6 | 580.5 a | 35.7 b | 30.4 b | 6745.6 b | ||
| W9 | 580.2 a | 33.5 c | 28.8 c | 6038.8 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, S.; Gai, P.; Wang, Y.; Ullah, N.; Zhang, W.; Fan, Y.; Shan, Y.; Huang, Z.; Hu, X. Carbohydrate Assimilation and Translocation Regulate Grain Yield Formation in Wheat Crops (Triticum aestivum L.) under Post-Flowering Waterlogging. Agronomy 2021, 11, 2209. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112209
Ma S, Gai P, Wang Y, Ullah N, Zhang W, Fan Y, Shan Y, Huang Z, Hu X. Carbohydrate Assimilation and Translocation Regulate Grain Yield Formation in Wheat Crops (Triticum aestivum L.) under Post-Flowering Waterlogging. Agronomy. 2021; 11(11):2209. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112209
Chicago/Turabian StyleMa, Shangyu, Panpan Gai, Yanyan Wang, Najeeb Ullah, Wenjing Zhang, Yonghui Fan, Yajing Shan, Zhenglai Huang, and Xia Hu. 2021. "Carbohydrate Assimilation and Translocation Regulate Grain Yield Formation in Wheat Crops (Triticum aestivum L.) under Post-Flowering Waterlogging" Agronomy 11, no. 11: 2209. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112209